
犬体内两种人兽共患棘口吸虫的分子鉴定
2021-01-04 07:37:10邱鸿宇兰卓果馨儒吴婷婷姜岩金程佳张晓轩王春仁
黑龙江八一农垦大学学报 2020年6期
邱鸿宇,兰卓,果馨儒,吴婷婷,姜岩,金程佳,张晓轩,王春仁
(黑龙江八一农垦大学动物科技学院,大庆 163319)
棘口吸虫为棘口科(Echinostomatidae)吸虫的统称,种类较多,包括棘口亚科(Echinostomatinae)、棘隙亚科(Echinochasminae) 等11 个亚科,棘口属(Echinostoma)、棘隙属(Echinochasmus)、似颈属(Isthmiophora)等51 个属的虫体600 多种,这类吸虫主要寄生于哺乳类和鸟禽类,大约有20 多种可以感染人[1-3]。日本棘隙吸虫(Echinochasmus japonicus)和圆圃似颈吸虫(Isthmiophora hortensis,圆圃棘口吸虫(Echinostoma hortensis)的同物异名就是其中重要的种类。日本棘隙吸虫主要终末宿主为犬和猫,还感染鸡、鸭等家禽及白鹭等野生鸟类,鼠类和食虫动物也可成为其自然终末宿主,淡水螺和淡水鱼为其第一中间宿主和补充宿主,主要流行于中国、日本、韩国、科威特、老挝、俄罗斯、泰国和越南等地[3-7]。圆圃似颈吸虫的终末宿主为犬和猫,猪和鼠类也有感染的报道,第一中间宿主也是淡水螺,泥鳅和蛙为其第二中间宿主,主要流行于中国、日本、韩国[3-4,8-10]。两种虫体均寄生于宿主的肠道中。动物是因采食生的淡水鱼蛙而获感染,感染后的主要症状集中在消化系统,如腹泻、腹痛、食欲不振等,重度感染可见肠黏膜出血、肠炎,导致脱水,严重者可引起死亡,对犬猫饲养影响很大[1-3]。同时,两种吸虫均可以感染人,具有重要的公共卫生意义[3-4]。
寄生虫的准确鉴定是寄生虫病防控和科学研究的基础,过去对寄生虫的分类鉴定主要依赖于形态学方法,但这种方法对于形态相似的虫体存在一定的局限性。棘口吸虫不仅种类繁多,而且很多种类形态相似,同一属内,甚至同一亚科内的虫体也很相似。头棘的数量和排列是棘口科吸虫鉴定的一种重要标志,然而同一属中的不同种的头棘数量也不尽相同,不同属间头棘的数量也有相同的,这就大大地增加了鉴定的难度。可以这样讲,棘口科吸虫是吸虫中最难准确鉴定的类群之一,然而分子生物学技术的出现解决了这一难题。试验以采自黑龙江省自然感染犬体小肠内的两种棘口吸虫为研究对象,用PCR 方法扩增其核糖体DNA 中的内转录间隔区(Internal transcribed spacer,ITS)序列,并与GenBank 上公布的相关棘口吸虫ITS 序列进行对比分析和进化分析,鉴定吸虫的种类。
1 材料与方法
1.1 材料
1.1.1 虫体来源
虫体来自黑龙江某地自然死亡的犬只,剖检时在小肠内检出虫体8 条,清洗后保存于70%的乙醇中,于-20 ℃保存备用。
1.1.2 实验材料
显微镜,载玻片,盖玻片,移液管,镊子,挑针,DNA 提取试剂盒(TIANamp Genomic DNA Kit),Ex Taq DNA 聚合酶(TaKaRa),dNTPs(TaKaRa),r Taq DNA 聚合酶(TaKaRa),pMD18-T 载体(TaKaRa),大肠杆菌DH5α 感受态细胞(TaKaRa),DNA Marker(2000),1×TAE,蛋白酶K,生理盐水,75%乙醇,琼脂糖,溴化乙锭,DNA 凝胶回收试剂盒(AxyPrep DNA Gel Extraction Kit),甘油,冰乙酸,去离子水等实验常规试剂。
1.2 方法
1.2.1 形态学初步观察
因犬死亡时间较长,送检较晚,虫体破损较为严重,无法制作装片进行仔细观察。于是将收集到的虫体用蒸馏水反复洗涤后,直接置显微镜下进行观察。依据虫体大小和外部形态,初步断定是两种吸虫,分别命名为吸虫A 和吸虫B。在不同倍数不断调整虫体位置来观察虫体外观及内部各器官的形态特征。
1.2.2 总DNA 提取
将两种吸虫清洗后,用TIANamp Genomic DNA Kit(TIANGEN)试剂盒严格按照说明书进行DNA 提取,提取后DNA 分装置于-20 ℃冰箱中保存,用于下一步PCR 扩增。
1.2.3 目的基因的扩增
以两种虫体DNA 为模板,PCR 扩增虫体ITS 序列,使用Gasser 等[11]设计的通用引物,上游引物NC5:5′-GTAGGTGAACCTGCGGAAGGATCATT-3′,下游引物NC2:5′-TTAGTTTCTTTTCCTCCGCT-3′。引物由哈尔滨擎科生物有限公司合成。PCR 反应体系为25 μL,1 μL DNA 模板,2.5 μL Ex Taq Buffer,2 μL dNTP mixture,0.5 μL 引物和0.2 μL Ex Taq DNA 酶。具体的扩增条件为:94 ℃预变性5 min;进入循环:98 ℃变性30 s,退火温度55 ℃30 s,72 ℃延伸1 min 30 s,共35 个循环;最后72 ℃延伸5 min。扩增出的产物通过琼脂糖凝胶电泳检测,凝胶成像系统拍照。切下目的条带后用DNA 琼脂糖凝胶回收试剂盒进行回收。然后将回收的DNA 产物连接到T-easy 载体上,经过无抗LB 培养后,涂在AMP+的固体LB 培养基上,克隆所得阳性质粒送到哈尔滨擎科科技有限公司进行测序。
1.2.4 序列分析与进化关系分析
测序得到的序列经拼接后,应用DNAStar 软件中的MegAlign 与棘口科的相关虫体进行同源性分析。利用MEGA X 软件,基于rDNA ITS2 序列,以肝片吸虫为外群,采用最大简约法(Maximum Parsimony,MP)将研究的两种虫体与棘口科13 属29 种吸虫构建进化树,来探讨这两种吸虫与其它棘口吸虫之间的亲缘关系。
2 结果与分析
2.1 虫体形态学初步观察
吸虫A:吸虫A 一共5 条虫体,虫体扁平,外观整体呈瓶状,有口腹两吸盘。体长1.03~1.04 mm,体最宽0.33~0.38 mm。有头部24 棘,背部中间无棘,左右各12 枚。口吸盘位于虫体前端,腹吸盘位于虫体前1/2 处。睾丸呈椭圆形,前后排列于虫体后1/3 处。卵巢看不清楚。参照相关文献[1]初步鉴定为棘口科棘隙属吸虫。
吸虫B:吸虫B 一共3 条虫体,虫体呈长叶形,有口腹两吸盘。体长8.6~9.1 mm,体最宽1.2~1.3 mm。有头棘27 枚,8 枚腹角棘平分,左右各4 枚。口吸盘位于虫体前端,腹吸盘在虫体前1/5 处,大于口吸盘。睾丸呈类椭圆形,前后排列于虫体中部。卵巢形状不清,卵黄腺分布于虫体两侧数量较多。参照相关文献[1,9]初步鉴定为棘口科似颈属吸虫。
2.2 目的基因的扩增
成功扩增两种虫体的ITS 序列,分别在1 300 bp和1 000 bp 左右出现明显条带,大小在目的基因范围之内,条带单一且明亮(图1)。
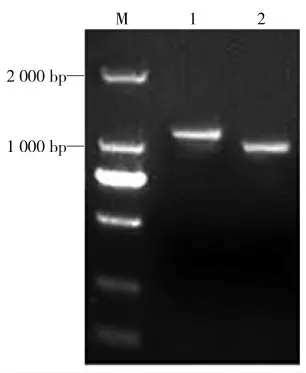
图1 两种吸虫ITS 序列的扩增结果Fig.1 The amplification of ITS sequence of two trematodes
2.3 序列分析与进化分析
经鉴定的阳性质粒送至哈尔滨擎科生物有限公司进行测序,拼接后得到长度分别为1 233 bp 和1 046 bp 的ITS 序列。吸虫A 的ITS1 长度为386 bp,A+T 含量为49.48%,5.8 S 长度为156 bp,A+T 含量为45.51%,ITS2 长度为691 bp,A+T 含量为47.47%,ITS 全序列A+T 含量为47.85%。经MegAlign 同源性序列分析比较发现,该虫体与GenBank 上已经发表棘口科冠孔属(Stephanoprora)、棘口属(Echinostoma)、棘缘属(Echinoparyphium)、真缘属(Euparyphium)和似颈属(Isthmiophora) 的同源性较低,在76.4%~82.9%之间;与同属虫体同源性较高,与Echinochasmus coaxatus 同源性为96.4%,而与来自越南的日本棘隙吸虫(KT873314.1)同源性高达99.5%。吸虫B 的ITS1、5.8 S 和ITS2 的大小分别是444、162 bp 和440 bp,A+T 含量分别是46.62%、47.53%和50.00%,ITS 全序列A+T 含量为48.18%。同源性分析显示与冠孔属、棘缘属、真缘属和棘隙属的同源性在74.9%~89.4%之间,与棘口属的Echinostoma caproni 的同源性仅为84.8%,但与来自黑龙江淡水鱼圆圃棘口吸虫囊蚴(KX832896.1) 和来自日本浣熊的圆圃似颈吸虫(AB189982.1)ITS2 序列同源性均为100%。
基于ITS2 序列,棘口科13 属29 种吸虫构建的进化树显示,整个棘口科吸虫形成两个大的分支,棘隙属虫体与冠孔属虫体形成一个大分支,棘口属及其它属吸虫形成另一大分支。在棘隙属与冠孔属这一大分支中,棘隙吸虫又单独形成一个小的分支,实验吸虫A 与来自越南的5 条日本棘隙吸虫聚集在一起,亲缘关系较另一种棘隙吸虫(Echinochasmus coaxatus)近。值得注意的是,吸虫B 并未与除圆圃棘口吸虫外的其他棘口属吸虫聚集在一起,而是与圆圃棘口吸虫、Isthmiophora hortensis 聚集在一起,与獾似颈吸虫(Isthmiophora melis)形成一独立分支(图2)。

图2 基于ITS2 序列构建的进化树Fig.2 The phylogenetic tree based on ITS2 sequences
因此,基于序列分析和进化分析,鉴定两种吸虫分别为日本棘隙吸虫和圆圃似颈吸虫。
3 讨论
日本棘隙吸虫和圆圃似颈吸虫感染的动物主要是犬和猫,因这些动物不是影响畜牧业经济的主要动物,所以人们关注和重视的程度不高。然而感染的犬和猫是人类感染的重要传染源,在疫病的流行中起到非常重要的作用。人是因为食用生的或未煮熟的淡水鱼和蛙类(蛙类仅为圆圃似颈吸虫的中间宿主)而感染。目前,世界上很多国家有人感染的病例报道,如中国、韩国、日本、泰国、老挝、越南等[3-7,12-15],可以看出这些国家主要集中在东南亚,这可能与人们的生活饮食习惯有关,这些地区的人们都吃生淡水鱼的习惯。在我国,虽然同样因吃生鱼而感染华支睾吸虫的患者广泛分布于27 省区[16],但日本棘隙吸虫仅在福建、广东、广西、江苏、安徽等地有人感染的报道[5,12,17-20],圆圃似颈吸虫仅在广西、辽宁、贵州和黑龙江有报道[21-24]。分析可能是由于该虫体寄生于小肠,虫体较小,人体感染强度通常又比较低,症状比较轻微,而且很少有人在没有寄生虫感染迹象时进行寄生虫虫卵检查,即使感染了,也很可能会被遗漏,估计实际感染的省份和病例远不止这些。虽然在黑龙江省多地犬小肠内检出日本棘隙吸虫[25],同时在鱼吸虫囊蚴检查时,也发现了大量日本棘隙吸虫囊蚴,而且感染鱼的种类多,地理分布较广,黑龙江省却还未见人感染的报道。尽管成年人少量感染棘口吸虫不能造成直接死亡,但由于头棘造成肠道损伤,易继发细菌感染,再加之毒素作用,对人的危害也不容忽视。幼儿和儿童大量感染可产生严重后果,江苏有一17 月龄患儿因寄生207 条日本棘隙吸虫引起长期腹泻,导致患儿重度营养不良并发念球菌感染,全身衰竭而死[19]。桂林也有一例5 岁男孩因大量感染日本棘口吸虫而死亡的报道[17]。因此,建议对棘口吸虫应足够重视,在条件允许情况下,对于有吃生鱼蛙史的人们,特别是采取生吃泥鳅偏方治病者,应定期进行粪便检查,发现虫卵,及时驱虫,以保证身体健康。
由于传统形态学鉴定方法的局限性,以核糖体和线粒体为标记基因来鉴定虫体得到了广泛应用。其中应用最多的就是ITS 序列,ITS 序列是18S 和28S 之间的区域片段,包括内转录间隔区1(ITS1)和内转录间隔区2(ITS2),核糖体DNA 中的ITS1 和ITS2片段是非编码区,不转录成RNA,进化选择压力小,相对变化较大,在种间有较大的差异,所以可以作为鉴定近缘种与种群的系统进化关系的标记基因[26]。Huang 等[27]在对来自不同地区的肝片吸虫和大片吸虫ITS 序列进行分析发现了一种介于两者之间的“中间型”,目前“中间型”片形吸虫已得到学术界的认可;高远等[28]通过对鼻形杯环线虫ITS 序列进行扩增和比对分析,准确地鉴定了该虫体;羊仰口线虫和牛仰口线虫两者形态比较相似,特别是雌虫,为了从分子水平上鉴别两种虫体,Wang 等[29]使用限制性内切酶Nde I 对两种线虫核糖体ITS 进行酶切,结果羊仰口线虫被切成两段,而牛仰口线虫不变,建立了区分两种线虫的PCR-RFLP 方法。ITS 序列不仅可以鉴定成虫,还可以鉴定吸虫囊蚴,华支睾吸虫和东方次睾吸虫囊蚴均寄生于鱼的肌肉,形态十分相似,很难通过形态学加以区分,我们团队以ITS 为标记基因建立了区分两种吸虫囊蚴的PCR-RFLP 方法,并获批了发明专利[30]。
日本棘隙吸虫与藐小棘隙吸虫等同属的大部分吸虫均为24 个头棘,大小相似;圆圃似颈吸虫和獾似颈吸虫(Isthmiophora melis)、真缘属的隐棘真缘吸虫(Euparyphium inerme)均有27 枚头棘,大小也差不多,形态学准确鉴定需要丰富的经验,难度较大,特别是当虫体形态受损时,形态观察则更加困难,因此采取PCR 方法扩增了待检虫体的ITS 序列,并与相关序列比较分析,同时构建了分子进化树来鉴定这两种虫体。序列分析发现,吸虫A 与GenBank 公布的日本棘隙吸虫(KT873311.1)同源性为99.5%。基于ITS 序列构建的棘口科吸虫进化树显示,吸虫A 与棘隙属吸虫一起,与冠孔属吸虫和棘隙属处于一个大分支上。在棘隙属吸虫分支中,试验虫体与其来自越南的5 条日本棘隙吸虫ITS 序列聚在一起,可以准确地鉴定该虫体为日本棘隙吸虫。关于圆圃似颈吸虫的分类一直有争议,该虫体最初由日本学者在1926年发现并命名为圆圃棘口吸虫[31]。2002 年,Kostadinova 和Gibson 在对似颈属和真缘属进行分类研究时,确定了这两属的有效性,并对相关虫体进行重新分类。通过对圆圃棘口吸虫的详细观察,认为该虫体的子宫区域、睾丸位置,头棘27 枚和不同种类棘的大小等特征更符合似颈属特征,正式将其从棘口属移至似颈属,并将其重命名为圆圃似颈吸虫(Isthmiophora hortensis)[32],该种名已得到业界的认可,成为有效种。但我国有的学者还使用圆圃棘口吸虫这一名称[24,33]。研究的序列分析也发现,检出的吸虫B与棘口属吸虫Echinostoma caproni 的同源性仅为84.8%,而与獾似颈吸虫同源性为95.4%,与圆圃似颈吸虫和圆圃棘口吸虫同源性为100%。进化分析显示吸虫B 没有和棘口属的吸虫在一起,而是与圆圃似颈吸虫处与同一分支。因此基于对吸虫核糖体ITS序列的研究也支持圆圃棘口吸虫应为圆圃似颈吸虫,研究所获得来自于犬的虫体为圆圃似颈吸虫,以前我们团队在GenBank 上公布的来自于犬的和鱼囊蚴的相关序列应为圆圃似颈吸虫序列,同时建议以后学者在发表学术论文和学术交流时采用圆圃似颈吸虫来代替圆圃棘口吸虫这一名称。研究再次证明了ITS 是寄生虫分子鉴定的重要靶标。
猜你喜欢
动物医学进展(2024年5期)2024-05-09 08:09:50
锦绣·下旬刊(2022年1期)2022-05-16 07:30:45
畜牧兽医科技信息(2021年2期)2021-03-29 10:40:30
中国工作犬业(2021年3期)2021-03-27 15:10:41
寄生虫与医学昆虫学报(2021年4期)2021-02-18 08:03:14
中国动物传染病学报(2020年5期)2020-10-17 06:56:46
兽医导刊(2019年1期)2019-02-21 01:14:28
中国人兽共患病学报(2017年9期)2017-10-09 04:58:27
科学中国人(2017年14期)2017-05-19 00:48:03
中国人兽共患病学报(2015年9期)2015-05-09 01:03:45
