
百合科植物psbA基因密码子偏好性分析
2020-12-21 07:24倪典墨李冉郡郭忠静高玲玲周嘉裕
生物学杂志 2020年6期
倪典墨, 李冉郡, 郭忠静, 高玲玲, 周嘉裕, 廖 海
(西南交通大学 生命科学与工程学院, 成都 610031)
百合科(Liliaceae)为单子叶植物中的一大科,广布于温带及亚热带地区,在全世界约有250属3500种[1]。我国百合科植物约有60属,600多种,主要分布在西南地区[1],其在药用、食用、园林绿化及工业等领域有较大的开发利用潜力。例如,川贝母是贝母属(Fritillaria)的代表植物,其药用历史首载于《神农本草经》,其鳞茎具有润肺、止咳与化痰等功效,被誉为“止咳神药”[2]。百合属(Lilium)植物百合(Liliumbrownii)不仅能药用和食用,而且因其花朵鲜艳、花色丰富,亦被视为庭园观赏和绿化珍品[3]。芦荟属(Aloe)植物芦荟(Aloevera),既是优良的观赏花卉,还能吸收苯等有毒物质,起到净化空气的作用[4],其在药理上具有杀菌抗炎、健胃下泄、强心活血、解毒化瘀、免疫和再生等作用[5]。黄精属(Polygonatum)植物黄精(Polygonatumsibiricum),主要用于药物和保健品,有养阴生津、补脾润肺、止虚咳、治消渴和降低胆固醇等作用,对治疗冠心病、脑动脉硬化疾病也有较好的疗效[6]。
然而,百合科植物在栽培及生长过程中受到了多种生物及非生物因素的影响:第一,地老虎、蚜虫等大规模虫害的出现,极易造成植株死亡[7];第二,贝母的耐热性弱导致贝母出现“倒苗”,严重影响产量[8];第三,百合科植物的生长受到黑腐病、灰霉病、干腐病与软腐病等的严重威胁[9];第四,百合科药用植物活性成分含量偏低,这也限制了该科植物药用价值的提升[10]。基因工程技术是改良植物性状、提高百合科植物抵抗外界不利因素能力与活性成分含量的一种有效手段。叶绿体因其表达蛋白稳定性较高,同时能将转化基因的扩散控制在最低程度[11],使之成为植物基因工程的理想载体。目前,叶绿体转基因技术在提高植物抗性及改良性状方面已取得较大进展,为提高植物的抗病虫害以及对增强干旱、盐碱等逆境的抗性开辟了一条更为有效的途径[12]。并且,叶绿体载体也大量应用于疫苗抗原、激素及抗体等外源蛋白质的生产[13]。相比较核基因组,叶绿体基因组在结构与序列上相对保守,组成及结构更简单,具有与核基因明显不同的密码子偏好性,因此为提高外源基因在叶绿体中的表达效率,研究叶绿体基因的密码子偏好性势在必行[14]。psbA基因是叶绿体中的重要光调控基因,其编码光合系统II反应中心的Dl蛋白,在光合作用中发挥光电子转运的功能[15]。psbA基因在叶绿体基因工程中发挥了重要作用,其启动子(PpsbA)与终止子(TpsbA)经常作为外源基因的启动子与终止子使用,并且psbA-trnK间隔区常常作为外源基因的插入位点[16]。
为了更好地设计叶绿体高效表达载体,进一步利用叶绿体基因工程技术改良百合科植物重要性状,分析百合科植物psbA基因密码子偏好性十分必要。本文以60种百合科植物的psbA基因为研究对象,分析该基因的碱基组成特点以及密码子的偏好性,以期对叶绿体基因工程研究提供理论参考。
1 材料与方法
1.1 序列来源
实验室近期完成了暗紫贝母(Fritillariaunibracteata)的叶绿体基因组测序,从中获得了暗紫贝母的psbA基因序列(登录号MN727396)。同时,登录NCBI数据库(http://www.ncbi.nlm.nih.gov/),下载来自百合科百合属、贝母属、黄精属与葱属等7个不同属来源的59种植物的psbA基因序列,具体序列信息见表1。
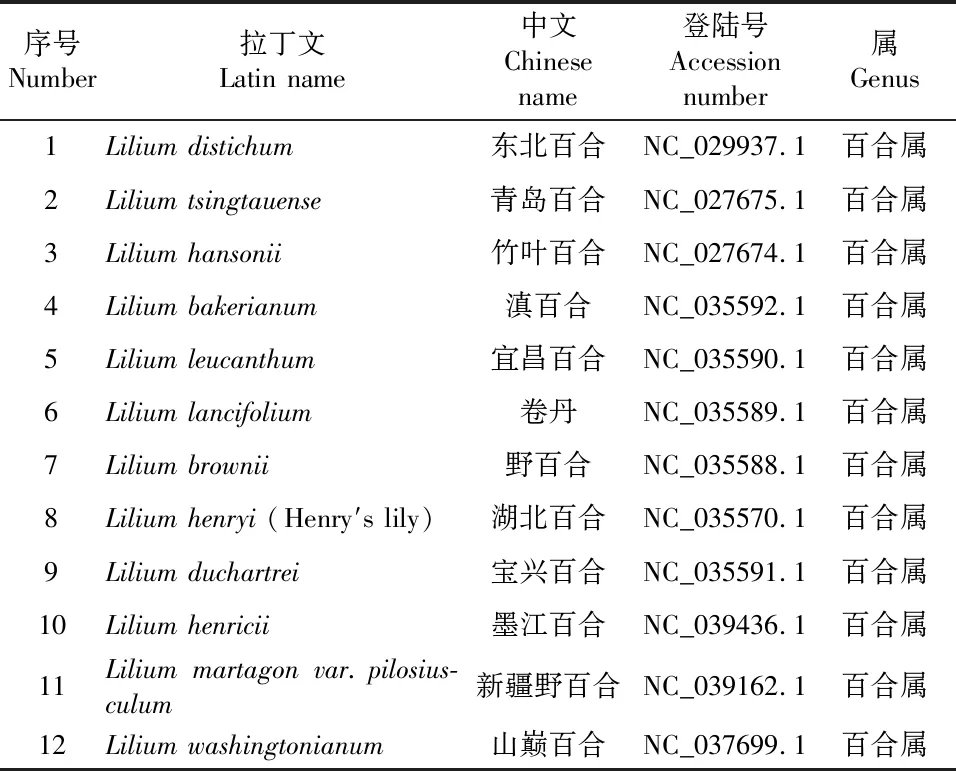
表1 60种百合科植物psbA基因序列

续表1 (Table 1 Continued)
1.2 方法
1.2.1psbA基因同义密码子偏好性研究
密码子偏好性的相关参数包括:相对密码子使用度RSCU(Relative synonymous codon usage);密码子适应指数CAI(Codon adaptation index);同义密码子第3位碱基的GC含量GC3s;同义密码子GC含量;有效密码子数ENc(Effective number of codons)。
首先,利用CodonW程序分析60种百合科植物psbA序列的RSCU、CAI、GC3s和GC含量;其次,利用CUSP和CHIPS在线程序计算各psbA序列的密码子出现频率和ENc值。
1.2.2 密码子偏好性与CDS的聚类分析
以60种百合科植物、烟草、拟南芥的密码子RSCU值为基础,利用SPSS软件进行密码子偏好性聚类分析。利用ClustalW软件对psbA基因的 CDS序列进行序列比对,再用DNAMAN软件将比对结果,利用邻近法构建系统进化树。
2 结果与分析
2.1 百合科植物psbA基因密码子偏好性分析
RSCU是指对于某一特定的密码子在编码对应氨基酸的同义密码子间的相对概率。若RSCU>1,则说明该密码子的使用频率高;若RSCU<1,则说明该密码子的使用频率低;若RSCU=1,则说明该密码子的使用不存在偏好性[17]。60种百合科植物的psbA基因中,有26个密码子具有偏好性,分别为GCU、GCC、UGC、GAU、GAA、UUC、GGU、GGA、CAU、AUU、AAA、UUG、CUC、AAU、CCA、CAA、AGA、AGG、AGU、AGC、ACC、ACG、GUU、UAC、UAA与UAG。其中GCC、GAU、GAA、UUC、CAU、CUC、CCA、CAA、AGA、AGG、AGC与ACC的偏好性极强,它们的RSCU值不小于2;AUG与UGG密码子的RSCU值为1,不具有偏好性。RSCU=0的密码子有13个,分别为GAC、GAG、UUU、CAC、CUU、CUA、CCG、CAG、CGU、CGC、CGA、CGG和UCA;RSCU<0.5的密码子有10个,分别为GCA、GCG、UGU、UUA、CCU、UCU、UCG、ACA、GUC和UGA;百合科植物倾向于UAA和UAG作为终止密码子(图1)。
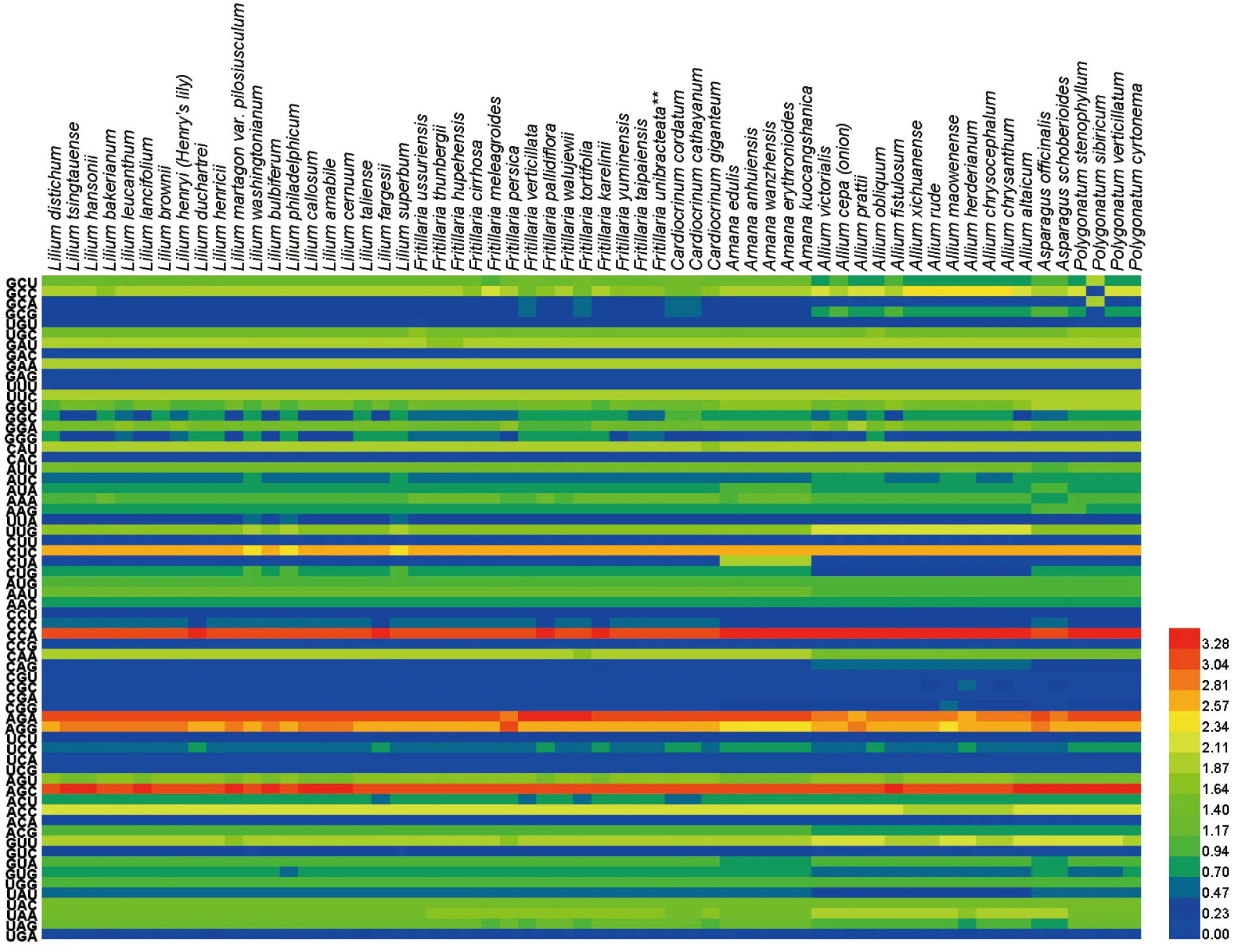
图1 60种百合科植物psbA基因的RSCU热图
2.2 ENc、CAI、GC3s和GC的数据分析
ENc值用来衡量同义密码子的偏好使用程度,介于20~61,以35为临界值,越靠近20,则表明该密码子的偏好性越强[18]。CAI值常用来衡量基因表达强度,其大小0~1,其值越高,则表明基因表达强度越高[19]。GC3s可以反映密码子第3位的使用偏好性,若GC3s大于0.5,则密码子偏向于G/C结尾。GC含量反映G/C碱基对的使用偏好性,若GC大于0.5,表明G/C碱基对的偏好性强,反之表示A/U的偏好性强。
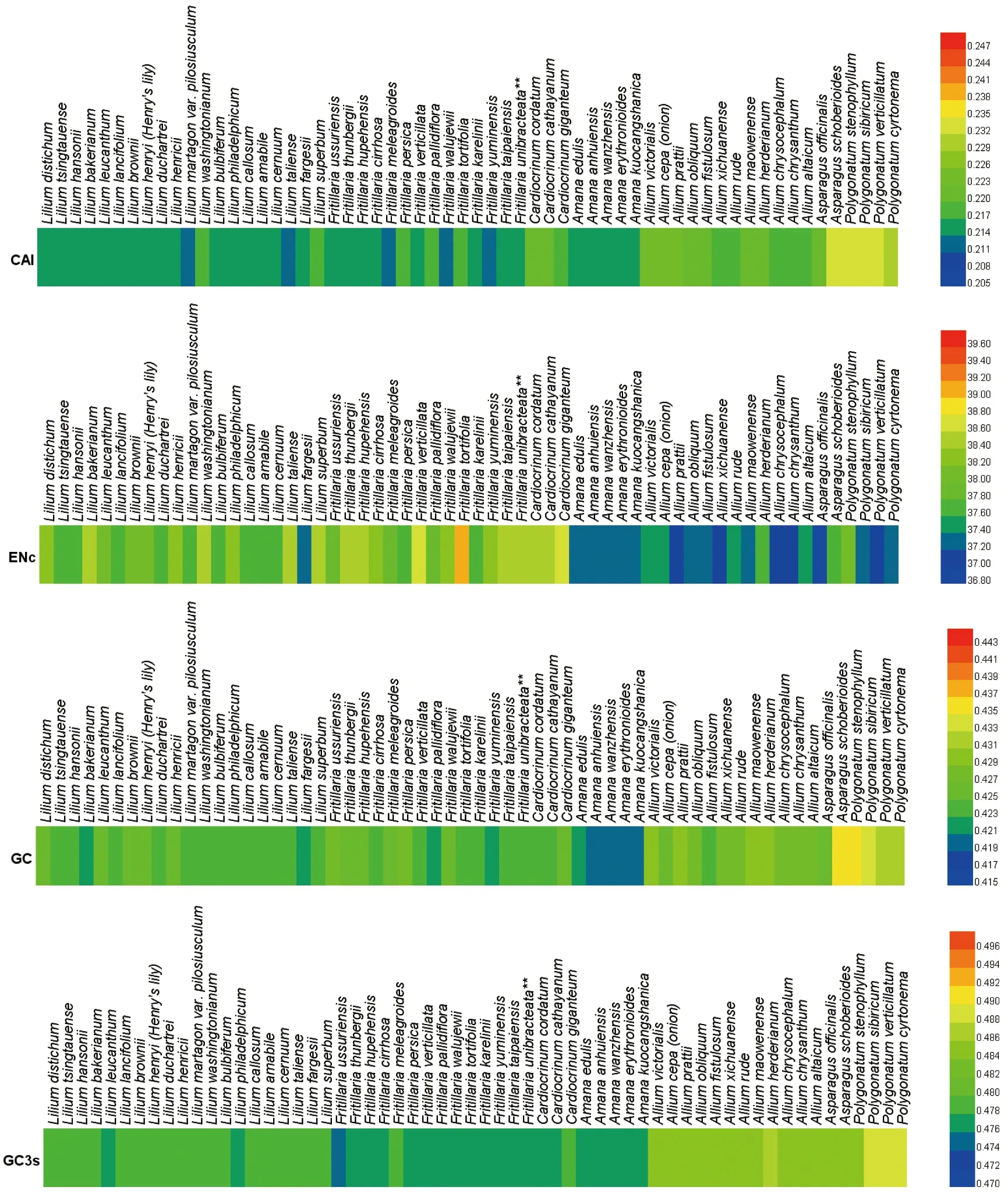
图2 psbA基因CAI、ENc、GC与GC3s热图
由图2可知:百合科植物psbA基因的ENc值超过35(范围为36.97~39.06),平均值为37.84,标准偏差为0.5147,说明百合科植物psbA基因的密码子偏好性整体较弱;CAI值为0.212~0.233,平均值为0.218,标准偏差为0.005 19,说明密码子的表达水平偏低[9]。由此,可以推断百合科植物中psbA基因的表达水平较低,D1蛋白含量低,成为光反应的限制性因素之一。进一步分析psbA基因的碱基组成发现,G+C含量介于42.1%~43.6%,平均值为42.5%,标准偏差为0.003 28,说明百合科植物更加偏向A与U碱基。以G+C为结尾的密码子含量(GC3s)介于47.5%~48.9%,平均值为48.0%,标准偏差为0.003 71,尽管GC3s低于50%,但仍然超过G+C的总体含量。
2.3 ENc-plot分析
ENc-plot分析以ENc为纵坐标、GC3s为横坐标作图,绘图中的标准曲线表示密码子偏好性仅由碱基组成决定基因位置,标准曲线计算方法[20]如下:ENc= 2 +GC3s+ 29/[GC3s2+(1-GC3s)2]。由此可构建GC3s散点图和ENc-GC3s期望曲线,各点离期望曲线的远近可反映密码子偏好性形成的原因是碱基突变或是自然选择[21]。若某一基因的密码子偏好性形成受突变影响较大时,其ENc-GC3s点将分布于期望曲线附近;若其受自然选择影响较大时,则会分布在偏离期望曲线较远的位置。利用Origin 8绘制的GC3s散点图和ENc-GC3s期望曲线可知,自然选择是影响百合科psbA密码子偏好性的主要原因(图3)。
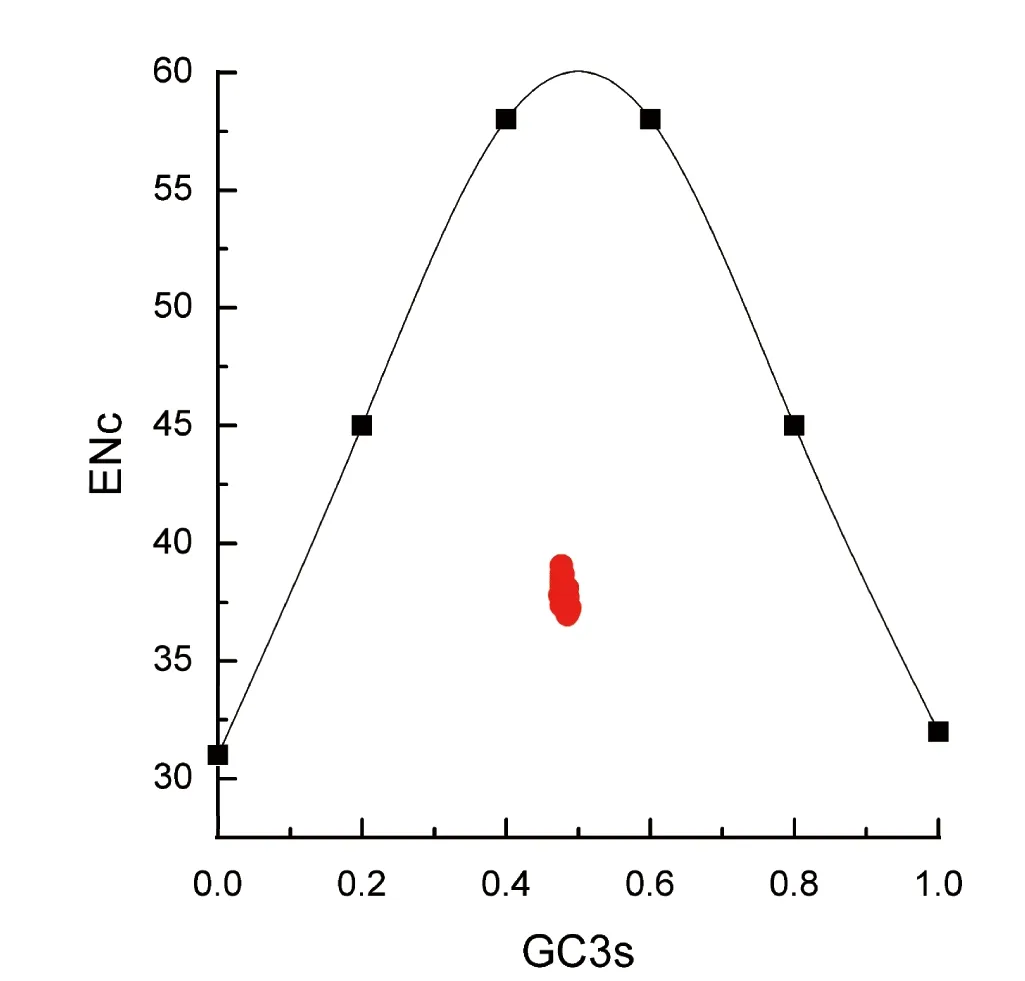
图3 psbA基因的ENc-plot分析
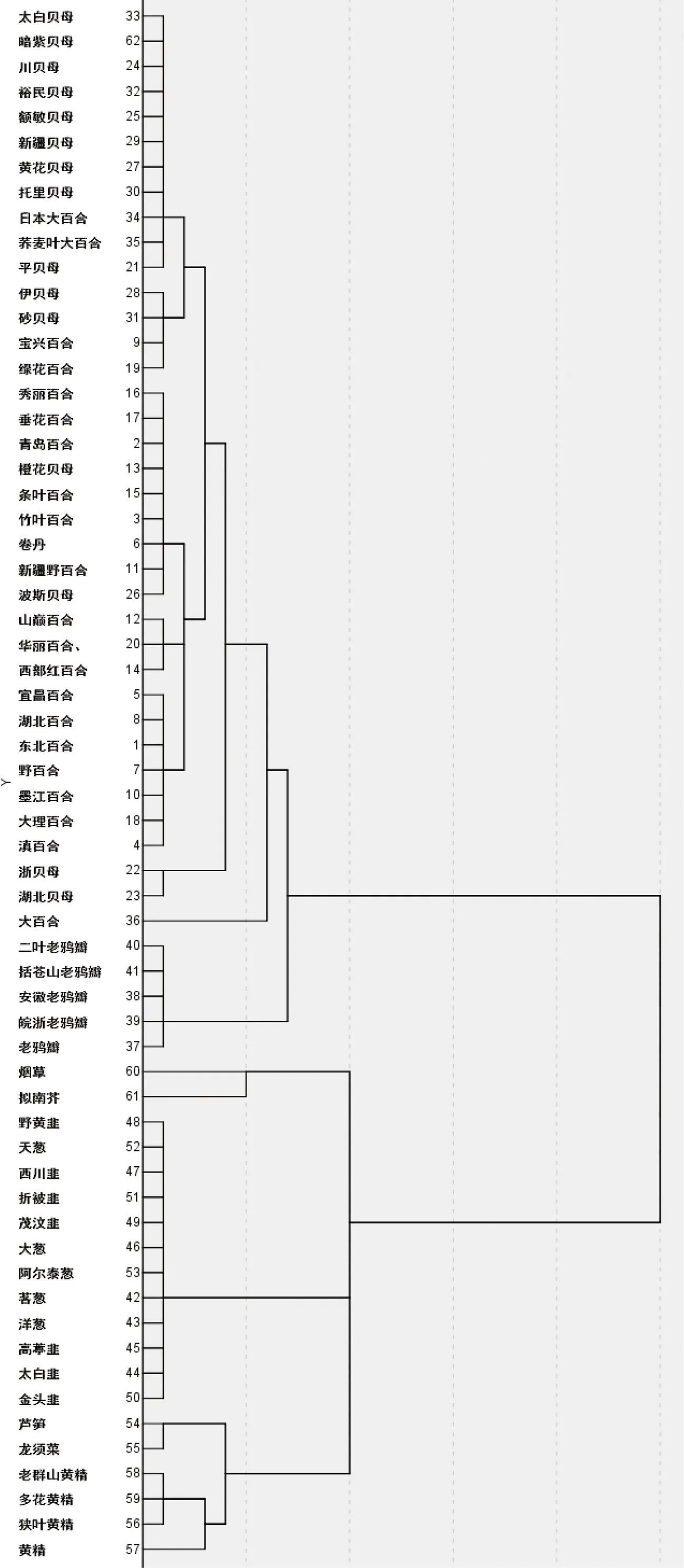
图4 psbA基因RSCU值聚类树状图
2.4 psbA密码子偏好性聚类分析
依据不同植物密码子的RSCU值进行聚类(图4),能够将所有百合科植物分为两簇,其中葱属、黄精属与天门冬属聚为第一簇,百合属、贝母属、大百合属和老鸦瓣属为第二簇。来自葱属的12种植物具有接近的密码子偏好性,然而,基于RSCU的聚类并不能反映出所有百合科植物之间的亲缘关系,比如:日本大白合、荞麦叶大白合与贝母属植物聚为一类;同为大百合属的3种植物未能单独聚为一类,其中大百合单独形成一个分支;百合属与贝母属的多种植物在聚类树中相互交叉,形成复杂的混合分布。尽管烟草与拟南芥不属于百合科,但却与葱属、黄精属与天门冬属同处在第一簇,与葱属有相近的密码子偏好性。
2.5 基于CDS序列的NJ进化树分析
基于CDS的系统进化树更接近这60种植物的真实系统分类,在NJ进化树(图5)中烟草、拟南芥与百合科植物形成2个不同的分支。百合科植物中,贝母属、百合属、大百合属与老鸦瓣属等聚为一簇,而黄精属与天门冬属等聚为一簇。具有相近亲缘关系的植物在NJ树中相对接近,表明psbA基因CDS的差异大小与物种的亲缘关系远近有较密切的联系。比如:来自贝母属的所用植物均集中在进化树中的一个分支;来自大百合属的3种植物集中在一个分支。
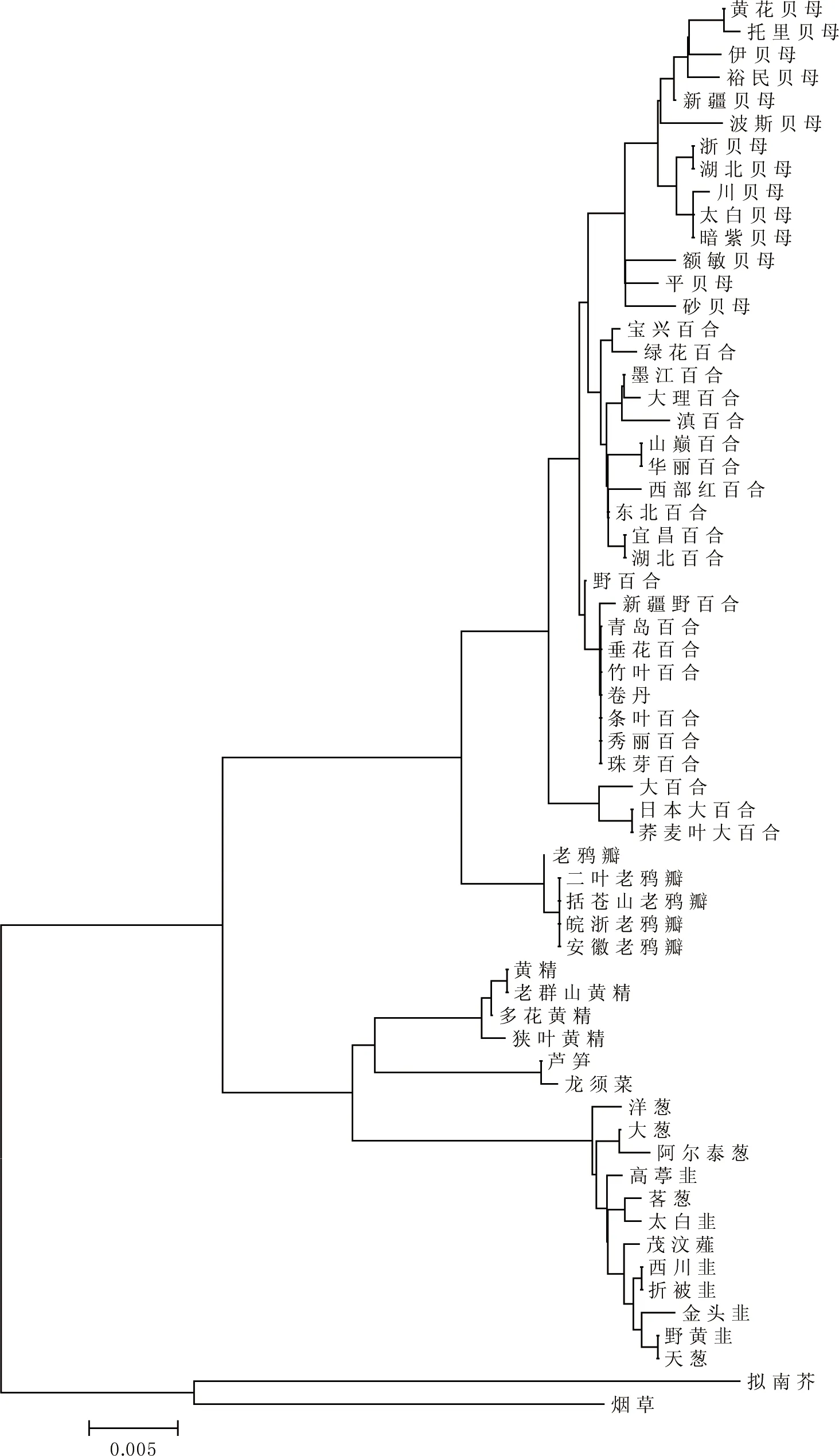
图5 psbA基因CDS的NJ进化树
3 讨论与结论
RSCU是衡量密码子偏好性的重要指标。对60种百合科植物psbA基因密码子的偏好性分析,发现有26种存在偏好性的密码子,其中有12个表现出较强的偏好性。有13个密码子的RSCU值均显示为0,另有10种密码子在60种百合科植物中应用较少(RSCU<0.5),表明在百合科叶绿体基因工程中,应尽量避免这23种使用率极低的密码子,而优选26种存在偏好性的密码子。从psbA基因的ENc值与碱基组成上看:百合科植物的ENc值均大于35,偏好性整体偏低;GC平均含量为42.5%,尽管GC结尾的密码子含量(48.0%)高于GC平均含量,但仍然倾向以A/U结尾的密码子,这与多数单子叶植物,如水稻(GC3s 49.5%)、小麦(GC3s 48.9%)、玉米(GC3s 49.7%)和高粱(GC3s 49.5%)等物种的密码子结尾特点相一致[22]。在26个具有偏好性的密码子中,有16个是以A/U结尾,这也表明百合科植物psbA基因偏向以A/U结尾的密码子。一般认为,单子叶植物偏好以G/C结尾,双子叶植物偏好以A/U结尾[23]。然而,本研究发现百合科与禾本科植物的GC3s均小于且接近50%,表明若开展百合科与禾本科叶绿体基因工程时,密码子倾向于A/U结尾。CAI值反映百合科植物中psbA基因的表达水平较低,表明若对百合科植物开展基因改造以提高光合效率,psbA基因可能是一个合适的候选靶基因。比如通过调控psbA基因的表达,能够显著提高烟草的光合效率及生物量积累[24]。
突变和选择是密码子偏好形成的主要原因。ENc-plot分析结果表明60种百合科植物psbA基因的ENc-GC3s点均远离标准曲线,基因的实际ENc值远小于理论ENc值,表明psbA基因密码子偏好性主要受到自然选择的作用。
基于RSCU的聚类分析与基于CDS的进化分析结果可能存在一定差异,这种现象在前人[22]的研究中也曾出现。在本研究中,图4和图5的2个聚类图也不完全一致:图4未能将百合属和贝母属的植物区分开,而图5则较好地反映了植物之间的进化关系,说明基因序列比密码子偏好性更适应于做进化分析使用。分析导致该现象的可能原因是:psbA基因的适应性进化主要源于负向选择[25],非同义密码子的突变往往会导致psbA功能的改变,因此基因序列受到的选择压力更大;psbA基因的功能多样性,植物在长期进化过程中,由于叶绿体基因与核基因组、原生噬藻体与宿主(蓝藻)之间[26]的物质与信息交流,导致psbA基因虽然表现出一定的种族特异性,但通过单基因的密码子偏好性聚类仍然难以反映出正确的亲缘关系。
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-21
艺术品鉴(2021年12期)2021-05-25
生物学通报(2020年11期)2020-10-22
少儿科技(2019年9期)2019-09-10
发明与创新·中学生(2019年6期)2019-06-26
基层中医药(2018年9期)2018-11-09
中成药(2018年7期)2018-08-04
中成药(2017年5期)2017-06-13
中国野生植物资源(2014年4期)2014-03-29
中国烟草学报(2012年2期)2012-04-09
