
不同浓度氮处理对毛果杨天冬氨酸转氨酶酶活性及基因表达的影响
2020-12-21 07:24闵杰,曹丹,韩梅
生物学杂志 2020年6期
闵 杰, 曹 丹, 韩 梅
(南京林业大学 生物与环境学院 南方现代林业协同创新中心, 南京 210037)
杨树在全球共有100多种,是世界上分布范围最广、适应性最强的树种之一[1]。相比于其他速生丰产林树种, 杨树有着更高的养分需求量,尤其是氮营养元素[2]。先前有研究表明供氮水平会调控根系生长和植物形态变化,如根系形态及构型的变化[3]、叶片形态的改变[4]、生物量在地上及地下分配的调节[5]等。氮素量的多少直接影响杨树的材积量与胸径, 在一定的氮素内,它们成正比,但氮素过高会产生抑制效果。随着氮素浓度的升高,杨树木材部分木质素含量有下降趋势[6],根系生长受到抑制,导致根冠比降低[7-8]。
从AspAT催化的生化反应和其在代谢网络所处的位置推测,AspAT在植物氮代谢通路中有非常重要的作用,包括:1)跟种子萌发过程中氮素的转移有关;2)参与营养器官的氮素代谢循环和转运;3)协调氮素从源到库的再分配;4)与种子的氮充实和成熟有关[13]。近年来,科研人员[11-12,15-16]在拟南芥等植物中对AspAT在植物体内的功能作用进行了广泛的探索,初步证明AspAT在细胞内参与氮和碳代谢平衡,对作物产量和品质的形成有重大的影响。根据对拟南芥和其他物种的序列鉴定、聚类分析,AspAT可分为两大家族:Iα和Iβ,两者序列相似度为15%左右[17]。Iα包含真核生物中的AspAT,Iβ则包括了原核型的AspAT。原核型的AspAT是双功能酶,除了具有天冬氨酸酶的功能外,还具有预苯酸氨基转化酶(PAT,prephenate aminotransferase)的功能[17]。AspAT家族基因至少分布在3个不同类型的细胞器中, 包括质体、细胞质和线粒体[11-12, 15]。
本课题组在先前研究[18]中,已在杨树中鉴定了10个编码PtAspAT蛋白的家族基因。然而,这些基因在毛果杨体内的生理功能和调控方式尚不清楚。本实验旨在通过对毛果杨幼苗设立3种不同浓度(0、2和10 mmol/L)氮处理,利用非变性胶的方式测定毛果杨中天冬氨酸转氨酶3种类型同工酶在不同组织部分的酶活性差异,同时通过实时荧光定量PCR(qPCR)研究PtAspAT家族10个基因相应的表达量变化,探讨PtAspAT在杨树缺氮、正常氮和高氮条件下的酶活性和基因表达变化规律,以期更深入地了解天冬氨酸转氨酶在杨树氮代谢过程中的分子机制,为提高杨树的氮利用率提供科学基础。
1 材料与方法
1.1 材料
采用毛果杨(Populustrichocarpa)的组培无菌苗,并使用含0.1 mg/L IBA的MS 培养基[19]进行继代培养,生长条件为:温度 25 ℃,16 h/8 h(光照/黑暗)。
1.2 方法
1.2.1 不同氮浓度培养基配制
在基本MS培养基中去除硝酸钾与硝酸铵,并添加0、1和5 mmol/L的硝酸铵,作为3种不同类型的氮梯度(0、2和10 mmol/L)培养基,对应杨树缺氮、正常氮和高氮处理[20]。
1.2.2 样本处理
从分化培养基选取高度一致的小苗,转接到MS生根培养基,待根长出1 cm左右,转入各浓度氮处理培养基进行氮处理。1个月后,收集幼叶(YL)、成熟叶(ML)、主茎(S)和根(R)等4个组织,液氮冷冻研磨并冻存于-80 ℃冰箱。
1.2.3 数据分析
在 SPSS 19.0 中,使用Kolmogorov-Smirnov检验和 Levene′s 检验对所有数据进行正态分析和方差齐性检验。采用单因素方差分析(ANOVA)进行数据比较,利用 Duncan 新复极差法检验处理间差异的显著性水平(P<0.05)。
1.2.4 总RNA提取及cDNA合成
使用植物总RNA提取试剂盒RNeasy Plant Mini Kit(Qiagen)提取RNA,取1 μg RNA使用TaKaRa的反转录试剂盒(PrimeScriptTMRT reagent Kit with gDNA Eraser)去除基因组DNA并反转录合成cDNA,-20 ℃保存备用。
1.2.5 组织特异性检测
选择毛果杨(P.trichocarpa)中UBIC基因(XM_002309363.2)作为内参基因,使用Primer3设计引物(http://primer3.ut.ee/),具体见表1。采用TB green Premix ExTaqTM(TaKaRa)试剂盒,在Applied Biosystems StepOneTM实时荧光定量PCR仪(Applied Biosystems)中进行定量PCR扩增反应。反应体系为:Premix ExTaqTM5 μL,10 μmol/L正反引物各0.2 μL,ROX reference Dye(50×)0.2 μL,模板cDNA 5 ng, 加水使反应总体积为10 μL。反应条件:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火34 s,共40个循环。实时荧光定量PCR数据采用2-ΔΔct方法计算相对表达量。
1.2.6 非变性胶测定酶活性
样本制备。参照Brauc等[21]的方法进行酶活性测定。在液氮中将样品研磨成粉末,之后加入2倍体积新鲜制备的提取液(50 mmol/L Tris-HCl pH 7.5,10%甘油,0.1%Triton X-100和1 mmol/L PMSF)。在4 ℃离心机以13 000 r/min离心10 min取上清即为酶提取液,通过Bradford 法(碧云天蛋白浓度测量试剂盒:货号P0006)测取蛋白浓度,计算量取40 μg总蛋白进行非变性电泳。

表 1 PtAspAT基因实时荧光定量PCR引物序列
电泳及染色。实验采用不连续聚丙烯酰胺凝胶(PAGE)电泳法,其中浓缩胶体积分数为4%,分离胶体积分数为7.5%。染色底物液为0.1 mol/L的 Tris-HCl (pH 7.5),0.2 mol/L L-天冬氨酸,0.2 mol/L 2-氧戊二酸,0.1 mol/L硝酸钙和5 g/L PVP-40,将凝胶放入染色液37 ℃孵育30 min,然后在fast violet blue溶液(2 mg/mL)中温育直至AspAT酶活性条带区被染成紫色带。
2 结果与分析
2.1 毛果杨PtAspAT蛋白序列比对分析结果
本课题组前期在毛果杨基因组数据库(phytozome v12.1,https://phytozome.jgi.doe.gov/pz/portal.html)中鉴定了10个PtAspAT家族基因候选成员[18]。对这10个PtAspAT基因蛋白序列进行比对分析(图 1),发现毛果杨10个PtAspAT有很高的同源性,其中有9个序列片段高度保守,它们是:1)N(V)KEYLPI;2)QS(C)LSGTGSL; 3)S(Q)PTWGNH; 4)CAHNPTG; 5)PFFDV(S)AYQGF; 6)V(M/I)AQSYS(A)KNL(M)G;7)YA(G)ERI(V)GA; 8)R(L)X2RPMY 和9)K(S)DGRISL(M)。这9个片段中每个片段至少包含1个与磷酸吡哆醛(Pyridoxal 5‘-phosphate)相互作用或与乙二酸/草酸底物结合的氨基酸残基。此外,在PtAspAT1和6中还发现对应拟南芥AtPAT (AT2G22250)基因Thr84, Lys169 和 Arg445的氨基酸残基,这些残基在特异性识别结合预苯酸(prephenate)和天冬氨酸底物方面发挥重要作用[22]。
为进一步了解PtAspAT的系统发育关系,采用MEGA 7.0的最大相似法为毛果杨10个PtAspAT蛋白序列构建系统发育进化树。如图2所示,系统进化分析显示,10个PtAspATs可聚集成两大类,即Iα和Iβ。前者包含真细菌和真核生物类型的PtAspAT,可以进一步分成3组:Iα-A,Iα-B和Iα-C。毛果杨PtAspATs的8个成员属于Iα家族,其中3个成员(PtAspAT3、4和10)属于Iα-A,3个成员(PtAspAT2、7和8)属于Iα-B, 2个成员(PtAspAT5和PtAspAT9)属于Iα-C。另外2个成员(PtAspAT1和PtAspAT6)属于Iβ家族。结合已有的文献报道[11-12,15,18],推测这些不同类型的PtAspAT蛋白至少分布在3个不同的亚细胞器中,其中Iα-A类型的PtAspAT蛋白可能定位在细胞质中,Iα-B类型的在线粒体中,而Iα-C和Iβ类型的则在叶绿体中。
2.2 不同浓度氮处理植物的表型变化
为研究PtAspAT在植物氮代谢过程中的作用,采用不同浓度的氮(0、2和10 mmol/L)对毛果杨幼苗进行缺氮、正常氮和高氮处理[20]。在持续1个月氮处理后,各氮浓度下植株表型发生明显变化。如表2所示,相对于对照组(2 mmol/L的氮浓度),缺氮(0 mmol/L的氮浓度)处理显著抑制了毛果杨生物量的积累,对地上与地下鲜重、株高和叶面积的抑制率分别达到85.588%、80.327%、49.659%和80.367%,但是根冠比显著提高,增幅率达到33.710%。随着氮浓度的提高,高氮(10 mmol/L的氮浓度)也会对毛果杨产生胁迫,相对于对照组,植株的地上鲜重、地下鲜重、株高、叶面积和根冠比均受到了显著抑制,抑制率分别达到56.765%、81.421%、28.968%、76.821%和58.757%。这些都与植物缺氮过氮表型吻合[3-5,8]。
2.3 氮处理对PtAspAT酶活性的影响
为研究PtAspAT酶活性变化规律,将不同浓度氮处理的毛果杨植物不同组织的酶粗提液进行非变性胶酶活性鉴定,结果如图3所示。根据先前的报道[16,23],非变性胶中观察到的3条主带,分别对应线粒体(Mt)、胞质(Cy)和叶绿体(Cp)中的天冬氨酸转氨酶同工酶。从图3可以看出:在正常供氮情况下线粒体PtAspAT在根和茎中活性很高,但在幼叶和成熟叶中活性很低;相反,叶绿体PtAspAT在叶片(幼叶和成熟叶)中的活性很高,茎中的中等,而在根中的活性极低。胞质PtAspAT具有与叶绿体PtAspAT同工酶类似的分布趋势,但其活性条带更弱。PtAspAT酶活性谱图清楚地显示,不同类型的PtAspAT同工酶表现出不同的特征酶谱模式,暗示它们在功能上有明显的差异。
在0、2和10 mmol/L不同氮浓度处理后,可以发现:在缺氮条件下,除成熟叶中3种PtAspAT同工酶酶活性无明显变化外,在幼叶、茎段和根中酶活性都呈现出显著的下降,这在幼叶的线粒体PtAspAT,茎和根的胞质及叶绿体的PtAspAT同工酶中体现得尤为明显;在高氮处理下,幼叶中线粒体PtAspAT酶活性显著降低,胞质和叶绿体PtAspAT活性略微下降,在成熟叶中各同工酶酶活性无明显变化,在茎段和根中,3种PtAspAT同工酶酶活性,尤其是线粒体PtAspAT活性显著上升。

表2 氮处理对毛果杨植物组培苗生长的影响

黑色框中为PtAspAT高度保守的氨基酸片段(1)~(9);“#”号代表原核型AspAT特有氨基酸

该进化树是利用MEGA7软件采用ML方法构建的,bootstrap测验采用1 000次重复,节点上的数字代表分枝的可信度。Cy:细胞质;Cp:叶绿体;Mt:线粒体
2.4 氮处理对PtAspATs基因表达量的影响
为进一步探讨PtAspAT在不同氮浓度条件下的变化规律,对PtAspATs各基因mRNA表达量进行qPCR定量分析。从图4可以看出:在缺氮条件下,除了在幼叶中的PtAspAT2,在成熟叶中的PtAspAT2、6和7以及在根系中的PtAspAT7&9表达量提高外,PtAspATs各基因在不同组织中的表达量总体上都呈现出下降的趋势;在高氮胁迫下,相对于正常氮处理,PtAspAT4、5和6在各组织中表达量略微下降,PtAspAT1在成熟叶、PtAspAT3和10在幼叶中表达量都显著提高。这些结果表明,低氮主要影响植物线粒体PtAspAT编码基因(即PtAspAT2和7)的转录,而高氮则主要影响细胞质PtAspAT编码基因(即PtAspAT3 和 10)的转录。

YL:幼叶;ML:成熟叶;S:茎;R:根;Mt:线粒体;Cy:细胞质;Cp:叶绿体
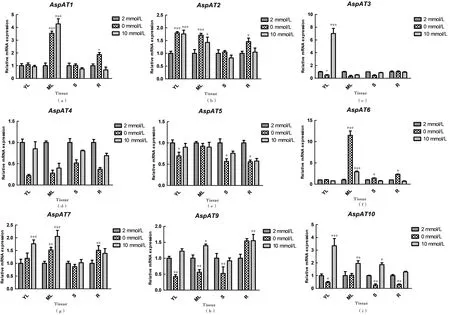
YL:幼叶;ML:成熟叶;S:茎;R:根。*表示差异显著性P<0.05;**表示P<0.01;***表示P<0.001
3 讨论与结论
氮高效利用是植物细胞、组织、器官和产量建成的关键,然而由于氮高效利用属于数量遗传性状,涉及的基因多且过程复杂(包括吸收、运输、同化和再转移等一系列过程)[24]。目前氮高效利用的生物学机制仍不清楚。天冬氨酸转氨酶催化天冬氨酸和谷氨酸相互转化,为植物氮和碳代谢通路提供原料,是植物氮吸收利用过程中的重要代谢酶[13,18],因而有必要对其在氮代谢过程中的功能和分子机制进行深入研究。

qPCR结果(图4)中,线粒体定位的PtAspAT2和7在低氮处理下表达量在根、茎、幼叶和老叶各组织总体都呈现出上升,但是线粒体PtAspAT同工酶酶活性在各组织(除老叶无明显变化)反而都显著下降(图3);在茎段中,在高氮条件下叶绿体定位的PtAspAT5和9表达量降低(图4),但是定位于叶绿体的PtAspAT同工酶酶活性却增高(图3)。可见PtAspAT同工酶酶活性大小与其对应的基因转录本的丰度高低变化趋势并不总是相同的。前人[25]在玉米N同化相关的酶编码基因研究中,也发现ZmAspAT的mRNA水平与酶活性变化趋势存在不一致的现象。实验结果与其相一致。这说明AspAT在植物体内的调控机制比较复杂,可能有转录或翻译后机制参与。
在N胁迫条件下,毛果杨不同组织(根、茎、老叶和新叶)中天冬氨酸转氨酶基因表达和酶活性实验结果表明,PtAspAT基因的mRNA表达量和酶活性大小与植物体氮浓度高低密切相关,为揭示AspAT在植物生长和胁迫响应中的代谢和生理意义奠定了理论基础。
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-21
中国动物保健(2022年9期)2022-09-25
中国农业科学(2022年13期)2022-07-26
微生物学杂志(2020年6期)2020-03-05
少儿科技(2019年9期)2019-09-10
江苏农业科学(2016年8期)2017-02-15
听力学及言语疾病杂志(2015年5期)2015-12-24
河北渔业(2015年9期)2015-09-15
中国当代医药(2015年21期)2015-03-01
中国酿造(2013年5期)2013-01-26
