
横断山云南山区不同地区大绒鼠种群的表型多样性
2020-12-21 07:24王政昆朱万龙
生物学杂志 2020年6期
任 月, 贾 婷, 章 迪, 张 浩, 王政昆, 朱万龙
(1. 云南省高校西南山地生态系统动植物生态适应进化及保护重点实验室 云南师范大学生命科学学院 生物能源持续开发利用教育部工程研究中心 云南省生物质能与环境生物技术重点实验室, 昆明 650500; 2.云南经济管理学院, 昆明 650106; 3.云南医药健康职业学院, 昆明 650106)
表型性状是探讨生物间系统发生和分类学的有效手段之一[1],由遗传和环境共同影响[2]。动物长期为适应不同地区的海拔、植被和气候,种间和种内表型性状可能出现分化,身体指标和头骨形态是哺乳动物重要的表型特征[3]。传统鉴定哺乳动物分类学方法亦应用身体指标和头骨形态特征为重要依据:Liu等[4]通过测量尾长、臼齿和阴茎形态,鉴定出四川一种新型田鼠,并通过形态学将指名亚属Eothenomys、东方绒鼠亚属Antheliomys和绒鼠平属Caryomys确立为绒鼠属Eothenomys的3个有效亚属[5]; Zeng等[6]通过对德钦绒鼠(Eothenomyswardi)的耳长、体长等34个表型性状研究,确立德钦绒鼠为一个有效物种;杨安峰等[7]对8种田鼠亚科和沙鼠亚科的阴茎形态进行比较,揭示属间和属内形态差异;王应祥[8]通过对树鼩表型性状的研究将中南半岛树鼩划分为6个亚种。表型性状特征可以推断不同环境下动物的进化特征和进化模式[9], Renaud等[10]揭示小林姬鼠(Apodemussylvaticus)下颌骨和臼齿表现出不同进化模式,并认为这与岛屿大小、远近和竞争水平有关;Yoram等[11]研究20世纪日本姬鼠(Apodemusspeciosus)和田鼠(Eothenomyssmithii)头骨,发现田鼠的眶间区颅骨最窄宽度和上齿列长有所增加,并将其归因于环境最低温度增加;Caumul等[3]对旱獭头骨进行研究,结果发现旱獭头骨形态的差异与其所处的环境因子之间具有密切关系。
横断山位于青藏高原(24°40′N~34°00′N,96°20′E~104°30′E),范围约36.4万km2,包括滇西、滇西北、川西、藏东南、青海东南部和甘肃西南部。横断山是世界25个多样性热点核心之一。横断山地区的地理隔离、构造隆升及气候变化造成该地区生物很可能存在生态物种表型分化情况[12]。大绒鼠(Eothenomysmiletus)属于绒鼠属中指名亚属,为横断山的固有种[13],本研究组之前研究发现不同地区的大绒鼠脏器和消化道出现分化[14],采用多维尺度分析发现5个地区大绒鼠头骨和臼齿均有一定的差异[15],本研究在这些研究基础上,采集德钦、香格里拉、丽江、剑川和哀牢山等5个地区大绒鼠,通过测定和比较身体指标和头骨指标的变化,探究大绒鼠对横断山不同地区的表型性状适应特征,阐明同一物种在不同环境中的表型性状适应对策。
1 材料与方法
1.1 动物来源
161只大绒鼠样本分别采自德钦(DQ)、香格里拉(XGLL)、丽江(LJ)、剑川(JC)和哀牢山(ALS)。实验动物均为非繁殖期的健康成年个体。各采样点的地理位置、气候特征以及样本数详见表1。
1.2 形态学测量
大绒鼠的身体指标测量参照高文荣[16]的方法。测量自然状态的体长、尾长等指标并记录(精确到0.01 cm)。大绒鼠的头骨指标测量参照杨奇森等[17]和夏霖等[18]的方法。测量颅全长、颅基长等指标并记录(精确到0.01 cm)。
1.3 统计方法
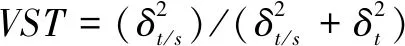

表1 本研究所有大绒鼠采样地点详细信息
2 结果与分析
2.1 种群间和种群内表型性状变异分析
单因素方差分析结果(表2)表明:丽江、剑川和哀牢山大绒鼠体长、尾长、胸宽、胸深、耳长和最大眶间距显著高于德钦和香格里拉种群;丽江种群躯干长和前肢长较大,香格里拉种群躯干长和前肢长较小;剑川种群后肢长显著大于其他种群;丽江、剑川和哀牢山种群的颅全长和颅高显著高于德钦和香格里拉种群,且剑川种群显著大于其他种群;剑川和哀牢山种群枕鼻长显著大于丽江和德钦种群;剑川种群颧宽显著大于哀牢山、德钦和香格里拉种群;哀牢山和香格里拉种群脑颅宽最大,德钦种群最小;剑川和哀牢山种群口盖长最大,德钦种群最小;剑川种群眼眶长最大,香格里拉种群最小;哀牢山种群听泡长显著大于其他种群;剑川和哀牢山种群上齿列长显著大于其他种群,且丽江种群显著大于德钦和香格里拉种群;哀牢山种群下齿列长最大,香格里拉种群最小;剑川种群最大,丽江种群最小;各种群间眼窝长无差异。
另外,表型性状种群内和种群间的方差分量及各性状的表型分化系数(表3)表明:24个表型性状在种群内的平均方差分量百分比为2.46%,种群间为26.62%,随机误差为 70.92%,说明种群间变异大于种群内变异; 种群间平均表型分化系数(VST)为0.875 4,说明种群间的贡献值为 91.09%,种群内的贡献值为 8.91%,种群间变异是表型变异的主要来源。
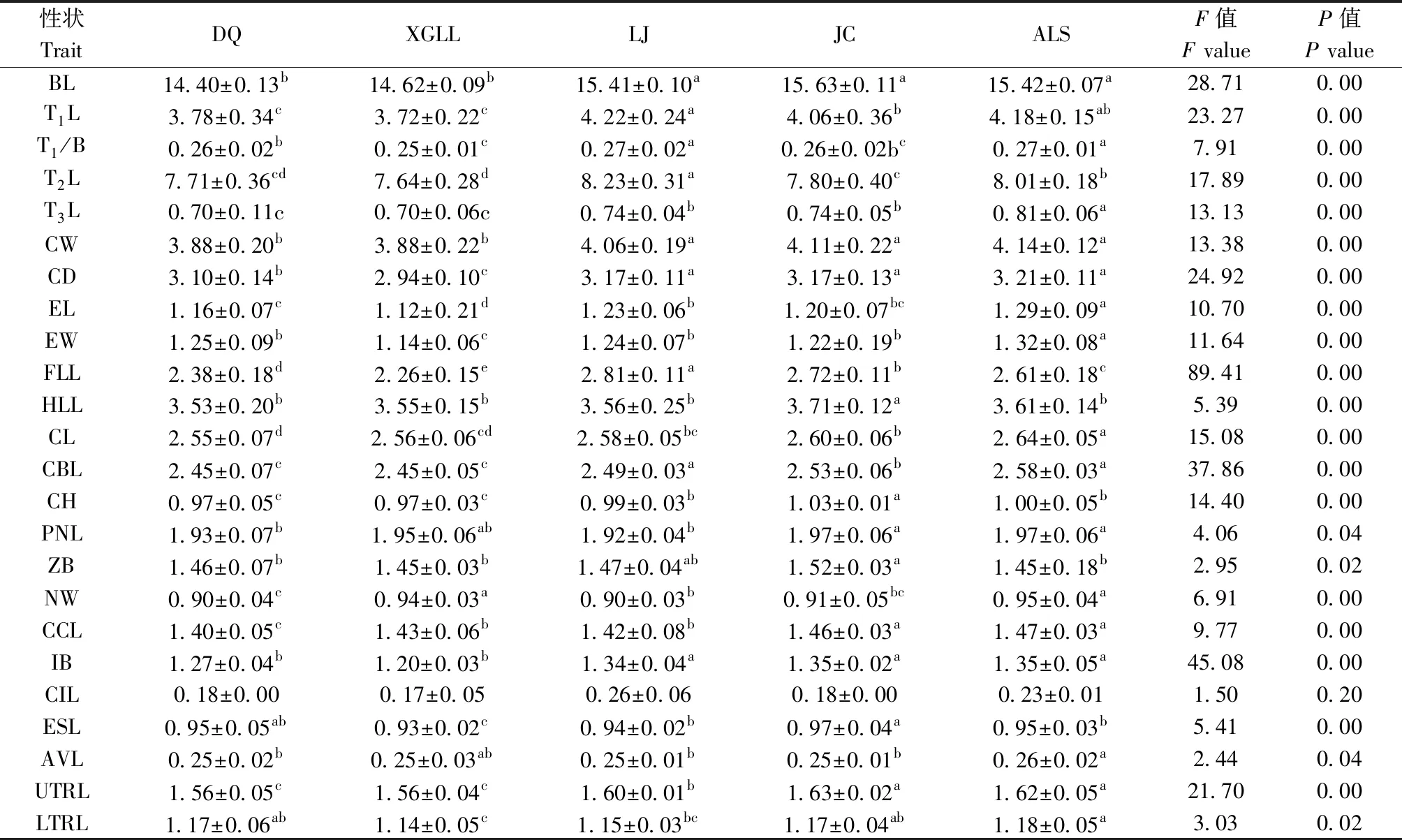
表2 大绒鼠身体指标和头骨指标比较
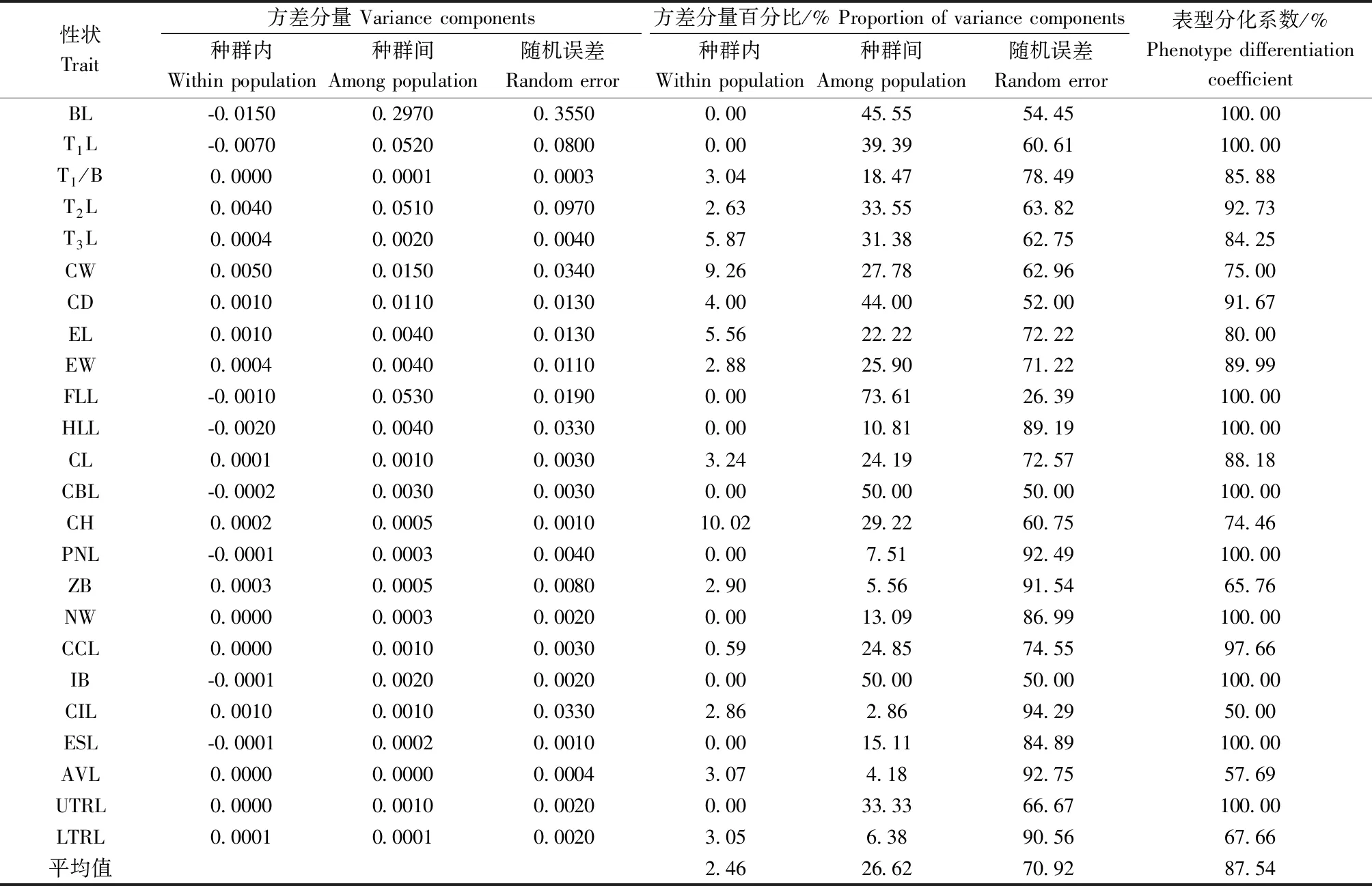
表3 大绒鼠表型性状的方差分量及其种群间表型分化系数
2.2 种群表型性状变异特征分析
24个表型性状在种群内不同个体的平均变异系数为4.8%,变异幅度为1.994%~8.660% (表4) 。吻长、耳长、眼窝长和听泡长表型性状的变异系数超过8.0%,远大于24个表型性状的平均变异系数; 除眼窝长和听泡长外,其余头骨指标远小于24个表型性状的平均变异系数4.8%,头骨性状稳定性大于身体指标稳定性。此外,尾长、尾体比、胸宽、耳宽前肢长和后肢平均变异系数均大于4.8%。
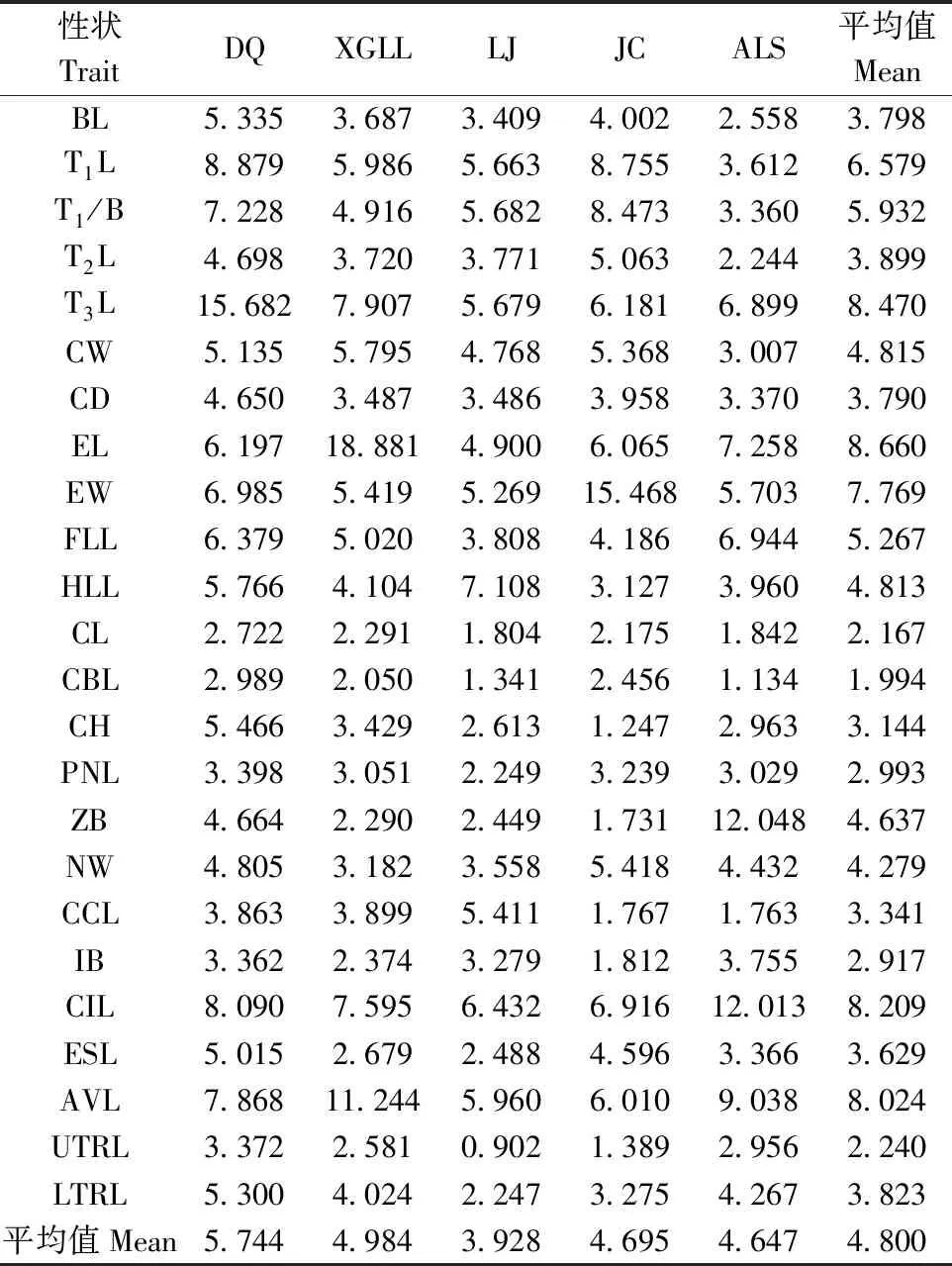
表4 大绒鼠种群表型性状的变异系数
2.3 身体指标主成分分析和聚类分析
利用主成分对大绒鼠外形和头骨数据进行主成分分析(表5和图1)发现:第一、第二和第三主成分的特征值依次为6.655、2.809和1.715,占全部变量的46.58%;以第一和第二主成分作散点图,德钦和香格里拉种群多分布于图片的右下方,丽江、剑川和哀牢山多分布于图片左上方;主成分分析图发现2大支并没有明确分开,来自不同地区的样本互相都有一些重叠。对大绒鼠身体指标和头骨数据进行聚类分析(图2)发现,5个种群的大绒鼠聚为2大支:德钦和香格里拉一支,丽江、剑川和哀牢山一支。
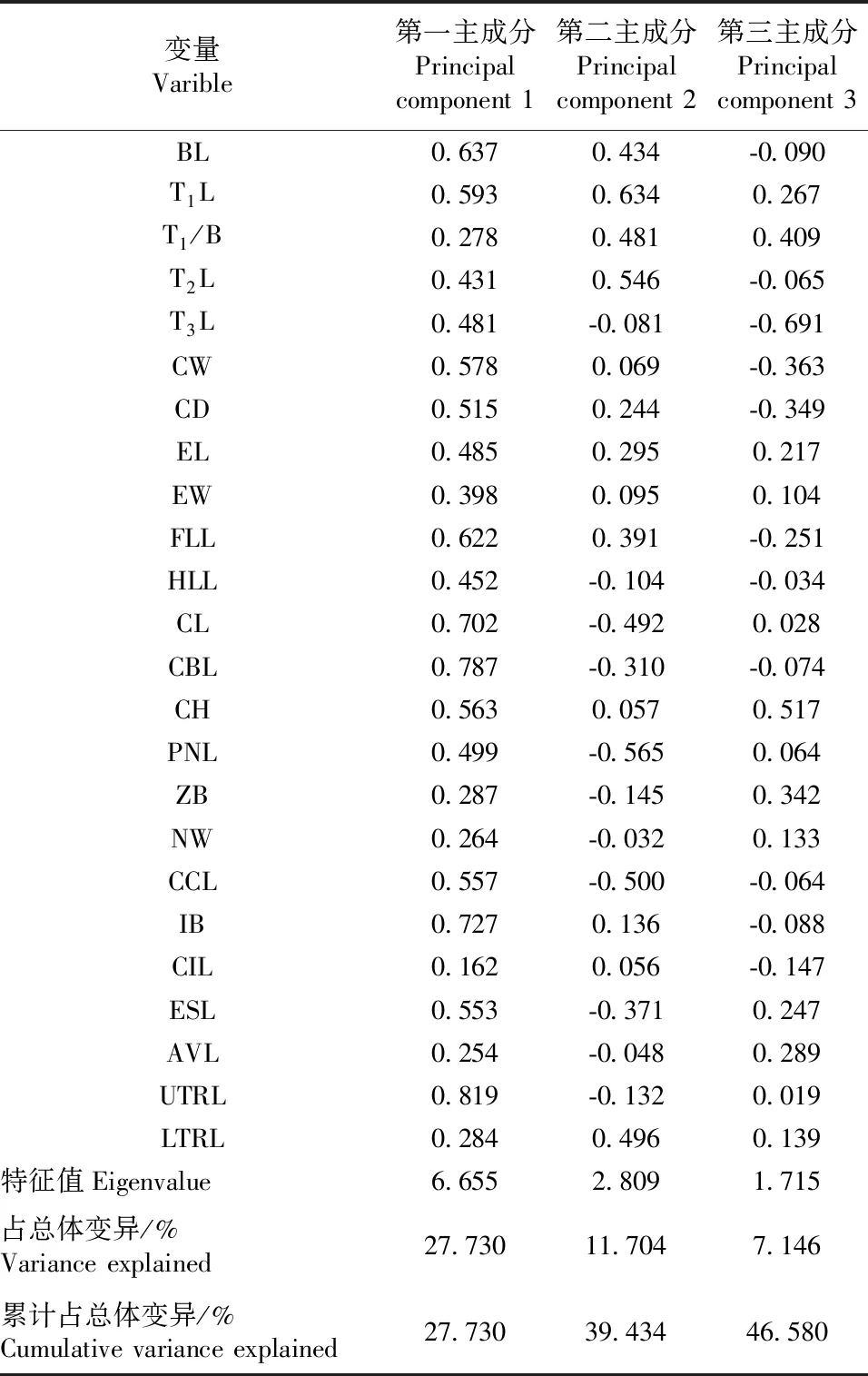
表5 大绒鼠形态学数据主成分分析前3个主成分的特征值、占总体变异量及累计占总体变异
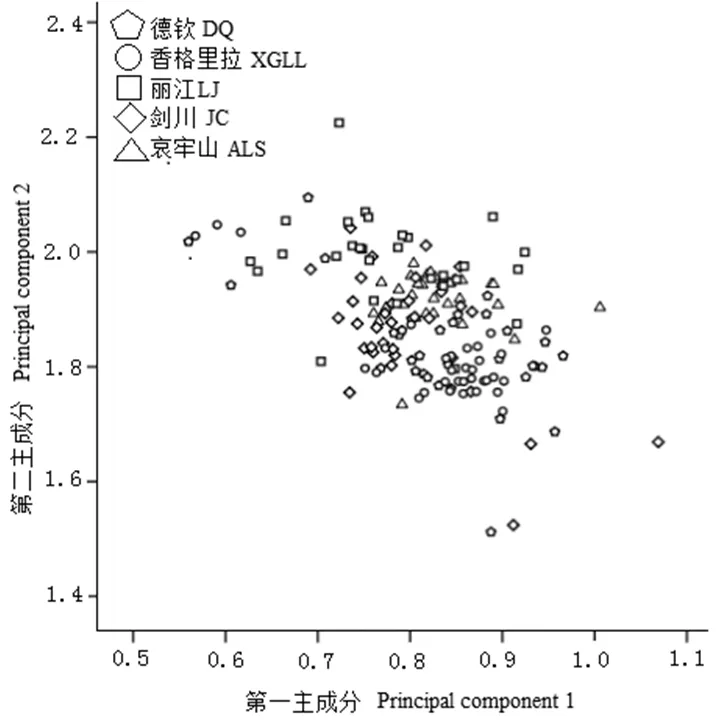
图1 大绒鼠主成分分析图
3 讨论与结论
身体指标和头骨形态学对研究种间和种内关系具有重要参考价值[20]。为适应不同海拔、植被、纬度和气候的环境,动物体型和头骨会分化。关于环境温度对动物体型和头骨形态影响有两种解释。一种是Bergmann定律,Bergmann指出环境温度高的地区动物体型较小,体表面积大,有利于动物散热;而寒冷环境中,动物体型较大,体表面积较小,有效于保存热量[21-23]。如:全球变暖导致以色列鸟类体重下降[24];迁徙滨鸟体重与环境温度呈正相关,与纬度成负相关[25]。另一种解释是环境温度升高会促进植被生长,增加动物食物供应,导致动物体型增加。研究发现过去两个世纪全球变暖,欧洲人的身高和体重显著增加[26],在动物中也发现类似的现象,如由于全球变暖,海港海豚(Phocoenaphocoena)食物增加,体型增加,雌性成熟期提前[27],丹麦的红狐(Vulpesvulpes) 和亚洲豹(Melesmeles) 头骨大小增加[28],以色列几种食肉动物体长也有增加[29]。中缅树鼩的颅全长、基长、颧宽、齿长、吻长、最大颅宽和最大腭宽均显著小于马来树鼩[30]。
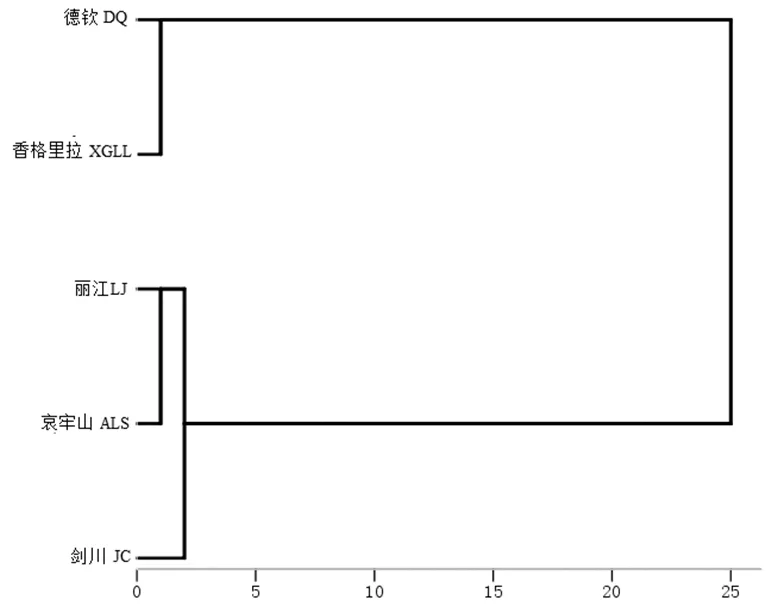
图2 基于大绒鼠表型形态学数据的聚类分析关系图
本研究采样点沿横断山云南山区自北向南依次是德钦、香格里拉、丽江、剑川和哀牢山,其中德钦和香格里拉海拔在3 000 m以上,年平均温度在5 ℃左右,主要植被为高山草甸;丽江、剑川和哀牢山海拔在2 000~3 000 m,年平均温度大于10 ℃,主要植被为灌木和草丛。相对低海拔、高温地区,大绒鼠体长、尾长、躯干长、胸宽、耳长、前肢长、最大眶间宽、上齿列长均大于高海拔、低温地区大绒鼠,与Bergmann定律相悖,推测由于低海拔、高温地区促进植物生长,延长植物生长周期,植物丰富度高于低海拔、高温地区,增加大绒鼠的食物供应。本研究组之前研究结果表明:在冬天和冷驯化条件下大绒鼠体重、体脂下降,静止代谢率、摄入能、非颤抖性产热、解偶联蛋白1表达增加,最终适应低温环境并在脱冷条件下恢复到对照组水平[31-34];对头骨进行多维尺度分析亦发现5个地区大绒鼠头骨存在差异,并推测这差异可能是受经度变化影响的[15],本研究结果与之前研究结果相符。Yoram等[11]对20世纪的田鼠头骨形态研究发现,随全球变暖,田鼠眶间颅骨最窄宽度和上齿列长有所增加,这与本研究发现相对低海拔、高温地区大绒鼠的最大眶间距和上齿列长大于高海拔、低温地区大绒鼠相符。
对5个不同地区大绒鼠的身体指标和头骨形态的主成分分析和聚类分析发现,相对高海拔、低温地区聚成一支,而低海拔、高温地区的大绒鼠聚成一支,推测不同地区的大绒鼠在环境海拔和温度影响下出现了表型分化。
综上所述,横断山云南山区5个不同地区大绒鼠在身体指标和头骨形态上出现表型分化,且这种表型分化可能是由海拔、温度、食物共同影响的。
猜你喜欢
环球时报(2022-11-10)2022-11-10
金沙江文艺(2021年10期)2021-10-08
云南画报(2021年5期)2021-07-22
奥秘(2020年7期)2020-08-02
滇池(2019年12期)2019-11-18
民族音乐(2019年3期)2019-08-14
现代园艺(2018年2期)2018-03-15
云南画报(2017年4期)2017-06-23
滇池(2015年12期)2015-12-11
西南学林(2013年2期)2013-11-12
