
二肽对猪卵母细胞体外成熟、受精及胚胎发育的影响
2020-11-25 01:17乔德瑞张晓隽
中国饲料 2020年20期
乔德瑞,张晓隽
(1.晋中职业技术学院,山西晋中 030600;2.河南六和饲料有限公司,河南焦作 454350)
猪卵母细胞的体外成熟和体外受精已被广泛研究。卵母细胞成熟是体外生产系统中的关键步骤,它可以影响细胞质量,进而影响胎儿发育和后代健康。但体外成熟的卵母细胞与体内成熟的卵母细胞相比,发育能力下降,表明体外成熟的卵母细胞仍处于次优状态。因此,对培养基进行一定的改善有助于提高卵母细胞体外成熟效率。氨基酸在大多数卵细胞成熟和胚胎培养的培养基中均有添加,因为它们具有多种生理功能,包括蛋白质和核苷酸的合成能源供应,抗渗透休克、氧化应激,pH 调节(Lane,2011)。氨基酸,尤其是谷氨酰胺的自发降解和分解代谢可导致氨的产生,这对活细胞在体内和体外都有毒(沈伟涛和魏婷,2018)。培养基中氨的存在导致细胞内pH 降低,减少氧化磷酸化和囊胚细胞数,最终改变胎儿发育和增长率(Zander 等,2006)。通过在培养基中添加二肽直接供哺乳动物细胞体外使用可避免谷氨酰胺的毒性作用,有研究表明,谷氨酰胺可以被L- 丙氨酰-L- 谷氨酰胺和L- 甘氨酰-L-谷氨酰胺取代,以达到小鼠胚胎发育水平的提高(Biggers,2004)。二肽在卵母细胞成熟和猪精子顶体反应中起重要作用(Tareq 等,2008)。虽然上述研究报道了二肽在胚胎发育中的重要性,但关于二肽(如甘氨酰谷氨酰胺或丙氨酰谷氨酰胺)对离体猪胚胎发育的直接影响数据有限。因此,本研究旨在探讨单独的二肽或联合使用对猪卵母细胞离体成熟、受精和发育及对培养基中氨积累的影响。
1 材料与方法
1.1 卵母细胞的恢复和体外成熟 从当地屠宰场收集母猪卵巢,收集后1 ~3 h 于30 ~35℃条件下补充100 IU/mL 青霉素和100 mg/mL 硫酸链霉素的生理盐水保存。参考刘晓鹏和王华岩(2016)的研究方法分离、收集及培养卵母细胞。在猪胚胎细胞培养液中用不同浓度的氨基酸及二肽组合处理胚胎,对照组为基础培养液,T1 ~T6组分别做如下处理:T1 组添加1 mmol 谷氨酰胺、T2 组加入1 mmol 谷氨酸、T3 组加入2 mmol 丙氨酰谷氨酰胺、T4 组加入2 mmol 甘氨酰谷氨酰胺、T5 组2 mmol 丙氨酰谷氨酰胺+2 mmol 甘氨酰谷氨酰胺、T6 组1 mmol 谷氨酰胺+1 mmol 谷氨酸。
1.2 精子准备、体外受精及胚胎细胞数评估 参考Tareq 等(2007)的研究方法进行体外受精。根据Hunter 和Polge(1996)所述的方法评估卵母细胞的减数分裂阶段。
1.3 放射性标记葡萄糖的掺入、氧化及氨氮浓度参考Tareq 等(2005)的方法测定培养基中氨氮浓度。放射性底物14C- 葡萄糖的标记、掺入及氧化测定参考Tsujii 等(2009)的研究方法。
1.4 统计分析 卵母细胞随机分布于各组,每组5 个重复,试验数据采用SAS 软件单因素方差统计模型,采用Duncan’s 法进行多重比较,P <0.05为差异有统计学意义。
2 结果与分析
2.1 二肽对猪卵母细胞成熟和受精率的影响由表1 可知,卵母细胞分裂中期成熟率及单精受精卵和精原细胞受精率在丙氨酰谷氨酰胺组、甘氨酰谷氨酰胺组及丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺组表现最高(P <0.05)。与丙氨酰谷氨酰胺或甘氨酰谷氨酰胺单独使用相比,联合使用组显著提高了卵母细胞分裂中期成熟率、单精受精卵和精原细胞受精率(P <0.05)。
2.2 二肽对猪卵母细胞成熟的影响 由表2 可知,联合使用丙氨酰谷氨酰胺和甘氨酰谷氨酰胺显著提高了卵母细胞成熟和14C- 葡萄糖的掺入和氧化(P <0.05),而对照组与谷氨酰胺、谷氨酸及联合使用谷氨酰胺和谷氨酸对卵母细胞成熟指标的影响无显著差异(P >0.05)。此外丙氨酰谷氨酰胺和甘氨酰谷氨酰胺联合使用组卵母细胞成熟与受精过程中培养液氨氮浓度表现最低(P<0.05)。
2.3 二肽对卵母细胞胚胎发育及胚胎14C- 葡萄糖掺入、氧化及氨氮浓度的影响 由表3 可知,丙氨酰谷氨酰胺、甘氨酰谷氨酰胺及丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺组胚胎发育2 ~4 个、8 ~16 个及桑椹胚和囊胚数显著高于其他组(P<0.05),同时丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺组胚胎发育2 ~4 个、8 ~16 个及桑椹胚和囊胚数显著高于丙氨酰谷氨酰胺和甘氨酰谷氨酰胺组(P <0.05)。
由表4 可知,丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺组2 细胞和囊胚14C- 葡萄糖掺入、氧化量显著高于对照组,而氨氮浓度显著低于对照组(P <0.05)。
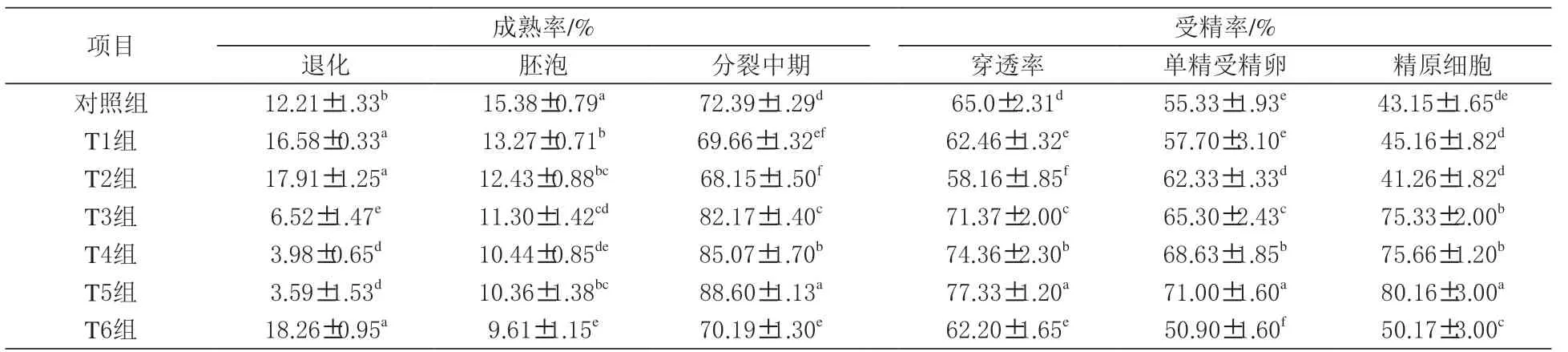
表1 二肽对猪卵母细胞成熟和受精率的影响
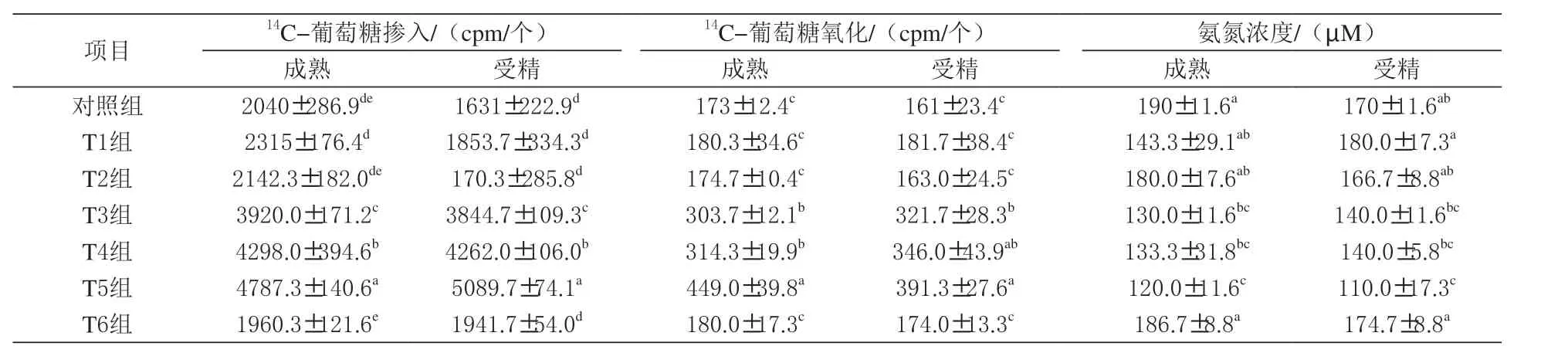
表2 二肽对猪卵母细胞成熟和受精过程14C- 葡萄糖掺入、氧化及氨氮浓度的影响
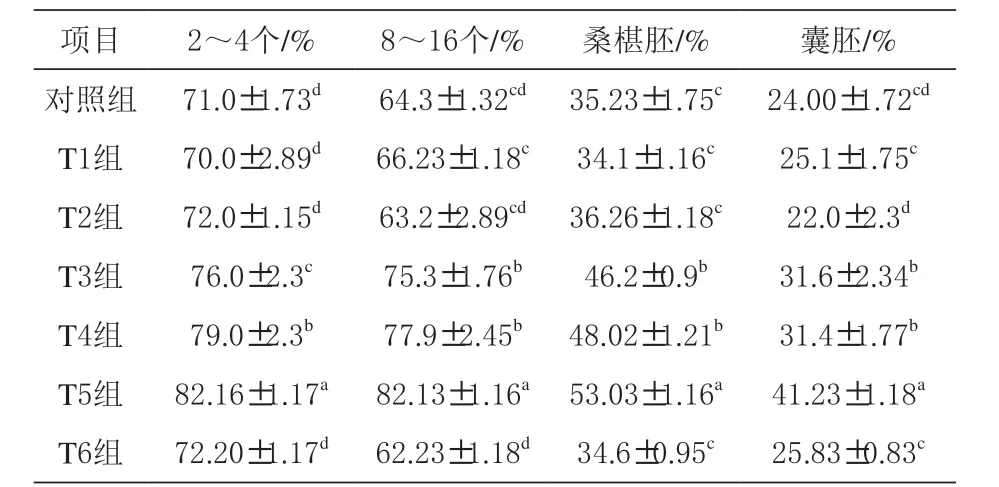
表3 二肽对猪卵母细胞胚胎发育的影响
2.4 二肽对囊胚细胞数量的影响 由表5 可知,丙氨酰谷氨酰胺、甘氨酰谷氨酰胺及丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺组内细胞团数和滋养外胚层细胞数显著高于对照组(P <0.05),同时丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺组内细胞团数和滋养外胚层细胞数显著高于其他各组(P <0.05)。
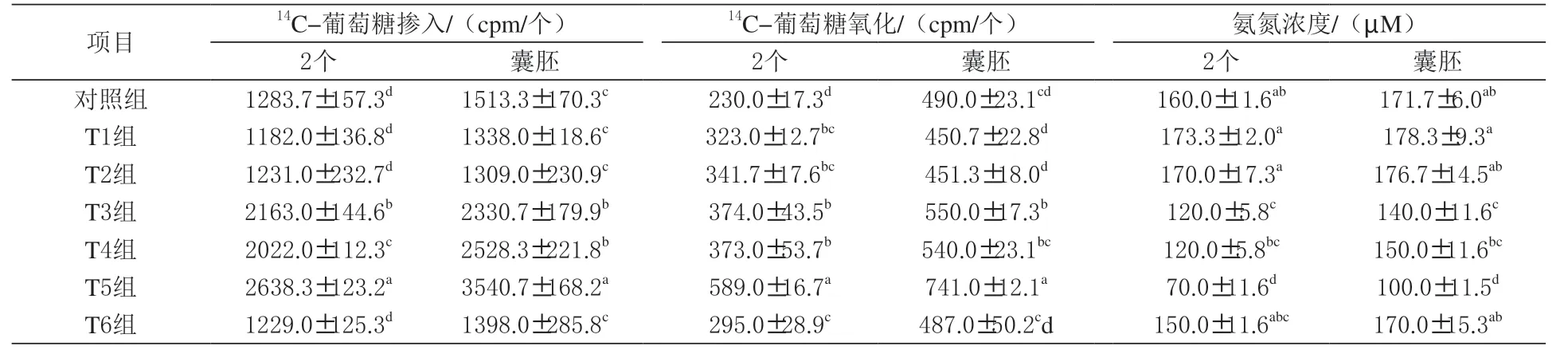
表4 二肽对猪卵母细胞胚胎14C-葡萄糖掺入、氧化及氨氮浓度的影响
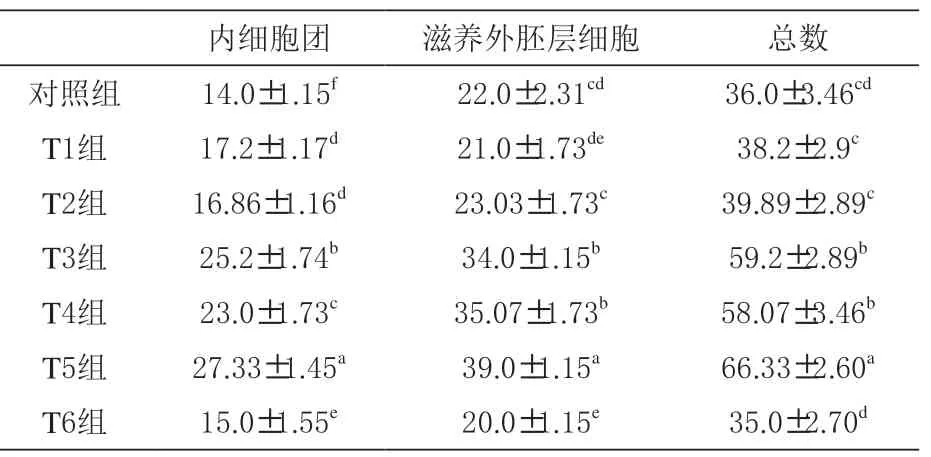
表5 二肽对囊胚细胞数量的影响
3 讨论
本试验发现,联合使用丙氨酰谷氨酰胺和甘氨酰谷氨酰胺使卵母细胞成熟、授精和胚泡阶段氨氮浓度较对照组分别降低了120、70 和100 μmol,其中本研究中氨基酸和二肽的最佳浓度与猪卵母细胞和精子的最佳浓度相似(Tareq 等,2008)。这些结果表明,对氨的敏感性取决于卵母细胞或胚胎的发育阶段,可能与卵丘细胞的存在有关。Yuan 和Krisher(2010)提出,氨对卵母细胞分离中期有不利影响,而氨是卵母细胞在体外成熟过程中产生的。卵母细胞在体外培养过程中卵丘细胞起至关重要的作用,因为卵丘细胞的体外生长和代谢受类似于卵泡液中氨浓度的改变,这些卵丘细胞支持卵母细胞体外成熟的能力受到损害。另外,卵丘细胞可以减少卵母细胞直接接触到氨的量,阻止培养基中的氨通过缝隙连接转运到卵母细胞(Tareq 等,2007)。本数据表明,丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺的添加对培养基中卵母细胞分离中期、单精受精卵和精原细胞形成、成熟和受精的影响最大。
体外研究证明,氨基酸在体内支持正常的着床前发育。氨基酸的自然降解和分解会形成吡咯烷酮碳酸和潜在的有害氨。氨对活细胞的毒性在体内外都是公认的。具体来说,氨含量的增加降低了pH,增加了渗透压,导致精子活力逐渐丧失,并降低了猪卵母细胞分裂中期和单精受精的速率(Tareq 等,2007)。本试验结果表明,与其他处理组相比,丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺处理减少了氨的积累,对卵母细胞成熟、受精和发育成囊胚方面有重要作用。
一些研究已经证明,哺乳动物葡萄糖代谢对植入前胚胎的影响,对猪胚胎的营养摄取研究表明,胚胎在发育的所有阶段都消耗葡萄糖并产生乳酸。因此,含葡萄糖的培养基常被用于体外培养猪胚胎。葡萄糖的结合和氧化是卵母细胞活力和产生卵子的指标(Tsujii 等,2009)。本研究丙氨酰谷氨酰胺+ 甘氨酰谷氨酰胺处理的卵母细胞在成熟、受精、细胞和囊胚阶段的14C 标记葡萄糖的掺入率和氧化率明显高于对照组,这与Tareq等(2012)的研究结果一致。因此,在胚胎发育阶段,当胚胎对能量的需求较高时,培养基中的高氨氮可能会降低胚胎细胞对ATP 的可利用性,导致退化卵增多,囊胚发育阶段减少。但丙氨酰谷氨酰胺+甘氨酰谷氨酰胺产生的内细胞团和滋养外层细胞数量的显著增加,表明丙氨酸和甘氨酸是通过一种未知的机制起主要作用(Tareq 等,2012)。
4 结论
本研究结果表明,二肽参与调控猪卵母细胞的成熟、受精和胚胎发育,其中将丙氨酰谷氨酰胺和甘氨酰谷氨酰胺结合可以降低氨积累,提高猪胚胎细胞成熟、受精、胚胎发育和葡萄糖的摄入速度。
猜你喜欢
化工管理(2022年14期)2022-12-02
生物技术进展(2022年5期)2022-10-11
中国畜牧杂志(2022年4期)2022-04-15
中国畜牧杂志(2021年5期)2021-12-05
天津医科大学学报(2021年2期)2021-03-29
食品与生物技术学报(2021年4期)2021-01-17
中国金属通报(2020年8期)2020-12-31
岭南现代临床外科(2020年5期)2020-12-13
中国畜牧杂志(2020年6期)2020-07-11
哈尔滨工业大学学报(2020年6期)2020-06-11
