
舟山近海入侵种
——条纹锯鮨的形态特征与DNA 条形码研究
2020-09-28 00:59王业辉高天翔李伟业
浙江海洋大学学报(自然科学版) 2020年1期
王业辉,高天翔,李伟业
(1.中国海洋大学水产学院,山东青岛 266003;2.浙江海洋大学水产学院,浙江舟山 316022;3.舟山市水产研究所,浙江舟山 316000)
条纹锯鮨Centropristis striata (Linnaeus 1758),又名美洲黑石斑,英文名BlackSeabass,属硬骨鱼纲Osteichthys、辐鳍亚纲Actinopterygii、鲈形目Perciformes、鮨科Serrnanidae、锯鮨属Centropristis,是海洋暖温性鱼类,分布于美国东海岸即大西洋沿岸25°-45° N,从加拿大东南沿海至美国的缅因州到佛罗里达州东北部和墨西哥湾东部近海都有分布,分布纬度与我国东海南部到日本海相当。条纹锯鮨具有生长快、抗逆性强,适应温度、盐度较广等特点,能够完全适应我国近海海域环境,适于在我国温带和亚热带海域进行养殖[1-3]。
条纹锯鮨因其在商业和休闲渔业方面的巨大价值,被认为是美国东南沿海最具商业开发价值的鱼类之一[1,4]。条纹锯鮨最先于2002 年由国家海洋局第一海洋研究所和荣成礼村渔业总公司分别从美国引进鱼苗和受精卵进行养殖和育苗试验,2003 年,青岛三高大菱鲆研究所从美国东海岸引入条纹锯鮨进行人工培育,并于2006 年生产商品苗种100 余万尾,为大规模养殖奠定了基础[2-3]。舟山市水产研究所于2016年从青岛引进鱼苗并开始繁育和养殖推广。国外研究主要集中在生态因子[5-6]、生活习性[7]等方面。在国内,对条纹锯鮨的研究主要有生理生化方面[8-9]以及饲料[10-11]、卵巢发育及性逆转[12]、以及病菌[13-14]等与养殖相关的方面。
生物入侵(biological invasion)是指生物由原栖息地经自然或人为的途径侵入到另外一个环境,对入侵地的生物多样性以及人类的生产和健康造成损失或生态灾难的过程[15]。鱼类入侵(fish invasion)指外来鱼种由原生地通过某种途径到达其历史上没有分布的新生态环境,在新生境繁殖、扩散并对入侵地生态系统、生物多样性或人类生活造成生态损害或经济损失的过程[16]。生物入侵已成为国内外研究的热点,鱼类入侵作为生物入侵的重要组成部分,亦需要得到人们的关注,但关于鱼类入侵方面的研究却较少[17-18]。
DNA 条形码(DNA barcoding)技术是通过对一个标准目的基因的DNA 序列进行分析从而进行物种鉴定的技术,能够快速有效的鉴定物种。HEBERT,et al[19]提出了采用线粒体DNA 的COI(细胞色素c 氧化酶亚基I)基因片段作为物种鉴定的标准片段——即DNA 条形码,对11 个门13 320 个物种的COI 基因序列进行分析,结果表明COI 基因能够很好地区分鱼类物种[19]。目前,DNA 条形码技术已经广泛应用于物种鉴定,新种、新纪录种的发现等方面[20-24]。
本研究对舟山西轩岛附近海域捕获的一尾条纹锯鮨和养殖样品进行了形态特征比较分析和DNA 条形码研究,并探讨了其入侵的可能性与风险,提出其入侵的危害和防止措施。
1 材料与方法
1.1 样品采集与处理
海捕条纹锯鮨样品(编号为Y1)于2018 年12 月在舟山市西轩岛附近海域(29°89′ N,122°31′ E)捕获。样品采集方法为底拖网。海捕野生条纹锯鮨样品于采集现场冷冻保存,并运输到实验室进行二次处理、拍照与标本制作。
用于比较研究的条纹锯鮨采自舟山市水产研究所养殖场,共计10 尾(5 尾成鱼,5 尾幼鱼,编号为T1-T10)。与海捕野生条纹锯鮨样品采用相同方法处理。
1.2 形态学测量
对野生与养殖的11 尾条纹锯鮨样品用电子天平进行称重,精确至0.01 g。采用传统形态学方法对11尾样品的7 个可数性状(背鳍鳍棘数、背鳍鳍条数、胸鳍鳍条数、腹鳍鳍棘数、臀鳍鳍棘数、臀鳍鳍条数、尾鳍鳍条数)进行直接计数。可量性状包括传统可量性状与框架法数据。传统可量性状包括体长(BL)、体高(BD)、体宽(BW)、头长(HL)、眼后头长(POL)、吻长(SL)、眼间隔(EI)、眼径(ED)、上颌长(UJL)、下颌长(LJL)、尾柄长(CPL)、尾柄高(CPD)共计12 种;框架法数据包括吻至背鳍起点(X1-2)、吻至腹鳍起点(X1-3)、背鳍起点至腹鳍起点(X2-3)、背鳍基长(X2-4)、背鳍起点至臀鳍起点(X2-5)、背鳍起点至臀鳍末端(X2-7)、背鳍末端至臀鳍起点(X4-5)、背鳍末端至臀鳍末端(X4-7)、背鳍末端至尾鳍腹部起点(X4-8)、尾鳍背部起点至臀鳍末端(X6-7)、尾鳍背部起点至尾鳍腹部起点(X6-8)、腹鳍起点至臀鳍起点(X3-5)、臀鳍基长(X5-7),臀鳍末端至尾鳍腹部起点(X7-8)共计14 种。所有长度均采用游标卡尺进行测量,精确至0.02 mm。
1.3 DNA 提取
采用苯酚-氯仿法提取基因组DNA,乙醇沉淀后的基因组DNA 溶于100 mL 灭菌水中,4 ℃保存。
1.4 PCR 扩增与测序
扩增12S rRNA 短片段采用正向引物MiFish-U-F 与反向引物MiFish-U-R[25],引物序列为:MiFish-UF:5’-TTTGTCGGTAAAACTCGTGCCAGC-3’,MiFish-U-R:5’-CAAACTGGGATTAGATACCCCACTATAA-3’。扩增CO I 片段采用正向引物F1 与反向引物R1[20],引物序列为:F1:5’-TCAACCAACCACAAAGACATTGGCAC-3’,R1:5’-TAGACTTCTGGGTGGCCAAAGAATCA-3’。
PCR 反应总体系为25 μL,包括17.5 μL 灭菌水,2.5 μL 的10×buffer,2 μL 的dNTPS (25 mM),正反向引物各1 μL,0.15 μL 的EasyTaq 酶(上海派森诺生物有限公司),1 μL 的DNA 模板。反应条件如下:94 ℃预变性5 min,然后94 ℃变性45 s,57 ℃退火45 s,72 ℃延伸45 s,35 个循环,最后72 ℃延伸10 min,4 ℃永久保存。PCR 仪为Takara PCR Thermal Cycler MP (TP3000)。PCR 产物采用1%琼脂糖凝胶电泳检测,条带明亮且单一的PCR 产物送至上海派森诺生物有限公司进行双向测序。
1.5 数据处理
对于可数性状,计算其平均数。对于可量性状,为消除不同个体大小对各参数值的影响,将头部的量度特征值除以头长,体宽除以体高,尾柄长除以尾柄高,其余各特征值均除以体长,得到25 个变量(X1-X25)进行数据分析。比较野生条纹锯鮨与养殖条纹锯鮨的形态特征差异。
将测得的12S rRNA 与CO I 基因片段序列用DNAStar 软件包中的SeqMan 软件进行比较,并辅以人工校正。从NCBI (national center for biotechnology information)上下载C.striata 的同源片段序列进行比较分析。使用MEGA6.0 软件计算11 尾条纹锯鮨的碱基组成与遗传距离,并基于K2P (kimura 2-parameter)模型构建邻接(neighbor-joining)系统发育树[26]。
2 结果
2.1 形态学特征
2.1.1 外部特征
舟山市水产研究所养殖条纹锯鮨(幼鱼、成鱼)与海捕野生条纹锯鮨外形照片见图1。条纹锯鮨体延长,稍侧扁。口中大,略倾斜,下颌稍突出。前鳃盖骨边缘具锯齿,鳃盖骨具棘。背鳍连续具浅缺刻,从鳍基部至鳍末端均有多排白色色带。侧线一条。体表呈烟灰色、褐色或蓝黑色,皮肤上有显著的菱形白色斑点[1,3]。
2.1.2 可数可量性状
可数、可量性状见表1-2。背鳍D:X—11~12;胸鳍P:18~19;腹鳍V:VI;臀鳍A:III—7~8;尾鳍C:17~18。
头长比体长为0.353-0.402,吻长比头长为0.235-0.301,眼径比头长为0.183-0.238,眼间隔比头长为0.323-0.473,眼后头长比头长为0.464-0.561,上颌长比头长为0.303-0.442,下颌长比头长为0.191-0.376,体高比体长为0.302-0.418,尾柄长比体长为0.176-0.222,尾柄高比体长为0.118-0.152,尾柄长比尾柄高为1.171-1.710。

图1 舟山市水产研究所养殖条纹锯鮨(幼鱼、成鱼)与海捕条纹锯鮨Fig.1 C.striata(juveniles,adult fish) of Zhoushan fisheries research institute and caught individual

表1 养殖样品与近海捕获样品的可数性状(括号内为平均值)Tab.1 Countable characters of aquaculture samples and caught samples(numbers in the brackets are mean values)
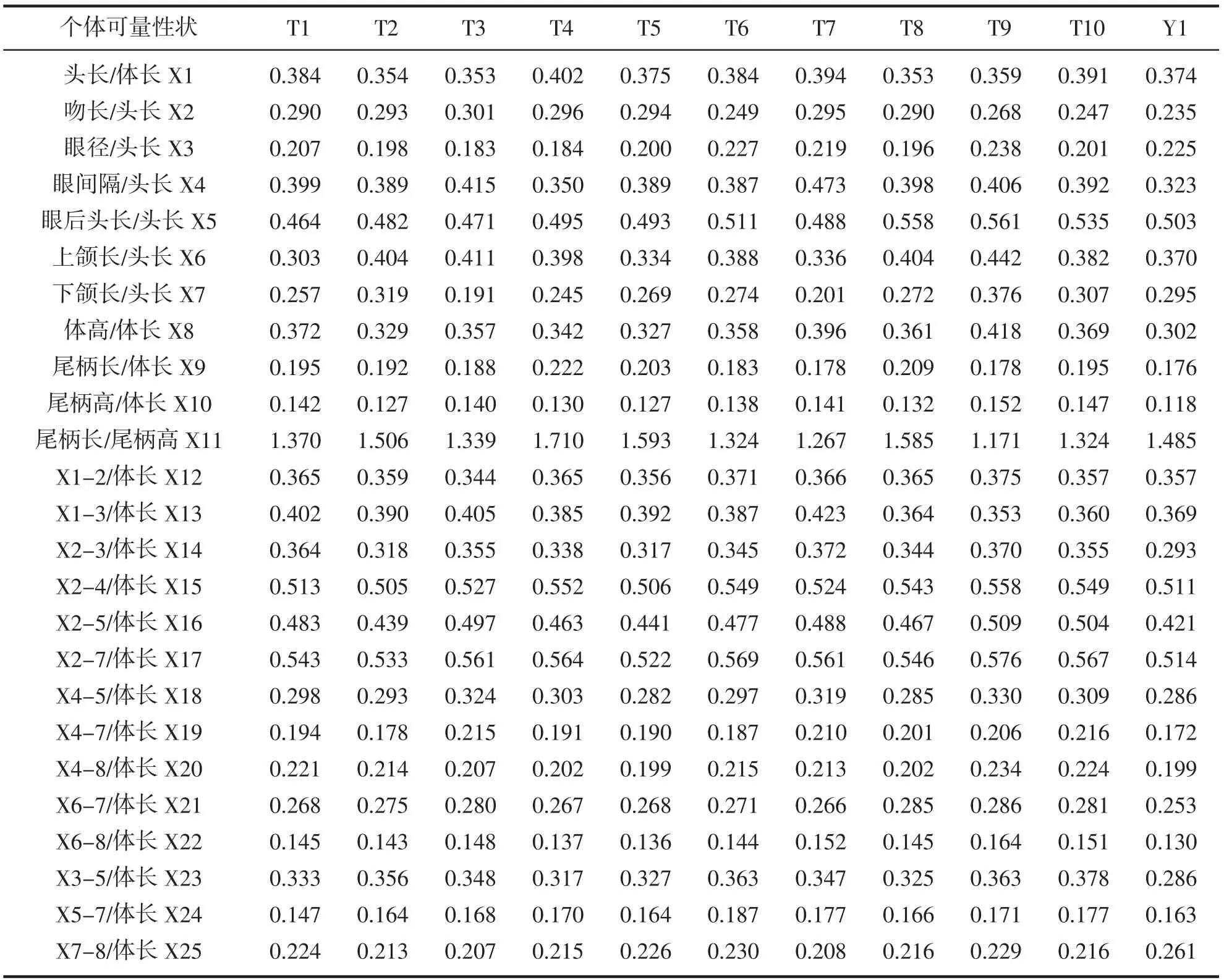
表2 养殖样品与近海捕获样品的可量性状Tab.2 Measurable characters of aquaculture samples and caught samples
2.2 DNA 条形码分析
2.2.1 序列分析
对海捕样品与10 尾养殖条纹锯鮨的CO I 与12S rRNA 基因进行双向测序后,CO I 基因得到652 bp的序列,12S rRNA 得到169 bp 的序列。
海捕条纹锯鮨样品的CO I 基因碱基组成为A:22.5%,G:21.0%,C:29.3%,T:27.1%,G+C:50.3%。10尾养殖个体的CO I 基因碱基组成相同,均为A:22.2%,G:21.0%,C:29.3%,T:27.5%,G+C:50.3%。其中G含量低于其他碱基含量,符合脊椎动物线粒体DNA 反G 偏倚的特点[27]。海捕样品与养殖个体相比,在CO I 基因片段第16 与第28 位点处发生转换,均为T→A。
海捕样品与10 尾养殖条纹锯鮨的12S rRNA 基因片段碱基组成完全相同,均为A:33.1%,G:23.1%,C:23.7%,T:20.1%,G+C:46.8%。
2.2.2 同源序列比较
从GenBank 中下载C.striata 的5 条同源片段序列(序列号分别为HQ024933,JN021294,JQ841849,KC015262,KX688305)进行比较分析,基于CO I 基因与12S rRNA 基因分别构建邻接系统发育树,均加入横纹九棘鲈Cephalopholis boenak 作为外群。从所构建的2 个系统发育树(图2,图3)中可以看出,所有条纹锯鮨序列均聚为一枝,并与外群明显分开。
基于CO I 基因条纹锯鮨的遗传距离最大为0.005,基于12S rRNA 基因条纹锯鮨的遗传距离为0,可判定为条纹锯鮨。
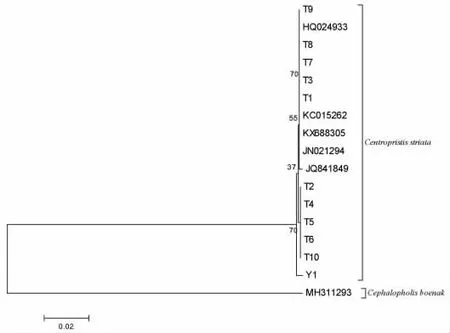
图2 基于CO I 基因构建邻接系统发育树Fig.2 Neighbor-joining phylogenetic tree based on CO I gene

图3 基于12S rRNA 构建邻接系统发育树Fig.3 Neighbor-joining phylogenetic tree based on 12S rRNA gene
3 讨论
3.1 条纹锯鮨成为入侵种的可能性分析
通过与舟山市水产研究所养殖条纹锯鮨的形态学与DNA 条形码比较分析,可以确定这尾海捕样品为条纹锯鮨。
入侵鱼种通常具有个体小、食性广、适用能力强、繁殖量大等特征[28-30]。条纹锯鮨属于底栖杂食性鱼类,其食性以动物性饵料为主,对食物适应性强,被认为是随机取食者和不挑食者,其食物的多样性与其特殊的栖息地生态环境有关[31,32]。条纹锯鮨对环境的适应能力也较强,其生存水温为5~30℃,适盐范围为5~36,需氧量低,能够适应较差环境[2]。其繁殖能力强,产卵量大,成鱼可产卵100 万粒以上[1]。条纹锯鮨因为其自身对环境的强适应性以及强繁殖能力,存在较强的生态入侵能力。
生物入侵地的自然环境特征决定了外来物种能否生长和繁衍[33]。条纹锯鮨在美国东海岸,分布纬度与我国东海南部到日本海相近,环境特征基本相同,适合我国近海生存,具备成为生态入侵种的条件。
本地鱼类区系的结构稳定性是对抗外来鱼类入侵的重要因素,具有高度生物多样性和稳定结构的水体不易被外来鱼类入侵,具有较低生物多样性和结构不稳定的水体则易被外来物种入侵[34]。由于我国近海捕捞压力较大,对近海环境和渔业资源破坏严重,鱼类生物多样性和结构不稳定,也为条纹锯鮨的入侵提供了契机。
3.2 入侵途径
几十年来,我国从国外引进了许多养殖的新品种,有的已经成为我国重要的养殖经济品种,如大菱鲆Scophthalmus maximus、凡纳滨对虾Litopenaeus vannamei 等,引进外来鱼种丰富了我国水产品的种类,创造了巨大的经济价值[18],但是引进的养殖品种同样也是外来生物入侵的重要途径之一,在养殖过程中,存在鱼类从养殖网箱中逃逸的风险[35]。条纹锯鮨从2002 年开始引入我国进行养殖,舟山市水产研究所于2016 年从青岛引入条纹锯鮨并开展养殖。条纹锯鮨非中国本地种,原产地为美国大西洋沿岸,因此海捕野生条纹锯鮨样品可能为养殖场逃逸的养殖鱼类,且捕捞地点距养殖场较近,为同一海域,但是否为养殖场鱼类,还需要进一步验证与讨论。
3.3 生物入侵的危害
外来物种进入自然环境后很难清除,外来物种可能导致本地物种被攻击甚至灭绝,严重破坏入侵地的生态系统,形成广泛的生物污染,危及土著群落的生物多样性并影响农业生产,造成了巨大的经济损失,对生物多样性和生态环境易造成无法修复的影响。一旦造成物种濒危和灭绝以及遗传资源的破坏,就再也无法恢复[35-36]。
鱼群逃逸在某种意义上来说本身就是一种生物入侵。特别是部分外来鱼类品种从网箱中出逃更令人担忧,逃逸的鱼类通常要经过较长的一段时间才对自然土著鱼类种群造成影响。这些外来物种主要是通过江河网箱或水族馆出逃到江河中,并通过掠食或摄食竞争严重时可造成当地种群灭绝、与当地种群杂交、掺杂基因库、破坏生境和引发流行病等,当其种群数量达到一定规模时,将威胁原有的鱼类生存空间,并破坏当地的生态平衡,降低当地水域的生物多样性[35]。
3.4 研究鱼类入侵方法及其防治策略
早期关于鱼类入侵的研究主要是针对入侵鱼类的形态描述,研究方法主要是分析水域调查的渔获组成。近些年来,随着研究的深入,各学科联合进一步推动了研究鱼类入侵方法的进步,数学模型、卫星遥感、遗传学等方法也逐步被应用于水生生物入侵当中[16]。本文采用的DNA 条形码技术,已经广泛地应用到鱼类鉴定当中,而应用于生物入侵领域,能够从分子层面的对入侵鱼类进行鉴定,更快更准确的确定其种类,从而对鱼类入侵提出针对性的防止措施。
综合人类社会、经济的发展来看,彻底隔绝外来种是不可能的,而适当地引进外来优良品种对我国的经济发展有促进作用[16],条纹锯鮨的引入与推广亦为我国的养殖行业带来了巨大的经济效益。但从环境的角度来看,我们需要谨慎对待引进物种,外来物种引进必须给与适量的监督和控制,否则会造成跟本地物种争夺资源,甚至排斥本地物种,条纹锯鮨的生长条件能够完全适应我国近海环境,会与本地物种争夺资源,破坏本地生境,污染基因库。因此我们需要建立健全外来引进种的管理机构,对其带来的利益与风险进行评估,同时也要加强对江海水域的生态环境的研究,建立水产外来物种的检测体系,及时发现生物入侵情况[37],最后要加强环保宣传教育,提高公众意识。
猜你喜欢
艺术启蒙(2020年6期)2020-07-16
中学生物学(2019年5期)2019-10-20
小福尔摩斯(2019年2期)2019-09-10
小学生学习指导(低年级)(2018年10期)2018-10-13
都市丽人(2017年4期)2017-04-12
都市家教·下半月(2016年11期)2016-12-29
现代农业科技(2016年20期)2016-12-20
科技知识动漫(2016年4期)2016-04-20
Coco薇(2016年3期)2016-04-06
现代企业(2015年2期)2015-02-28
