
水稻Wxmp背景下SSIIa和SSIIIa等位变异及其互作对蒸煮食味品质的影响
2020-09-24 13:46张亚东刘燕清赵春芳周丽慧赵庆勇BalakrishnaPillay王才林
作物学报 2020年11期
姚 姝 张亚东 刘燕清 赵春芳 周丽慧 陈 涛 赵庆勇 朱 镇 Balakrishna Pillay 王才林,*
水稻Wx背景下和等位变异及其互作对蒸煮食味品质的影响
姚 姝1张亚东1刘燕清1赵春芳1周丽慧1陈 涛1赵庆勇1朱 镇1Balakrishna Pillay2,*王才林1,*
1江苏省农业科学院粮食作物研究所/ 江苏省优质水稻工程技术研究中心 / 国家水稻改良中心南京分中心, 江苏南京 210014;2夸祖鲁-纳塔尔大学(西维尔校区)农业工程科学院生命科学系, 南非夸祖鲁-纳塔尔省德班市 4000
为了明确Wx基因背景下不同半糯粳稻品质差异的原因, 以淀粉合成酶基因和表现多态性而其他淀粉合成相关基因无多态性的武粳13和关东194 (Milky Princess)杂交后代衍生的64个半糯品系为材料, 分析了Wx基因背景下,和基因等位变异对直链淀粉含量(amylose content, AC)、胶稠度(gel consistency, GC)、糊化温度(gelatinization temperature, GT)及RVA谱特征值的影响。结果表明,和等位变异对AC、GC、GT和RVA谱特征值都有显著影响, 且2个基因间存在互作效应。2和2(2表示该基因来源于非半糯亲本武粳13)有使AC增高的趋势, 分别使AC提高1.87%和1.23%, 2年结果基本接近。单个和等位变异对GT无显著影响, 而基因型11(1表示该基因来源于半糯亲本关东194)的GT比22高1.34℃, 达显著水平, 表明2个基因的互作对GT有显著影响。GC在不同基因型间均存在极显著差异,2和1可分别使GC增加8.74 mm和9.62 mm。从2个基因的互作效应来看, 基因型21的GC比基因型12和22分别增加10.64 mm和16.95 mm。2使最高黏度、热浆黏度、冷胶黏度、崩解值增加, 回复值和消减值下降; 而2的效应则相反。2个基因的互作效应, 最高黏度、热浆黏度和冷胶黏度均以21最大, 崩解值和回复值均以22最大, 消减值21最小。本研究结果为半糯粳稻蒸煮食味品质的改良提供了一定的理论依据。
半糯粳稻;;; 蒸煮食味品质; 等位基因效应; 互作
水稻是我国的主要粮食作物之一, 长期以来一直以高产作为育种的主要目标。近年来, 随着产量和生活水平的提高及消费观念的变化, 人们对稻米品质有了更高的要求, 优质特别是优良食味品质成为育种的首要目标[1]。稻米直链淀粉含量是影响食味品质的关键因素。鄂志国等通过对1979—2018年我国省级以上审定的9563个水稻品种的系统研究发现, 长江中下游稻区中籼品种和黄淮海与长江下游稻区粳稻品种的直链淀粉含量明显降低, 食味品质提升[2]。低直链淀粉含量的稻米以其柔软、富有弹性的米饭质地特点, 越来越受到人们的青睐[3-5]。我们以含有半糯基因Wx的日本品种关东194 (Milky Princess)为亲本, 与江苏粳稻武香粳14、武粳13、武粳15等杂交, 育成了南粳46[6]、南粳5055[7]、南粳9108[8]、南粳3908[9]、南粳2728[10]等不同生育期类型的优良食味半糯粳稻系列品种, 在生产上已累计推广种植超过400万公顷, 生产的优质大米深受广大消费者欢迎。
我们在研究中发现, 在主效基因Wx背景下, 不同半糯水稻品种(系)间的直链淀粉含量仍有较大差异[11]。这除了与后期氮肥用量和灌浆充实期温度等环境因素有关以外[12], 主要是亲本所携带的淀粉合成相关基因的多态性所致。已经报道的与淀粉合成相关的淀粉合成酶基因有10个, 分别为()和[13]。其中,是控制稻米糊化温度(GT)的主效基因[14]。Umemoto等[15]将基因定位在水稻6号染色体短臂位点上, 并通过单倍型分析发现, GT主要受第8外显子的3个SNP组合突变类型控制, 等位变异类型主要分为A-GC、G-GC和G-TT三种组合类型[16]。Bao等[17]检测到在第8外显子中GC/TT的变异对糊化温度的变异具有较大影响。Waters等[18]测定了3个澳大利亚水稻品种的基因序列, 发现了4个SNPs, 其中第8外显子的2个SNPs可把70个品种分成高GT和低GT两种类型。
位于8号染色体上, 在支链淀粉的合成中, 负责长链B1和B2链(DP25-35)的合成, 且与其他淀粉合成酶存在互作[19-20]。Fujita等[21]研究表明,不仅对支链淀粉长链的延长起作用, 并且对SSI、GBSS和AGPase的表达起调节作用。杨博文等[22]研究表明,与基因的互作对水稻表观直链淀粉含量、糊化温度、最高黏度、崩解值和成糊温度有极显著影响, 且基因对基因具有显性上位性。还有研究表明,主要在胚乳中表达[23-25], 与稻米品质的关系比更密切。
本研究选择可溶性淀粉合成酶基因和表现多态性, 而其他淀粉合成酶基因相同的武粳13和关东194杂交后代衍生的半糯品系为材料, 利用分子标记, 对含有Wx主效基因的半糯品系进行和基因型鉴定, 分析Wx基因背景下,和等位变异对直链淀粉含量、胶稠度、糊化温度及RVA谱特征值的影响, 探讨在Wx基因背景下稻米蒸煮食味品质的变异原因, 为优良食味水稻新品种的选育提供理论依据。
1 材料与方法
1.1 供试材料
为了明确Wx基因背景下淀粉合成相关基因和对蒸煮食味品质的影响, 供试材料采用含有Wx基因的半糯水稻品系与、基因位点有差异的粳稻品种(Wx)杂交, 构建、、基因的不同基因型群体。
1999年以江苏省(武进)水稻研究所育成的粳稻品种武粳13为母本, 日本半糯水稻品种关东194 (Milky Princess)为父本杂交, 经多代自交获得的64个稳定品系为试验材料(图1)。64个品系的抽穗期均在8月下旬。在和位点上关东194与武粳13带有不同的等位基因[26], 利用分子标记关东194在和位点上可分别检测到344 bp和248 bp条带, 武粳13可分别检测到248 bp和177 bp条带。为方便起见, 将来源于关东194的基因型表示为“1”, 来源于武粳13的基因型表示为“2”。
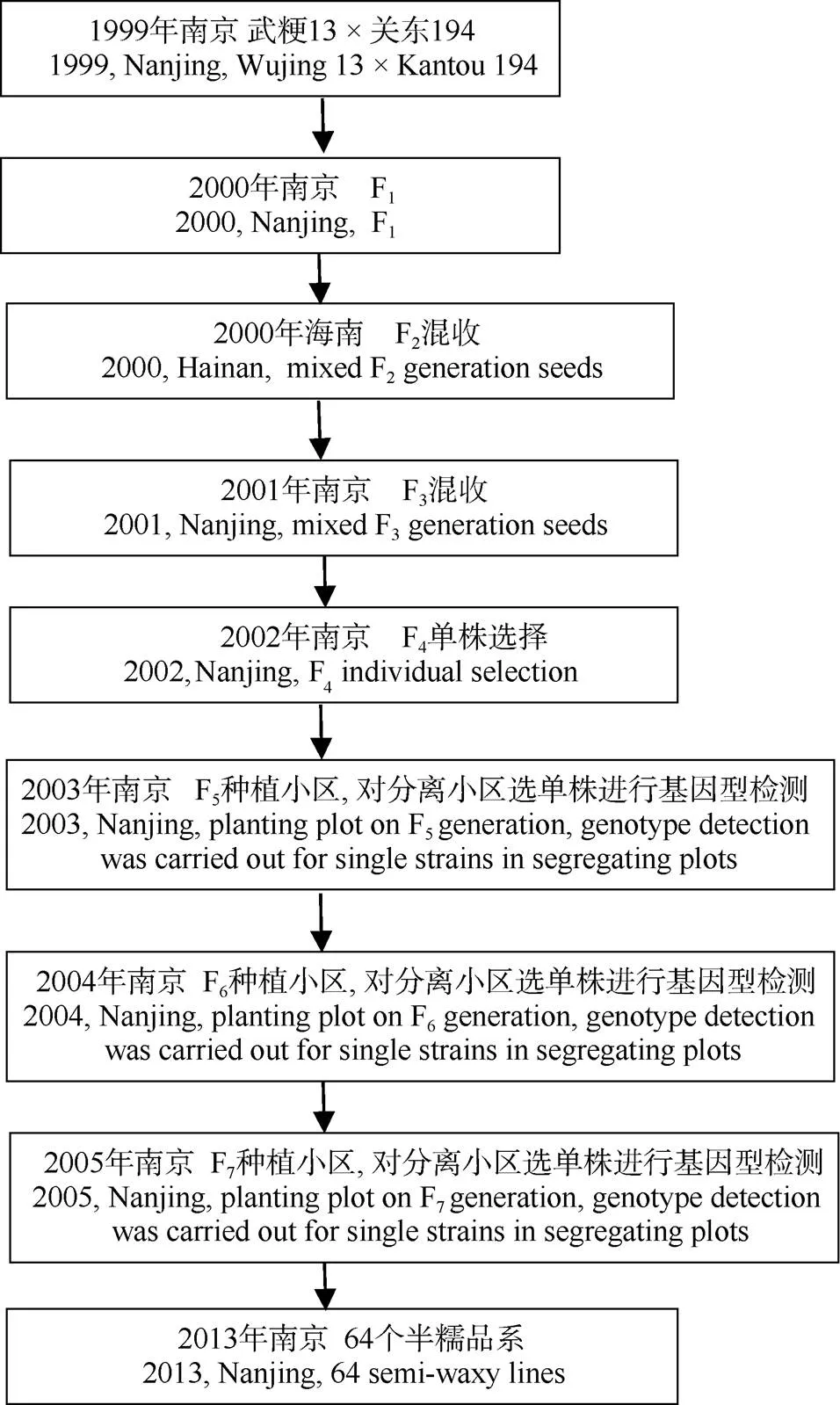
图1 来源于武粳13/关东194的64个半糯品系的培育
1.2 试验材料的种植与试验设计
试验材料分别于2013年和2014年正季种植在江苏省农业科学院粮食作物研究所试验田。每个株系种成1个小区, 每小区种植4行, 每行10株。行、株距分别为27 cm和17 cm。试验材料顺序排列, 重复3次。5月10日播种, 6月10日移栽。田间管理同大田生产。取分蘖盛期的幼嫩叶片用于DNA的提取, 待种子成熟后收获用于稻米理化指标和品质性状的测定。
1.3 基因型检测
在半糯品系培育过程中, 从F5开始, 分蘖盛期在各小区中间行选取5个单株进行Wx基因型鉴定, 直至获得稳定的半糯品系。2013年和2014年在对Wx基因型确认的基础上, 利用和基因的STS分子标记L22和L25进行基因型分型。
分蘖盛期在各小区中间行第2株开始, 连续取6个单株的新鲜幼嫩叶片, 用CTAB法提取DNA[27]。Wx基因的检测, 利用四引物扩增受阻突变体系PCR技术[28]。、基因的检测, 利用文献报道的淀粉合成途径中的调控基因及所用到的分子标记信息[29-31], 由上海英俊生物科技有限公司合成分子标记检测引物, 表1为用于半糯品系检测的淀粉合成相关基因的分子标记及其引物序列。根据PCR扩增产物的片段大小以及胶的分辨能力, 选择1.5%琼脂糖凝胶或者9%聚丙烯酰胺凝胶进行电泳检测。
1.4 稻米理化性状的测定
水稻种子成熟后, 在每小区中间随机收获5个单株, 风干至含水量14.5%左右时按单株脱粒。用韩国双龙砻谷机(SY88-TH)去壳出糙, 用台州伯利恒小型精米机(BLH-3120)出精米, 用瑞士产水分分析仪(Metteler)测定精米含水量, 用瑞典产FOSS旋风式磨粉机(CT193)研磨成米粉, 过100目网筛后, 用于品质性状测定。AC的测定按照农业部颁布标准NY147-88[32]进行, AC的4个标准样品(1.5%、10.4%、16.2%和26.5%)购自中国水稻研究所。每个样品测定3次。米粉胶稠度(GC)按国标GB/T 22294-2008[33]测定。用差示扫描量热仪DSC (200-F3, Netzsch German)测定的峰值温度(TP)表示糊化温度(GT)。每个性状每份样品均测定3次, 取平均值为性状表型值。
1.5 稻米RVA谱特征值的测定
采用澳大利亚Newport Scientific公司生产的3-D型黏度速测仪(RVA仪)及其配套软件TCW, 按照美国谷物化学协会操作规程进行测定。黏度单位用cP表示。淀粉黏度参数有5个一级数据和3个二级数据, 一级数据包括峰值黏度(peak viscosity, PKV)、热浆黏度(hot viscosity, HPV)、冷浆黏度(cool paste viscosity, CPV)、起浆温度(pasting temperature, PaT)、峰值时间(peak time, PeT); 二级参数有: 崩解值(breakdown viscosity, BDV = PKV-HPV)、消减值(setback viscosity, SBV = CPV-PKV)和回复值(consistency viscosity, CSV = CPV-HPV)。每个样品测定3次, 取平均值为性状表型值。

表1 SSIIa、SSIIIa和Wxmp基因分子标记
1.6 统计分析
按照莫惠栋[34]介绍的方法进行方差分析和差异显著性测验, 多重比较采用Duncan’s新复极差法。
2 结果与分析
2.1 供试材料的基因型检测结果
2.1.1Wx基因型的检测 根据孟德尔遗传原理, 携带基因的粳稻与携带Wx基因的半糯粳稻杂交后, 分离世代有、WxWx和Wx3种基因型。利用四引物受阻扩增突变体系PCR系统可以同时检测3种基因型[28]。结果表明, 64个半糯品系与2个亲本都能扩增出439 bp的特异性条带。另外, 64份半糯品系与关东194还能扩增出292 bp的特异性条带, 而不含Wx的武粳13不能扩增出292 bp条带, 但能扩增出一条200 bp的特异条带(图2), 表明这64份供试材料都含有Wx基因。

图2 64个半糯品系Wxmp基因的检测
M: marker; 1~7: 部分半糯品系; 8: 关东194; 9: 武粳13。
M: marker; 1–7: part of the semi-waxy lines; 8: Kantou 194;9: Wujing 13.
2.1.2和基因型的检测 利用和基因位点设计的STS分子标记L22和L25对64个半糯品系进行检测。如果含有来源于半糯亲本关东194的和基因, 就能分别检测到251 bp和197 bp条带, 如果含有来源于非半糯亲本武粳13的和基因, 就能分别检测到248 bp和177 bp条带(图3)。检测结果表明, 检测到1(1表示该基因来源于半糯亲本关东194, 下同)和2(2表示该基因来源于非半糯亲本武粳13)基因型的品系分别为23个和41个,1和2基因型的品系分别为49个和15个。2个基因的4种基因型11、12、21和22的品系分别为10个、13个、39个和2个(表2)。

图3 64个半糯品系SSIIa (A)和SSIIIa (B)基因的检测
1~7: 部分半糯品系; 8: 关东194; 9: 武粳13。
1–7: part of the semi-waxy lines; 8: Kantou 194; 9: Wujing 13.
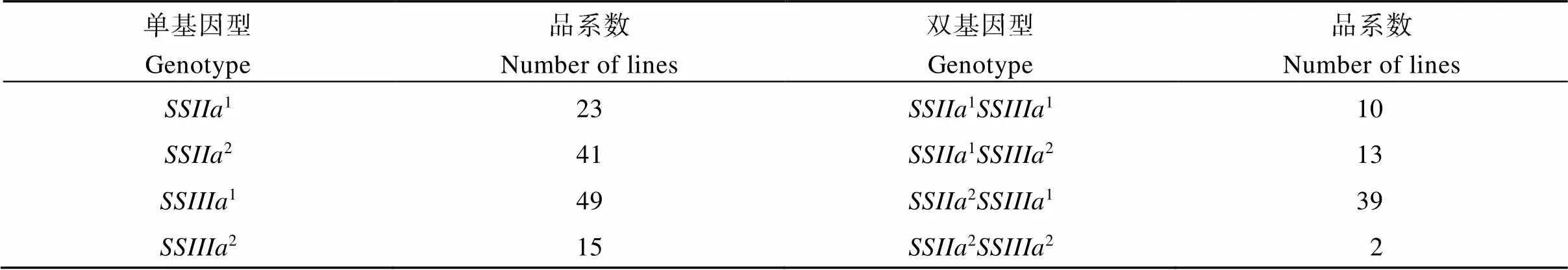
表2 来源于武粳13/关东194的64个半糯品系SSIIa和SSIIIa位点的基因型
1表示该基因来源于半糯亲本关东194,2表示该基因来源于非半糯亲本武粳13。
1indicated that the allele was derived from the semi-waxy parent of Kantou 194, and2indicated that the allele was derived from the parent of Wujing 13.
2.2 Wxmp背景下不同SSIIa和SSIIIa基因型蒸煮食味品质的变异
2.2.1 直链淀粉含量、胶稠度和糊化温度的变异 对直链淀粉含量(AC)、糊化温度(GT)以及胶稠度(GC)进行测定的结果表明(图4), 64个半糯品系的AC分布在7.0%~11.0%之间, 其中分布在8.0%~ 10.0%之间的有54份, 占测定总数的85.71%。GT主要分布在68.2~73.0℃之间, 主要集中在69~71℃,但总体变化不大, 变异系数为1.56%。GC主要分布在50.0~102.0 mm之间, 平均值为79.6 mm, 变异系数为16.05%。

图4 64个半糯品系AC、GC、GT的频率分布
AC: 直链淀粉含量; GC: 胶稠度; GT: 糊化温度。
AC: amylose content; GC: gel consistency; GT: gelatinization temperature.
2.2.2 RVA谱特征值的变异 64个半糯品系RVA谱特征值的分布见图5。最高黏度的变异幅度在1746~3511 cP之间, 其中分布在2099~3158 cP之间的有52份, 占测定总数的82.5%。热浆黏度的变异幅度在925~2160 cP之间, 其中分布在1172~1913 cP之间的有51份, 占测定总数的81.0%。冷胶黏度的变异幅度在1533~2769 cP之间, 其中分布在1780~2521 cP之间的有50份, 占测定总数的79.4%。崩解值的变异幅度在617~1646 cP之间, 其中分布在822~1646 cP之间的有60份, 占测定总数的95.2%。消减值的变异幅度在–1018~-1 cP之间, 分布在各区间的相对均匀。回复值的变异幅度在480~752 cP之间, 其中分布在588~752 cP之间的有56份, 占测定总数的88.9%。其中消减值的变异幅度最大, 其变异系数达到了42.4%, 其次为崩解值、热浆黏度、最高黏度、冷胶黏度和回复值。
2.3 SSIIa和SSIIIa不同等位基因蒸煮食味品质的效应分析
2.3.1 直链淀粉含量 对2013年和2014年种植的64个半糯品系的AC进行方差分析的结果见表3。AC在64个品系间、年份间和年份与品系的互作间都存在极显著的差异。进一步分析表明, 尽管AC在11、12、21和22四种基因型内的差异均达到极显著水平, 但4种基因型间的差异更大。除了基因型11与12间的AC差异不显著以外, 其余基因型间AC的差异均达极显著水平。
表4列出了2013—2014年64个半糯品系和位点不同基因型AC的平均值。无论是位点还是位点, 其AC均是非半糯亲本基因型高于半糯亲本基因型。平均而论,2基因型的AC比SSIIa基因型高出1.87个百分点,2基因型的AC比SSIIIa基因型高出1.23个百分点, 表明的效应大于。
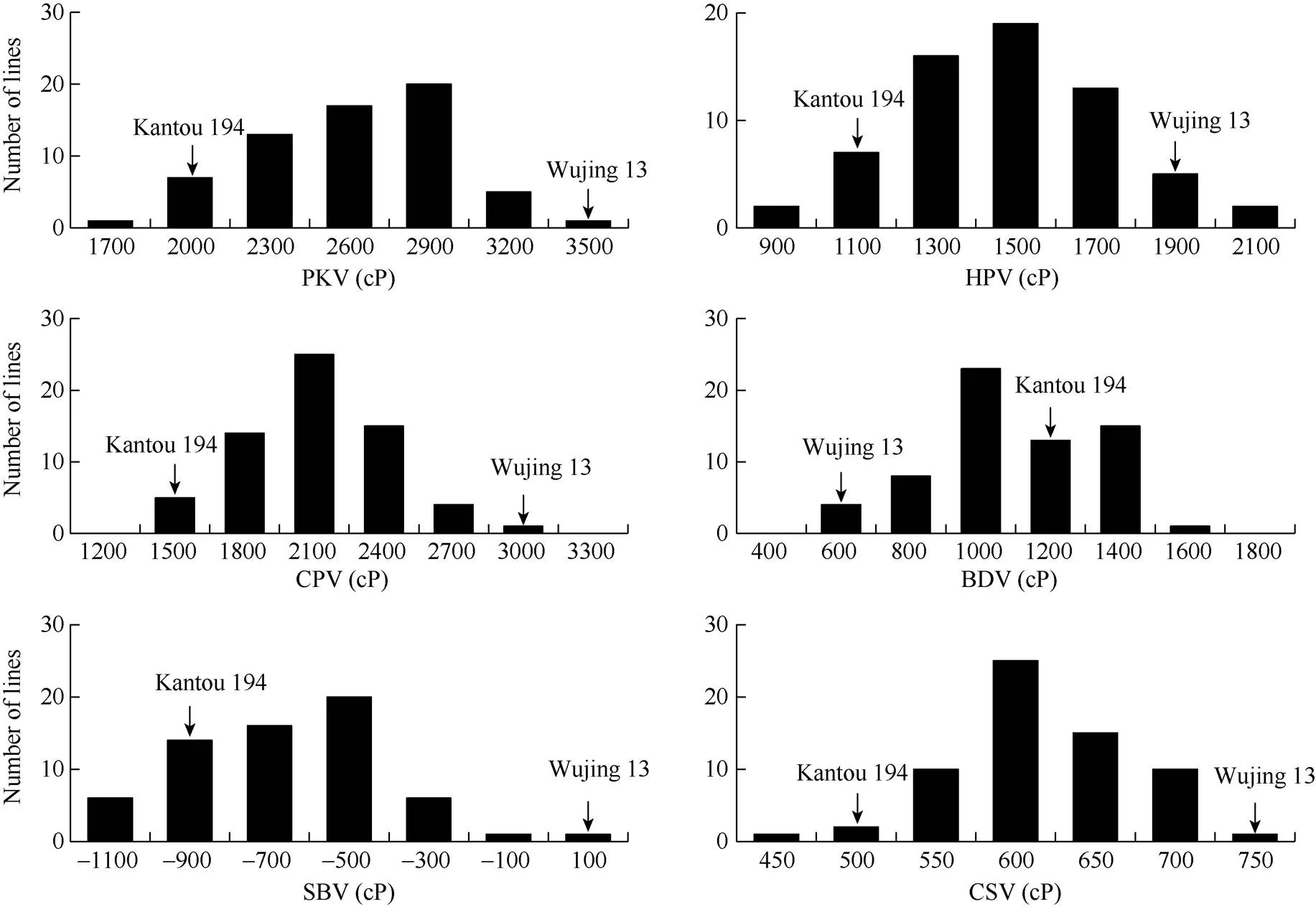
图5 64个半糯品系RVA特征值的频率分布
PKV: 峰值黏度; HPV: 热浆黏度; CPV: 冷浆黏度; BDV崩解值; SBV消减值; CSV回复值。
PKV: peak viscosity; HPV: hot paste viscosity; CPV: cool paste viscosity; BDV: breakdown viscosity; SBV: setback viscosity; CSV: consistence viscosity.
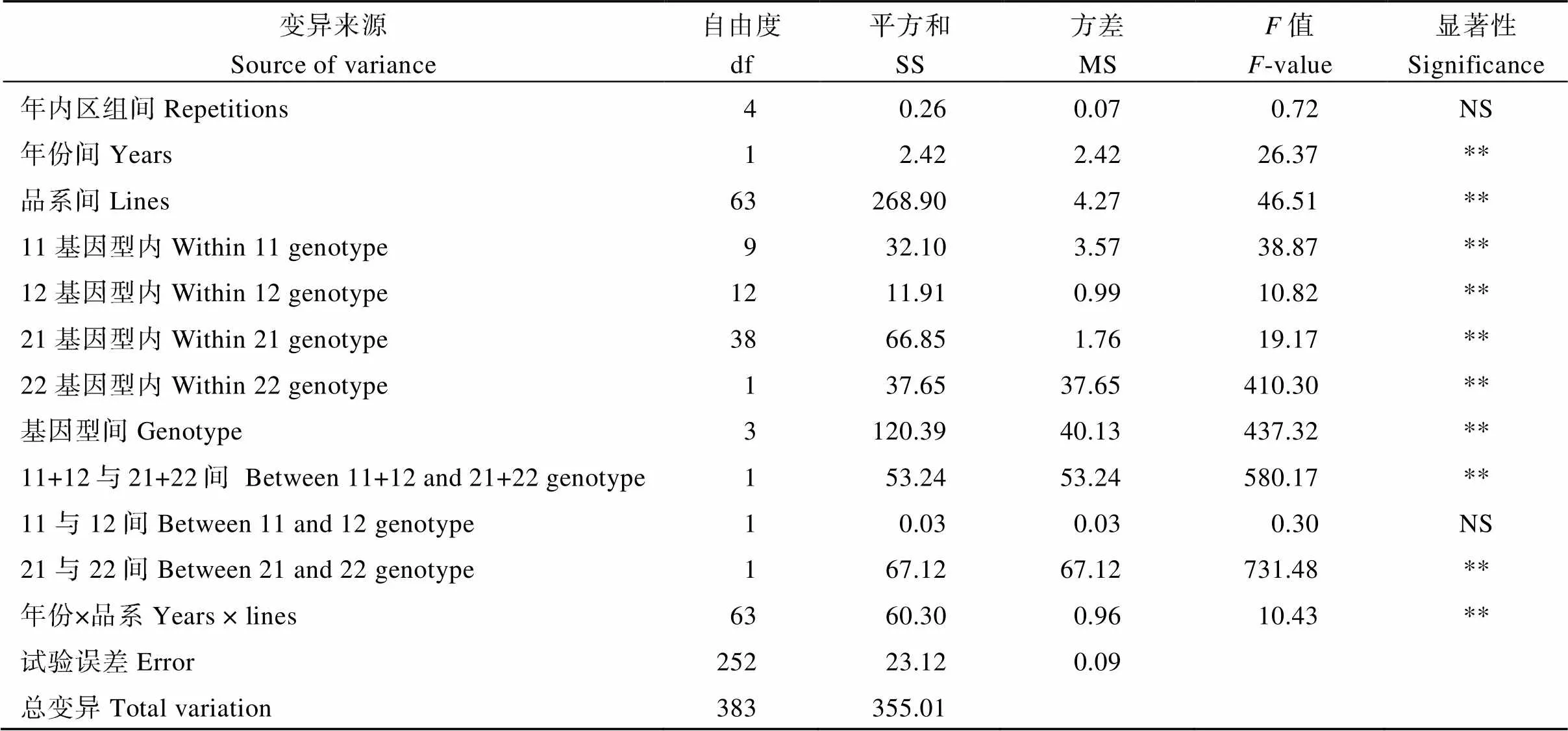
表3 64个半糯品系SSIIa和SSIIIa位点不同基因型不同年份AC的方差分析
11、12、21和22基因型分别表示11、2、21和22基因型。**: 1%水平显著, NS: 不显著。
Genotype 11, 12, 21, and 22 represent the four genotypes of11,12,21, and22, respectively. **: significant at the 1% probability level; NS: not significant.
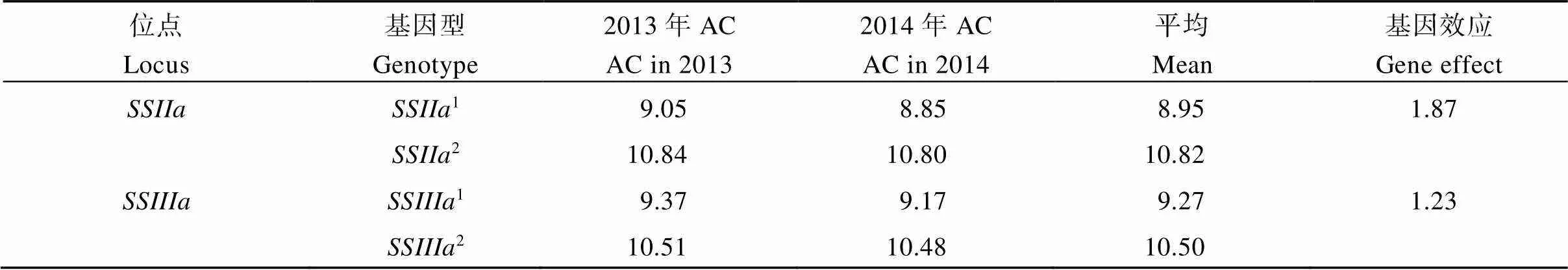
表4 2013年和2014年64个半糯品系SSIIa和SSIIIa位点不同等位基因对AC的效应
AC: 直链淀粉含量。AC: amylose content.
表5列出了2013—2014年64个半糯品系和位点4种基因型AC的平均值、最大值、最小值及年份间的差异。4种基因型的AC平均值分为3类, 基因型11和12的AC最低, 两者间无显著差异; 基因型22的AC最高, 基因型21的AC居中, 且3类的AC均达极显著水平。从表4的单基因遗传效应来看, 来源于非半糯亲本基因型的AC均高于半糯亲本基因型。但在1基因背景下,2的AC并不比1高, 表明和位点间存在互作。
2.3.2 糊化温度与胶稠度 不同基因型的糊化温度和胶稠度数据列于表6。不同亲本来源的和基因型间的糊化温度均无显著差异, 而不同基因型间的胶稠度均有极显著差异, 来源于武粳13的基因和来源于关东194的基因分别使胶稠度增加了8.74 mm和9.61 mm。
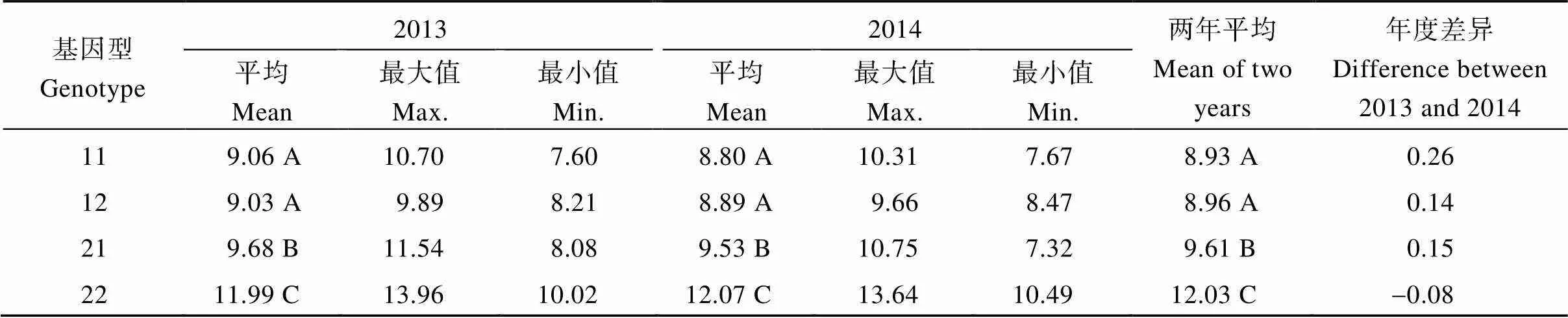
表5 64个半糯品系SSIIa和SSIIIa位点不同基因型不同年份AC的差异
11、12、21和22基因型分别表示11、12、21和22基因型。同一列不同大写字母表示差异达1%显著水平。
Genotype 11, 12, 21, and 22 represent the four genotypes of11,12,21, and22, respectively. Different uppercase letters in the same column indicate significant differences at the 1% probability level.

表6 64个半糯品系SSIIa和SSIIIa位点不同基因型糊化温度与胶稠度的差异
**, 1%显著水平; 同一列不同字母表示在5%水平上差异显著。
**, significant at the 1% probability level; data followed by different lowercase letters in the same column indicated significant at the 5% probability level.
多重比较结果显示, 基因型11的糊化温度最高, 其次为基因型21、12和22, 但是4种基因型的糊化温度比较接近。除了基因型11与22之间有显著差异外, 其余基因型间均无显著差异。基因型21的胶稠度最高, 依次为基因型11、12和22, 基因型21与12和22之间达到显著差异, 其余基因型相互之间均未达到显著差异。
2.3.3 RVA谱特征值 不同基因型RVA谱特征值的测定结果见表7。对于基因而言, 亲本来源不同的基因型间的特征值都有显著或极显著差异; 同样对于基因而言, 除了热浆黏度和冷胶黏度亲本来源不同的基因型间的差异不显著外, 最高黏度、崩解值、回复值以及消减值在亲本来源不同的基因型间都具有显著或极显著差异。

表7 64个半糯品系SSIIa和SSIIIa位点不同基因型RVA特征值的差异
*,**或同一列数据后的不同大、小写字母分别表示在1%和5%水平差异显著。
PKV: peak viscosity; HPV: hot paste viscosity; CPV: cool paste viscosity; BDV: breakdown viscosity; CSV: consistence viscosity; SBV: setback viscosity. Data followed by*,**or different uppercase and lowercase letters in the same column indicate significant at the 1% and 5% probability levels, respectively.
进一步对2个基因的4种基因型间RVA谱各特征值进行多重比较的结果显示, 4种基因型间各特征值都存在显著或极显著的差异。当基因的亲本来源都为关东194 (即基因型为1)时, 不同亲本来源的基因型(即1与2)之间, 各特征值的差异均不显著; 当基因的亲本来源都为武粳13 (即基因型为2)时, 除了回复值在不同亲本来源的基因型(即1与2)之间的差异极显著外, 其他特征值均无显著差异。当基因的亲本来源都为关东194 (即基因型为1)时, 除了回复值在不同亲本来源的基因型(即1与2)之间的差异不显著外, 其他特征值均具有显著或极显著差异; 当基因的亲本来源都为武粳13 (即基因型为2)时, 不同亲本来源的基因型(即1与2)之间, 最高黏度、热浆黏度和冷胶黏度的差异不显著, 消减值的差异显著, 崩解值和回复值的差异极显著。由此可见, 影响RVA谱特征值的基因主要是。半糯品系的最高黏度和崩解值大于亲本武粳13, 热浆黏度、冷胶黏度、回复值以及消减值均小于亲本武粳稻13。隋炯明等[35]研究认为RVA谱特征值与食味品质指标间存在极为密切的关系, 说明以RVA谱特征值反映稻米食味品质优劣的可行性。进一步研究表明, 崩解值、消减值、回复值对评价食味品质的优劣较为准确, 可以作为选择食味品质优良材料的有效指标。崩解值较高且消减值和回复值较低的品种, 其蒸煮食味品质较好。本试验中崩解值较高的是21和22两种基因型, 两者没有显著差异。回复值较低的是21和11, 两者没有显著差异, 消减值较低的是21和22两种基因型, 两者也没有显著差异。由此推断基因型为21的品种, 其蒸煮食味品质较好, 其次为22基因型。
3 讨论
3.1 SSIIa和SSIIIa的遗传效应
研究表明, 稻米蒸煮食味品质除了受主效基因控制外, 还受到其他淀粉合成相关基因的调控[36]。可溶性淀粉合成酶基因也被称为糊化温度控制基因[37], 负责将支链淀粉的短支链(A+B1)合成至中等长度的分支链(B2+B3)。据报道,基因有3种单倍型, 高GT的G-GC单倍型、低GT的G-TT和A-GC单倍型[37-38]。我们在前期的研究表明, 武粳13和关东194在和位点分别含有不同的等位基因[26]。本研究对来源于武粳13/关东194的64个半糯品系进行基因型分类研究的结果表明, 不同亲本来源的和等位基因对AC、GT、GC和RVA谱特征值都有影响。来源于非半糯亲本武粳13的和基因有使AC增高的趋势, 分别使AC提高1.87个和1.23个百分点, 2年结果基本接近。不同亲本来源的或等位基因对糊化温度无显著影响, 而基因型11的糊化温度显著高于22, 表明虽然单个和基因等位变异对糊化温度无显著影响, 但2个基因同时存在时对糊化温度有显著影响, 基因型的糊化温度比22高1.34℃。胶稠度在不同基因型间均存在极显著差异, 来源于武粳13的基因和来源于关东194的基因可分别使胶稠度增加8.74 mm和9.62 mm。从2个基因的联合效应来看, 基因型21的胶稠度最高, 比基因型12和22分别增加10.64 mm和16.95 mm, 差异达到显著水平。
一般认为,是控制糊化温度的主效基因。本研究表明等位变异对糊化温度无显著影响, 但与基因同时存在时对糊化温度有显著影响; 此外,等位变异对AC也有显著影响, 这与以往研究结果不尽一致。我们分析,和等基因都是微效基因, 与主效基因相比, 效应要小得多, 而糊化温度在粳稻中的差异较小, 在半糯粳稻中的差异更小, 可能是主效基因Wx掩盖了基因的作用。此外,和基因在不同主效基因(如Wx,Wx,Wx)背景下的效应也不同, 即基因之间存在相互作用。实际上, Gao等[37]的研究虽然证实可溶性淀粉合成酶基因是控制糊化温度的主要基因, 但对AC、GC和淀粉的糊化特性也有影响。Tian等[39]关联分析结果也表明,()对AC、GC和GT都有影响, 此外,、()、和对AC也有影响。He等[40]利用“NJ11/Balilla”的DH群体进行研究, 除了主效基因外, 也检测到了、和基因对AC、GC、GT的遗传效应。因此, 关于和等淀粉合成相关基因对食味品质的影响机制值得进一步研究。
RVA谱特征值尤其是崩解值、回复值和消减值可以反映水稻品种间蒸煮食味的差异, 可以作为评价稻米食味品质的辅助手段。吴洪凯等[41]的研究表明, 糯稻中的基因对RVA谱特征值有重要影响。本研究中不同亲本来源的等位基因间, 各特征值几乎都有显著或者极显著差异; 而等位基因间, 影响品质的几个主要的特征值都无显著差异, 说明对RVA谱特征值的作用大于。隋烔明等[35]认为具有较低的热浆黏度、冷胶黏度、消减值、回复值、较短的峰值时间, 较低的起浆温度以及较高的崩解值, 可能具有较好的柔软性、粘性、冷饭质地和较高的综合评分。舒庆尧等[42]认为RVA谱特征值中最高黏度、崩解值、消减值以及回复值可以反映食味品质的优劣, 不管是籼稻还是粳稻品种, 具有相对较高的最高黏度和崩解值、相对较低的回复值和消减值的品种, 其食味品质相对较好。本研究中所有半糯品系的崩解值几乎都大于亲本武粳13, 最高黏度, 热浆黏度, 冷胶黏度, 回复值和消减值均小于亲本武粳13, 而半糯品系具有较好的柔软性、弹性和粘性, 这与前人研究一致。
参与淀粉合成的酶数目众多, 包括颗粒淀粉合成酶(granule-bound starch synthase, GBSS), 可溶性淀粉合成酶(soluble starch synthase, SSS)、分支酶(starch branching enzyme, SBE)、脱分支酶(starch debranching enzyme, DBE)等, 其对应的基因有20多个[36,41,43]。因此, 淀粉的合成是由一系列淀粉合成酶的协同作用来完成, 不同的基因之间存在互作效应, 基因间的相互作用比较复杂且多样, 1个性状的改变大多是由多个基因所控制的几个生化过程的连锁反应的结果, 基因之间形成了复杂的调控网络。本研究分析了和基因对稻米蒸煮食味品质的效应, 作者最近还报道了和基因等位变异的效应[44], 对于其他淀粉合成相关基因对稻米蒸煮食味品质的效应有待于进一步研究。
3.2 SSIIa与SSIIIa间的互作效应
生物体的各个性状都由一系列的基因所调控, 形成一个复杂的基因调控网络。不同基因之间必然存在相互作用。水稻淀粉的生物合成途径已经非常明确, 直链淀粉和支链淀粉共用底物ADP葡萄糖在不同酶催化下分别合成, 2条合成途径存在竞争关系[45]。直链淀粉由颗粒结合淀粉合成酶催化合成,而支链淀粉由可溶性淀粉合成酶、淀粉分支酶和淀粉去分支酶协同合成。参与编码这些酶的基因有20多个, 包括10个编码淀粉合成酶的基因(); 6个编码淀粉分支酶和去分支酶的基因、等[46-47]。He等[40]在“NJ11/Balilla”的DH群体中曾检测到较多的淀粉合成酶基因相关间的互作效应。本研究涉及和淀粉合成酶基因。从单基因效应看, 无论是位点还是位点, AC均是非半糯亲本武粳13的基因型高于半糯亲本关东194的基因型。2年平均基因效应,和分别为1.87个和1.23个百分点(表4)。但在1基因背景下,的平均基因效应只有0.03个百分点, 而在2基因背景下,的平均基因效应高达2.42个百分点。同样, 在1基因背景下,的平均基因效应只有0.67个百分点, 而在2基因背景下,的平均基因效应高达3.07个百分点(表5)。表明与2基因间存在互作效应(图6)。
同样, GT、GC、最高黏度、热浆黏度、冷胶黏度、崩解值、回复值和消减值在不同亲本基因型背景下和的遗传效应均不完全相同(表8)。除了GT在不同亲本基因型背景下和的遗传效应差异均不显著外, 其余性状的差异均达显著水平, 表明这些性状在和基因间均存在互作效应。
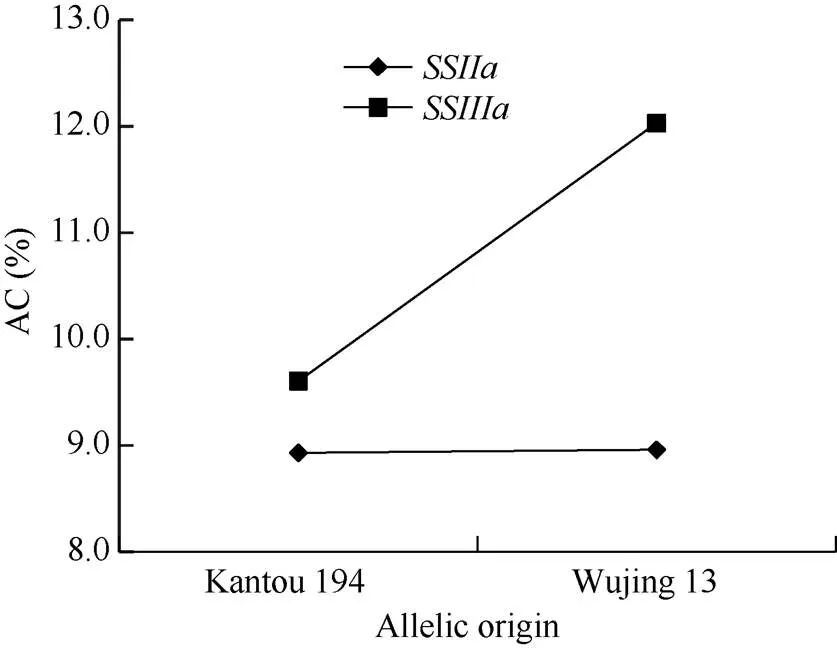
图6 SSIIa和SSIIIa位点不同基因型间的互作
AC: 直链淀粉含量。AC: amylose content.
3.3 本研究结果在优良食味粳稻品质改良中的应用
半糯粳稻因具有较低的直链淀粉含量而呈现出乳白色、云雾状不透明的表型, 这对稻米外观品质具有不利影响, 但不同半糯基因的直链淀粉含量不同, 因此, 半糯基因的选择对培育优良食味粳稻有重要意义。我们的育种实践表明, 含半糯基因的稻米食味品质虽然总体上均较好, 但半糯粳稻的直链淀粉含量也不是越低越好。直链淀粉含量过低(低于8%), 米粒不透明, 外观品质变差, 口感太软, 缺乏弹性; 在半糯基因背景下适当提高直链淀粉含量, 可以使外观品质得到改善, 而食味品质保持不变。当直链淀粉含量达到12%以上, 不仅米饭柔软性好, 富有弹性, 冷而不硬, 口感润滑, 符合长三角地区人们喜食软糯粳米的口感需求, 而且外观品质变好(图7)。来源于日本粳稻越光的EMS突变体Wx的直链淀粉含量在9%左右, 但即使来源于含Wx基因的同一组合的不同半糯品系之间, 其直链淀粉含量仍有很大差异, 表明除了位点以外, 还有其他基因影响直链淀粉的合成。本研究发现在半糯基因Wx背景下, 武粳13来源的和等位基因可分别使直链淀粉含量提高1.87和1.23个百分点, 同时拥有这2基因则可以使直链淀粉含量提高3.1个百分点, 达到12%左右。这样的半糯品系不仅食味品质优良, 而且外观透亮。因此, 通过和不同等位基因型的选择, 可以使半糯粳稻既好吃又好看。

表8 不同亲本基因型背景下SSIIa和SSIIIa的遗传效应
AC: 直链淀粉含量; GT: 糊化温度; GC: 胶稠度。缩写同表7。*表示在= 0.05水平上差异显著。
AC: amylose content; GT: gelatinization temperature; GC: gel consistency. Abbreviations were the same as those given in Table 7.*-values are significant at= 0.05.
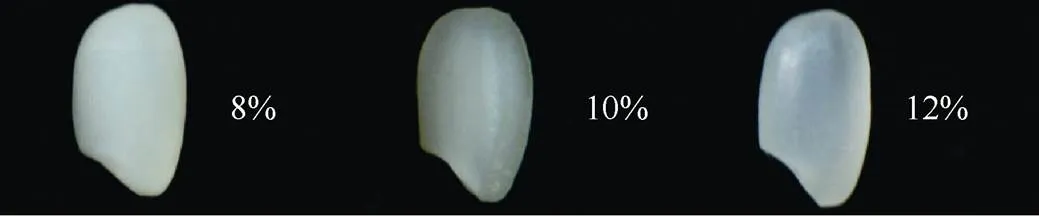
图7 不同直链淀粉含量半糯粳稻米粒的外观品质
当然, 影响稻米品质的基因还有很多。研究某个基因的遗传差异或效应, 必须在其他基因背景相同的情况下进行, 最好是利用近等基因系或通过基因编辑研究其功能。但实际育种工作中所遇到的常常是1个多基因系统控制下的复杂个体。相对于同一背景下特定地研究某个基因的遗传效应或某些基因之间的互作效应, 在复杂遗传背景下分析不同基因间的联合效应和变异趋势更符合育种实际, 而目前有关这方面的研究报道不多。本研究采用来源于和等位基因有差异、其他淀粉合成相关基因相同的武粳13和关东194杂交后代衍生的半糯品系为材料, 在Wx基因背景下, 研究和等位基因的遗传效应及其互作对食味品质的影响, 用以指导实际育种。同时, 为了尽可能排除生育期、环境因素等的影响, 供试材料都选取生育期相近(8月22日至30日抽穗)的64份材料。由于选择过程中偏向于品质和产量等性状的选择, 4种基因型品系数的分布不均衡, 最少的基因型只有2个, 难免对研究结果有一定影响, 但其变异趋势仍然具有参考价值。我们正是通过这些遗传信息的利用, 在南粳46、南粳5055和南粳9108之后, 选出了符合市场需要, 受农民欢迎的南粳晶谷、南粳5718、南粳2728等南粳系列后续品种, 这些品种在保持优良食味品质的基础上, 外观品质、抗性和产量性状均有提高。通过对更多品质相关基因遗传效应的研究, 可以为培育更多符合市场需要、食味品质优良的理想基因型的优质水稻品种提供理论依据。
4 结论
在Wx基因背景下, 淀粉合成酶基因和等位变异对直链淀粉含量(AC)、胶稠度(GC)、糊化温度(GT)及RVA谱特征值有显著影响, 且2个基因间存在互作效应。2和2分别使AC提高1.87%和1.23%。基因型11的糊化温度比22高1.34℃,2和1分别使胶稠度增加8.74 mm和9.62 mm, 基因型21的胶稠度比基因型12和22分别增加10.64 mm和16.95 mm。2使最高黏度、热浆黏度、冷胶黏度、崩解值增加, 回复值和消减值下降; 而2的效应则相反。最高黏度、热浆黏度和冷胶黏度均以21最大, 崩解值和回复值均以22最大, 消减值21最小。综合各性状的效应, 基因型21的直链淀粉含量适中, 胶稠度最高, 最高黏度、崩解值最大, 回复值和消减值最小, 蒸煮食味品质最佳。和基因的等位变异及其互作效应对半糯粳稻蒸煮食味品质的改良有重要参考价值。
[1] 王才林, 张亚东, 朱镇, 陈涛, 赵庆勇, 赵凌, 周丽慧, 姚姝, 赵凌, 李余生. 水稻优质抗病高产育种的研究与实践. 江苏农业学报, 2012, 28: 921–927. Wang C L, Zhang Y D, Zhu Z, Chen T, Zhao Q Y, Zhou L H, Yao S, Zhao L, Li Y S. Studies and practice on the development ofrice varieties with good quality, disease resistance and high yield.2012, 28: 921–927 (in Chinese with English abstract).
[2] 鄂志国, 程本义, 孙红伟, 汪玉军, 朱练峰, 林海, 王磊, 童汉华, 陈红旗. 近40年我国水稻育成品种分析. 中国水稻科学, 2019, 33: 523–531. E Z G, Cheng B Y, Sun H W, Wang Y J, Zhu L F, Lin H, Wang L, Tong H H, Chen H Q. Analysis on Chinese improved rice varieties in recent four decade., 2019, 33: 523–531 (in Chinese with English abstract).
[3] Suto M, Ando I, Numaguchi K. Breeding of low amylose content paddy rice variety Milk Queen with good eating quality., 1996, 46: 221–224.
[4] 黄祖六, 许如根, 陈德辉, 陈勇. 中泰软米资源直链淀粉含量的遗传研究. 扬州大学学报(农业与生命科学版), 2003, 24: 34–36.Huang Z L, Xu R G, Chen D H, Chen Y. Research on inheritance for amylose content of soft rice from China and Thailand.(Agric Life Sci Edn), 2003, 24: 34–36 (in Chinese with English abstract).
[5] 赵春芳, 岳红亮, 黄双杰, 周丽慧, 赵凌, 张亚东, 陈涛, 朱镇, 赵庆勇, 姚姝, 梁文化, 路凯, 王才林. 南粳系列水稻品种的食味品质与稻米理化特性. 中国农业科学, 2019, 52: 909–920. Zhao C F, Yue H L, Huang S J, Zhou L H, Zhao L, Zhang Y D, Chen T, Zhu Z, Zhao Q Y, Yao S, Liang W H, Lu K, Wang C L. Study on eating quality and physicochemical properties in Nanjing rice varieties., 2019, 52: 909–920 (in Chinese with English abstract).
[6] 王才林, 陈涛, 张亚东, 朱镇, 赵凌, 林静. 通过分子标记辅助选择培育优良食味水稻新品种. 中国水稻科学, 2009, 23: 25–30. Wang C L, Chen T, Zhang Y D, Zhu Z, Zhao L, Lin J. Breeding of a new rice variety with good eating quality by marker assisted selection., 2009, 23: 25–30 (in Chinese with English abstract).
[7] 王才林, 张亚东, 朱镇, 陈涛, 赵庆勇, 赵凌, 周丽慧, 姚姝. 优良食味粳稻新品种南粳5055的选育及利用. 农业科技通讯, 2012, (2): 84–87. Wang C L, Zhang Y D, Zhu Z, Chen T, Zhao Q Y, Zhao L, Zhou L H, Yao S. Breeding and application of new good eating quality rice variety Nanjing 5055., 2012, (2): 84–87 (in Chinese with English abstract).
[8] 王才林, 张亚东, 朱镇, 姚姝, 赵庆勇, 陈涛, 周丽慧, 赵凌. 优良食味粳稻新品种南粳9108的选育与利用. 江苏农业科学, 2013, 41: 86–88. Wang C L, Zhang Y D, Zhu Z, Yao S, Zhao Q Y, Chen T, Zhou L H, Zhao L. Breeding and application of new good eating quality rice variety Nanjing 9108., 2013, 41: 86–88 (in Chinese with English abstract).
[9] 陈涛, 张亚东, 赵庆勇, 朱镇, 姚姝, 周丽慧, 赵凌, 赵春芳, 王波, 王才林. 优良食味抗病高产晚粳稻新品种南粳3908的选育和栽培技术. 江苏农业科学, 2019, 47: 72–74. Chen T, Zhang Y D, Zhao Q Y, Zhu Z, Yao S, Zhou L H, Zhao L, Zhao C F, Wang B, Wang C L. Breeding and cultivation technique of a newrice variety Nanjing 3908 with good eating quality, resistant diseases and high yield., 2019, 47: 72–74 (in Chinese with English abstract).
[10] 蔡锋, 吴俊生, 王才林. “南粳2728”特征特性及栽培技术要点. 上海农业科学, 2019, (373): 44. Cai F, Wu J S, Wang C L. Characteristics and cultivation techniques of “Nanjing 2728”., 2019, (373): 44 (in Chinese).
[11] 于新, 赵庆勇, 赵春芳, 张亚东, 朱镇, 赵凌, 陈涛, 周丽慧, 姚姝, 王才林. 携带基因的不同类型水稻新品种(系)直链淀粉含量分析. 江苏农业学报, 2012, 28: 1218–1222. Yu X, Zhao Q Y, Zhao C F, Zhang Y D, Zhu Z, Zhao L, Chen T, Zhou L H, Yao S, Wang C L. Analysis of amylose content in different types of new rice varieties (lines) carryinggene., 2012, 28: 1218–1222 (in Chinese with English abstract).
[12] 姚姝, 于新, 周丽慧, 陈涛, 赵庆勇, 朱镇, 张亚东, 赵春芳, 赵凌, 王才林. 氮肥用量和播期对优良食味粳稻直链淀粉含量的影响. 中国水稻科学, 2016, 30: 535–549. Yao S, Yu X, Zhou L H, Chen T, Zhao Q Y, Zhu Z, Zhang Y D, Zhao C F, Zhao L, Wang C L. Effects of nitrogen and sowing date on amylose content in good eating quality rice (L.)., 2016, 30: 535–549 (in Chinese with English abstract).
[13] Sato H. Genetics and breeding of high eating quality rice: status and perspectives on the researches of low amylose content rice., 2002, 77: 20–28.
[14] Gao Z Y, Zeng D L, Cui X, Zhou Y H, Yan M X, Huang D N, Li J Y, Qian Q. Map-based cloning of thegene, which controls the gelatinization temperature of rice.-, 2003, 46: 661–668.
[15] Umemoto T, Yano M, Satoh H, Shomura A, Nakamura Y. Mapping of a gene responsible for the difference in amylopectin structure between-type and-type rice varieties., 2002, 104: 1–8.
[16] Umemoto T, Aoki N. Single-nucleotide polymorphisms in ricethat alter starch gelatinisation and starch association of the enzyme.,2005, 32: 763–768.
[17] Bao J S, Corke H, Sun M. Nucleotide diversity inand validation of single nucleotide polymorphisms in relation to starch gelatinization temperature and other physicochemical properties in rice (L.).,2006, 113: 1171–1183.
[18] Waters D, Henry R J, Reinke R F, Fitzgeraid M A. Gelatinization temperature of rice explained by polymorphisms in starch synthase., 2006, 4: 115–122.
[19] Commuri P D, Keeling P L. Chain-length specificities of maize starch synthase I enzyme: studies of glucan affinity and catalytic properties., 2001, 25: 475–486.
[20] Hirose T, Terao T. A comprehensive expression analysis of the starch synthase genes family in rice (L.)., 2004, 220: 9–16.
[21] Fujita N, Yoshida M, Asakura N, Ohdan T, Miyao A, Hirochika H, Nakamyra Y. Functional and characterization of starch synthase I using mutants in rice., 2006, 140: 1070–1084.
[22] 杨博文, 向珣朝, 许顺菊, 许亮, 王茜.基因与基因互作对稻米蒸煮食味品质的影响. 西北植物学报, 2017, 37: 879–884. Yang B W, Xiang X C, Xu S J, Xu L, Wang Q. Effects for interaction ofandon rice eating and cooking qualities., 2017, 37: 879–884 (in Chinese with English abstract).
[23] Li S F, Wei X T, Ren Y L, Qiu J H, Jiao G A, Guo X P, Tang S Q, Wan J M, Hu P S.encodes an-glucose transporter involved in starch synthesis and compound granule formation in rice endosperm., 2017, 7: 1–13.
[24] Nuzhdin S V, Friesen M L, Mcintyre L M. Genotype phenotype mapping in a post-GWAS world., 2012, 28: 421–426.
[25] Kong X, Zhu P, Sui Z, Bao J, Physicochemical properties of starches from diverse rice cultivars varying in apparent amylose content and gelatinization temperature combinations.,2015, 172: 433–440.
[26] 刘燕清, 强新涛, 赵春芳, 于新, 姚姝, 周丽慧, 陈涛, 赵庆勇, 朱镇, 张亚东, 王才林. 水稻淀粉合成相关基因分子标记的筛选与利用. 江苏农业学报, 2015, 31: 471–476. Liu Y Q, Qiang X T, Zhao C F, Yu X, Yao S, Zhou L H, Chen T, Zhao Q Y, Zhu Z, Zhang Y D, Wang C L. Selection and application of molecular markers for starch synthesis-related genes in rice., 2015, 31: 471–476 (in Chinese with English abstract).
[27] Rogers S O, Bandit A J. Extraction of DNA from plant tissues., 2012, 54: 979–990.
[28] 陈涛, 骆名瑞, 张亚东, 朱镇, 赵庆勇, 赵凌, 周丽慧, 姚姝, 王才林. 利用四引物扩增受阻突变体系PCR技术检测水稻低直链淀粉含量基因. 中国水稻科学, 2013, 27: 529–534. Chen T, Luo M R, Zhang Y D, Zhu Z, Zhao Q Y, Zhao L, Zhou L H, Yao S, Wang C L. Detection ofgene for low amylose content by tetra-amplification refractory mutation system PCR in rice., 2013, 27: 529–534 (in Chinese with English abstract).
[29] 蔡秀玲, 刘巧泉, 汤述翥, 顾铭洪, 王宗阳. 用于筛选直链淀粉含量为中等的籼稻品种的分子标记. 植物生理与分子生物学报, 2002, 28: 137–144. Cai X L, Liu Q Q, Tang S Z, Gu M H, Wang Z Y. Development of a molecular marker for screening the rice cultivars with intermediate amylose content insubsp.., 2002, 28: 137–144.
[30] 田志喜, 严长杰, 钱前, 严松, 谢会兰, 王芳, 徐洁芬, 刘贵富, 王永红, 刘巧泉, 汤述翥, 李家洋, 顾铭洪. 水稻淀粉合成相关基因分子标记的建立. 科学通报, 2010, 55: 2591–2601. Tian Z X, Yan C J, Qian Q, Yan S, Xie H L, Wang F, Xu J F, Liu G F, Wang Y H, Liu Q, Tang S Z, Li J Y, Gu M H. Development of gene-tagged molecular markers for starch synthesis-related genes in rice., 2010, 55: 2591–2601 (in Chinese with English abstract).
[31] 万映秀. 水稻淀粉生物合成途径中关键酶基因分子标记的开发及应用. 四川农业大学, 四川成都, 2006.Wan Y X. Development and Application of the Key Enzyme Gene Sequence-tagged Molecular Markers of Starch Biosynthesis in Rice (L.). MS Thesis of Sichuan Agricultural University, Chengdu, Sichuan, China, 2006 (in Chinese with English abstract).
[32] 中华人民共和国农业部标准米质测定方法NY147–88. 北京: 中国标准出版社, 1988. pp 4–6. Standard Rice Quality Measurement Method of Ministry of Agriculture of the People’s Republic of China NY147-88. Beijing: Standards Press of China, 1988. pp 4–6 (in Chinese).
[33] 中华人民共和国国家标准. 粮油检验大米胶稠度的测定, GB/T 22294–2008. 北京: 中国标准出版社, 2009. National Standard of the People’s Republic of China. Inspection of the Grain and Oil-determination of Rice Adhesive Strength, GB/T 22294–2008. Beijing: Standards Press of China, 2009 (in Chinese).
[34] 莫惠栋. 农业试验统计. 上海: 上海科学技术出版社, 1992. pp 151–166.Mo H D. Agricultural Experiment Statistics. Shanghai: Shanghai Scientific and Technical Publishers, 1992. pp 151–166 (in Chinese).
[35] 隋炯明, 李欣, 严松, 严长杰, 张蓉, 汤述翥, 陆驹飞, 陈宗祥, 顾铭洪. 稻米淀粉RVA谱特征与品质性状相关性研究. 中国农业科学, 2005, 38: 657–663. Sui J M, Li X, Yan S, Yan C J, Zhang R, Tang S Z, Lu J F, Chen Z X, Gu M H. Studies on the rice RVA profile characteristics and its correlation with the quality.2005, 38: 657–663 (in Chinese with English abstract).
[36] Nakamura Y. Towards a better understanding of the metabolic system for amylopectin biosynthesis in plants: rice endosperm a model tissue., 2002, 43: 718–725.
[37] Gao Z Y, Zeng D L, Cheng F M, Tian Z X, Guo L B, Su Y, Yan M X, Jiang H, Dong G J, Huang Y C, Han B, Li J Y, Qian Q., the key gene for gelatinization temperature is a modifier gene for gel consistency in rice., 2011, 53: 756–765.
[38] Bao J S. Towards understanding of the genetic and molecular basis of eating and cooking quality of rice., 2012, 57: 148–156.
[39] Tian Z X, Qian Q, Liu Q Q, Yan M, Liu X, Yan C J, Liu G, Gao Z, Tang S, Zeng D, Wang Y, Yu J, Gu M H, Li J Y. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities., 2009, 106: 21760–21765.
[40] He Y, Han Y P, Jiang L, Xu C W, Lu J F, Xu M L. Functional analysis of starch-synthesis genes in determining rice eating and cooking qualities., 2006, 18: 277–290.
[41] 吴洪恺, 梁国华, 顾燕娟, 单丽丽, 王芳, 韩月澎, 顾铭洪. 水稻淀粉合成相关基因对稻米RVA谱特征的影响. 作物学报, 2006, 32: 159–163. Wu H K, Liang G H, Gu Y J, Shan L L, Wang F, Han Y P, Gu M H. The effect of the starch-synthesizing genes on RVA profile characteristics in rice (L.), 2006, 32: 159–163 (in Chinese with English abstract).
[42] 舒庆尧, 吴殿星, 夏英武, 高明尉, McClung A. 稻米淀粉RVA谱特征与食用品质的关系. 中国农业科学, 1998, 31: 25–29.Shu Q Y, Wu D X, Xia Y W, Gao M W, McClung A. Relationship between RVA profile character and eating quality inL., 1998, 31: 25–29 (in Chinese with English abstract).
[43] 陈峰, 张正球, 张士永, 张洪瑞, 杨连群, 严长杰. 稻米淀粉的生物合成与品质改良的研究进展. 山东农业科学, 2008, (3): 20–25.Chen F, Zhang Z Q, Zhang S Y, Zhang H R, Yang L Q, Yan C J. Research progress of rice starch biosynthesis and quality improvement., 2008, (3): 20–25 (in Chinese).
[44] 姚姝, 张亚东, 刘燕清, 赵春芳, 周丽慧, 陈涛, 赵庆勇, 朱镇, Pillay B, 王才林.Wx基因背景下和基因对水稻蒸煮食味品质的影响. 中国水稻科学, 2020, 34: 217–227.Yao S, Zhang Y D, Liu Y Q, Zhao C F, Zhou L H, Chen T, Zhao Q Y, Zhu Z, Pillay B, Wang C L. Allelic effects on eating and cooking quality ofandgenes underWxbackground in rice., 2020, 34: 217–227 (in press) (in Chinese with English abstract).
[45] Asai H, Abe N, Matsushima R, Crofts N, Oitome F N, Nakamura Y, Fujita N. Deficiencies in both starch synthaseand branching enzymelead to a significant increase in amylose in-inactiverice seeds., 2014, 65: 5497–5507.
[46] Hannah L, James M. The complexities of starch biosynthesis in cereal endosperms., 2008, 19: 160–165.
[47] Zhou S, Yin L, Xue H. Functional genomics based understanding of rice endosperm development., 2013, 16: 236–246.
Effects ofandalleles and their interaction on eating and cooking quality underWxbackground of rice
YAO Shu1, ZHANG Ya-Dong1, LIU Yan-Qing1, ZHAO Chun-Fang1, ZHOU Li-Hui1, CHEN Tao1, ZHAO Qing-Yong1, ZHU Zhen1, Balakrishna Pillay2,*, and WANG Cai-Lin1,*
1Institute of Food Crops, Jiangsu Academy of Agricultural Sciences / Jiangsu High Quality Rice Research and Development Center / Nanjing Branch of China National Center for Rice Improvement, Nanjing 210014, Jiangsu, China;2School of Life Sciences, College of Agriculture, Engineering and Science at the University of KwaZulu-Natal (Westville Campus), Durban 4000, Natal, South Africa
In order to explain the variation of eating and cooking quality (ECQ) in different semi-waxy rice lines withWxallele, sixty-four semi-waxy lines withWxwere developed by crossing from Wujing 13 and Kantou 194 (Milky Princess). The polymorphism of markers between the two parents was detected in soluble starch synthesis genesandbut not in other starch synthesis related genes. Genetic effects ofandon the amylose content (AC), gel consistency (GC), gelatinization temperature (GT) and rapid visco analyzer (RVA) profile characteristics were analyzed inWxrice lines. The results showed thatandhad significant effects on the characteristic values of AC, GT, GC and RVA profile, and the interactive effects existed between the two genes.2and2alleles (2indicated that allele was derived from Wujing 13, the same as in the below) had a tendency to increase AC by 1.87% and 1.23%, respectively. This result was consistent in two years. There was no significant effect on GT forandallelic variation, whereas the GT of11(1indicated that the genes were derived from Kantou 194, the same as in the below) was significantly higher than that of22by 1.34°C. This indicated that GT was not significantly affected by single gene ofandbut was remarkably influenced by the interaction of the two genes. The GC was significant varied among rice lines with different genotypes. The2and1alleles could increase GC of 8.74 mm and 9.62 mm respectively. From the interaction of the two genes, the GC of21genotype was 10.64 mm higher than the12genotype, and was 16.95 mm higher than the22genotype. The allele2increased the peak viscosity (PKV), hot paste viscosity (HPV), cool paste viscosity (CPV) and breakdown viscosity (BDV), but decreased the consistency viscosity (CSV) and setback viscosity (SBV). However, the effect of allele2was just the opposite, which decreased the PKV, HPV, CPV, BDV and increased CSV and SBV. For the combination ofand,21genotype showed the largest values in PKV, HPV and CPV,22genotype showed the largest values in BDV and CSV, and21genotype showed the least value in SBV. These results provide a theoretical basis for improving eating and cooking quality of semi-waxyrice.
semi-waxyrice;;; eating and cooking quality; allelic effects; interaction
10.3724/SP.J.1006.2020.02006
本研究由江苏省自然科学基金项目(BK20180302), 江苏省农业科技自主创新基金项目(CX[18]1001), 江苏省重点研发计划项目(BE2018357), 江苏省农业重大新品种创制项目(PZCZ201703), 国家现代农业产业技术体系建设专项(CARS-1-62)和江苏省作物基因组学和分子育种重点实验室开放课题(PL201902)资助。
This study was supported by the Jiangsu Natural Science Foundation (BK20180302), the Jiangsu Agricultural Science and Technology Innovation Fund (CX[18]1001), the Jiangsu Key Research and Development Program (BE2018357), the Jiangsu Major New Varieties Creation Project (PZCZ201703), the Earmarked Fund of China Agriculture Research System (CARS-1-62), and the Open Project of Key Laboratory of Jiangsu Crop Genomics and Molecular Breeding (PL201902).
王才林, E-mail: clwang@jaas.ac.cn
E-mail: rice19820911@hotmail.com
2020-02-11;
2020-06-02;
2020-07-06.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20200706.1152.004.html
猜你喜欢
西南农业学报(2022年5期)2022-06-06
热带亚热带植物学报(2022年2期)2022-04-14
包装工程(2022年1期)2022-01-26
广东农业科学(2021年3期)2021-04-23
中国农业科学(2021年7期)2021-04-21
中国农业科学(2021年6期)2021-03-25
农产品加工(2020年3期)2020-03-11
红楼梦学刊(2020年2期)2020-02-06
热带农业科技(2019年1期)2019-01-14
—— 品尝员的识别能力与适口性
北方水稻(2016年4期)2016-01-09
