
嗜热甘露聚糖酶manBK基因密码子优化表达及在魔芋聚糖降解中的应用
2020-09-17 02:59卢海强谷新晰袁巧敏谈苏慧田洪涛
食品科学 2020年18期
卢海强,谷新晰,袁巧敏,谈苏慧,田洪涛
(河北农业大学食品科技学院,河北 保定 071000)
β-甘露聚糖酶(EC 3.2.1.78)是β-1,4-甘露聚糖水解酶的简称,它可以对甘露聚糖分子主链的内部随机水解β-1,4-甘露糖苷键,在食品加工领域具有广泛的应用前景[1-3]。甘露寡糖是由2~10 个甘露糖残基通过β-1,4-糖苷键聚合而成的低聚糖,属于益生元[4]。近年来,生物酶解制备甘露寡糖技术应运而生,魔芋富含葡甘露聚糖,其水溶液呈高黏度,致使无法靠增大底物浓度方式提高酶法制备甘露寡糖效率,极大限制了甘露寡糖工业化生产的发展[5-6]。研究发现葡甘露聚糖溶液黏度值随温度升高而降低,因此嗜热甘露聚糖酶在魔芋聚糖深加工中具有巨大应用潜力。
嗜热酶是一类最适温度为60 ℃以上的酶,主要从嗜热微生物中获得[7]。迄今为止,人们已成功获得了数十个嗜热甘露聚糖酶[8-9]。Thu等[10]从黑曲霉中纯化获得了甘露聚糖酶ManBK01,该酶最适反应温度为80 ℃,属于嗜热酶。Do等[11]将该基因进行了异源表达及性质测定研究。由于嗜热甘露聚糖酶ManBK巨大的应用潜力,Huang Jianwen等[12]对ManBK酶进行了晶体结构的解析,探究了结构与功能的关系。目前,关于提高该酶表达量及提升酶稳定性的相关研究鲜有报道,而上述问题的解决有利于ManBK酶在食品工业中的推广应用。
巴斯德毕赤酵母pPIC9K表达系统具有许多优点,如高细胞密度、高表达量、易于遗传操作、复杂的翻译后修饰能力以及较低的内源基因分泌表达,这使得该系统成为目前最成熟的真核表达系统之一[13-14]。为进一步提升基因的异源表达,已经在巴斯德毕赤酵母中应用了许多方法,如增加基因的拷贝数、选择合适的信号肽、高效转录启动子和细胞培养的优化[15-17],其中,将基因进行密码子优化是提高基因表达量的最有效方法[18]。
因此,本研究通过对嗜热甘露聚糖酶manBK基因密码子优化及工程菌产酶条件研究,探究提升ManBK酶表达的策略;通过对嗜热酶酶学性质的分析,探讨金属离子对酶稳定性影响及魔芋聚糖酶解产物的抗氧化性性能,旨在为ManBK酶在魔芋深加工行业中的应用提供理论依据。
1 材料与方法
1.1 材料与试剂
巴斯德毕赤酵母GS115菌株和质粒pPIC9k由本实验保存;魔芋聚糖(纯度>95%) 合肥博美生物技术有限公司;大肠杆菌感受态细胞TransI-T1 北京全式金生物技术有限公司;质粒pMDTM19-T、FastpfuDNA聚合酶、T4 DNA酶、EcoR I、NotI和BglII 宝生物工程(大连)有限公司;参照毕赤酵母表达手册制备酵母浸出粉胨葡萄糖(yeast extract peptone dextrose,YPD)培养基、最少葡萄糖(minimal dextrase agar,MD)培养基、甘油(buffered glycerol complex,BMGY)培养基、甲醇诱导(buffered methanol complex,BMMY)培养基;引物合成和核酸测序由生工生物工程(上海)股份有限公司完成。
1.2 仪器与设备
CR-GIII立式高速低温离心机 日本日立公司;聚合酶链式反应(polymerase chain reaction,PCR)核酸扩增仪 德国Biometra公司;TU-1810紫外分光光度计北京普析通用仪器有限公司;1652100电穿孔仪 美国伯乐公司。
1.3 方法
1.3.1 甘露聚糖酶manBK基因的密码子优化与合成
参照密码子使用数据库(http://www.kazusa.or.jp/codon/)中的毕赤酵母相关数据,将黑曲霉来源的manBK基因中使用频率低于0.3的密码子进行优化,同时使用RNA折叠网络服务器和mRNA翻译折叠优化的密码子以进行蛋白质二级结构预测手动校正。GC含量设定在40%~50%之间,没有局部峰,同时结合pPIC9K表达质粒的序列特征和插入位点,在目标基因的两端设计并添加了酶切位点。其中,在目标基因5’端设计EcoR I(GAATTC)酶切位点序列;此外,在目标基因3’端终止密码子(TAA)之后添加NotI(CTCGAG)酶切位点,优化后的基因由金斯瑞生物科技有限公司合成。
1.3.2 重组菌株的构建及表达及分析
以合成的质粒pUC57-manBKopt为模板,采用引物manBKopt-PF(5’-GAATTCTCTTTCGCTTCCACTTCCGG TC-3’)(EcoR I)和manBKopt-PR(5’-GCGGCCGCTTA AGCAGAACCGATAGCAGCAACG-3’)(NotI)进行基因扩增和检测。PCR条件:94 ℃预变性3 min,94 ℃变性30 s;55 ℃退火30 s;72 ℃延伸2 min,30 个循环;72 ℃延伸10 min。合成后的目标基因经测序验证无误后,采用EcoR I和NotI进行双酶切并与pPIC9K质粒酶切片段进行连接构建重组质粒pPIC9k-manBKopt;连接产物转化至大肠杆菌DH5α感受态细胞中,涂布于含氨苄抗生素的LB平板上,37 ℃过夜培养。挑取单克隆进行菌落PCR鉴定,阳性克隆送生工生物工程(上海)股份公司测序验证。将pPIC9k-manBKopt质粒进行BglII酶切线性化,并电转化至GS115感受态中,参照毕赤酵母表达手册和酶活性筛选方法对转化子进行筛选及诱导表达,离心收集发酵液,即为粗酶液,进行活力测定及十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)分析。
1.3.3 ManBK酶活力测定
参考Miller[19]的3,5-二硝基水杨酸(3,5-dinitrosalicylic acid,DNS)法,取100 μL适当稀释酶液和900 μL角豆胶(0.5%),在pH 5.0、50 mmol/L磷酸氢二钠-柠檬酸缓冲液中,80 ℃反应10 min后,加入1.5 mL DNS终止反应。对照则在加入1.5 mL DNS后,补加100 μL稀释酶液。沸水浴5 min并冷却至室温后在540 nm波长处测定吸光度。以每分钟生成1 μmol甘露糖所需的酶量定义为一个酶活力单位(U)。
1.3.4 诱导时间和温度对ManBK酶表达量的影响
将菌株1 mL菌液接种于100 mL的BMGY培养基中,在30 ℃条件下培养2d后离心收集菌体,转接至100 mL的BMMY培养基中,并在30 ℃摇床培养。收集24、48、72 h和96 h时分别取样,离心收集粗酶液,并测定酶活力,绘制菌株的产酶曲线。
将菌株1 mL菌液接种于100 mL的BMGY培养基中,在30 ℃条件下培养2 d后离心收集菌体,转接至100 mL的BMMY培养基中,分别在22、25、28、30 ℃和32 ℃条件下200 r/min摇床培养3 d,每隔24 h添加终体积分数为1%的甲醇并取样测定酶活力,测定菌株产酶的最适温度。
1.3.5 反应温度和pH值对ManBK酶活力的影响
在pH 2.0~12.0范围条件下,测定重组酶ManBK活力,以酶活力最高值为100%计算,分析该酶的pH值反应范围。最适pH值条件下,分别在30~90 ℃范围内,测定重组酶ManBK活力,以酶活力最高值为100%计算,分析其反应温度范围。
1.3.6 金属离子对ManBK酶活力及热稳定性的影响
在标准反应条件下,分别测定终浓度为1 mmol/L和5 mmol/L的金属离子条件下酶活力,探究金属离子对酶活力的影响,为重组酶的贮藏、纯化及应用提供一定的指导。
1.3.7 重组酶ManBK动力学参数测定
在酶最适反应条件下,以不同质量浓度的角豆胶(10、8、6、5、4、3、2、1 mg/mL)为底物测定重组酶ManBK活力,根据米氏方程双倒数法(Lineweaver-Burk法)求得Km和Vmax。
1.3.8 魔芋聚糖降解产物含量测定及组成分析
准确称取1 g魔芋聚糖作底物,加入到50 mL pH 5.0的50 mmol/L HAc-NaAc缓冲液,搅拌30 min,随后加入50 mL重组酶ManBK(1 100 U),70 ℃水浴90 min,100 ℃加热灭活10 min,参照Somogyi[20]方法测定魔芋聚糖降解产物中的直接还原糖(direct reducing sugar,DRS)和总还原糖(total reducing sugar,TRS),并按下式计算平均聚合度:
平均聚合度=TRS含量/DRS含量
1.3.9 魔芋聚糖降解后的抗氧化性分析
参照Brand-Williams等[21]方法测定1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除能力,具体参照试剂盒说明书的步骤进行。
1.4 数据分析及处理
每次实验重复测定3 次,利用Excel和SPSS 19.0软件对测定结果进行统计分析,数据结果采用平均值±标准误差表示。
2 结果与分析
2.1 嗜热甘露聚糖酶优化型manBKopt基因的设计和密码子优化
不同物种的密码子使用具有偏好性差异,采用毕赤酵母偏好密码子替代目标基因中的部分密码子开展重组酶的表达研究,基于巴斯德毕赤酵母有利密码子合成和构建manBKopt基因。来自黑曲霉甘露聚糖酶manBK基因(XM 006962518.1)全长为1 152 bp,编码一个含有信号肽序列(38 aa)和成熟蛋白(345 aa)的组成。去除其信号肽序列后,其成熟肽序列基因总长为1 038 bp(含终止密码子)。重新设计的目标基因manBKopt与野生型manBK-wt序列的一致性为76%。设计后的目标基因编码一条长度为345 个氨基酸残基组成的重组蛋白,与manBK-wt一致。
大量研究指出,密码子偏好性显著影响外源基因在表达宿主中的转录和翻译速率。对manBK-wt基因进行分析,结果见表1。manBKopt基因密码子的使用频率与毕赤酵母差异较大。CTC(Leu)、CGC(Arg)、GCG(Ala)和CCG(Pro)在毕赤酵母中的密码子使用频率低于30%,并且存在多个稀有密码子聚集,如Leu87(CTC)、Arg89(CGC)和Leu90(CTC)。
CTC(Leu)密码子使用偏好性差异最大,该基因中共出现13 次。CGC(Arg)密码子在毕赤酵母中的使用频率仅为10%,这些数据表明manBK-wt基因在毕赤酵母中的表达会受到一定的影响。因此,进一步对目标基因进行优化,分别将CTC(Leu)、CGC/CGT(Arg)、GCG(Ala)和CCG(Pro)替换为TTG/CTG(Leu)、AGA(Arg)、GCT/GCC(Ala)和CCT(Pro)。manBK-wt基因在优化前,其密码子适应指数(codon adaptation index,CAI)经OptimumGeneTM软件在线(http://www.genscript.com.cn/technology-support/on-line-tools)计算为0.61;经优化后,manBKoptCAI值达到了0.87。CAI值大于0.8表示该目标基因可以在毕赤酵母中高效表达。经分析,manBKopt基因中GC含量的变化范围为30%~80%,过高的GC含量会影响到基因转录及翻译速率,经优化后GC含量变化范围为40%~53%,这使得GC平均值由54%调整为47%,共有242 个碱基进行了优化。
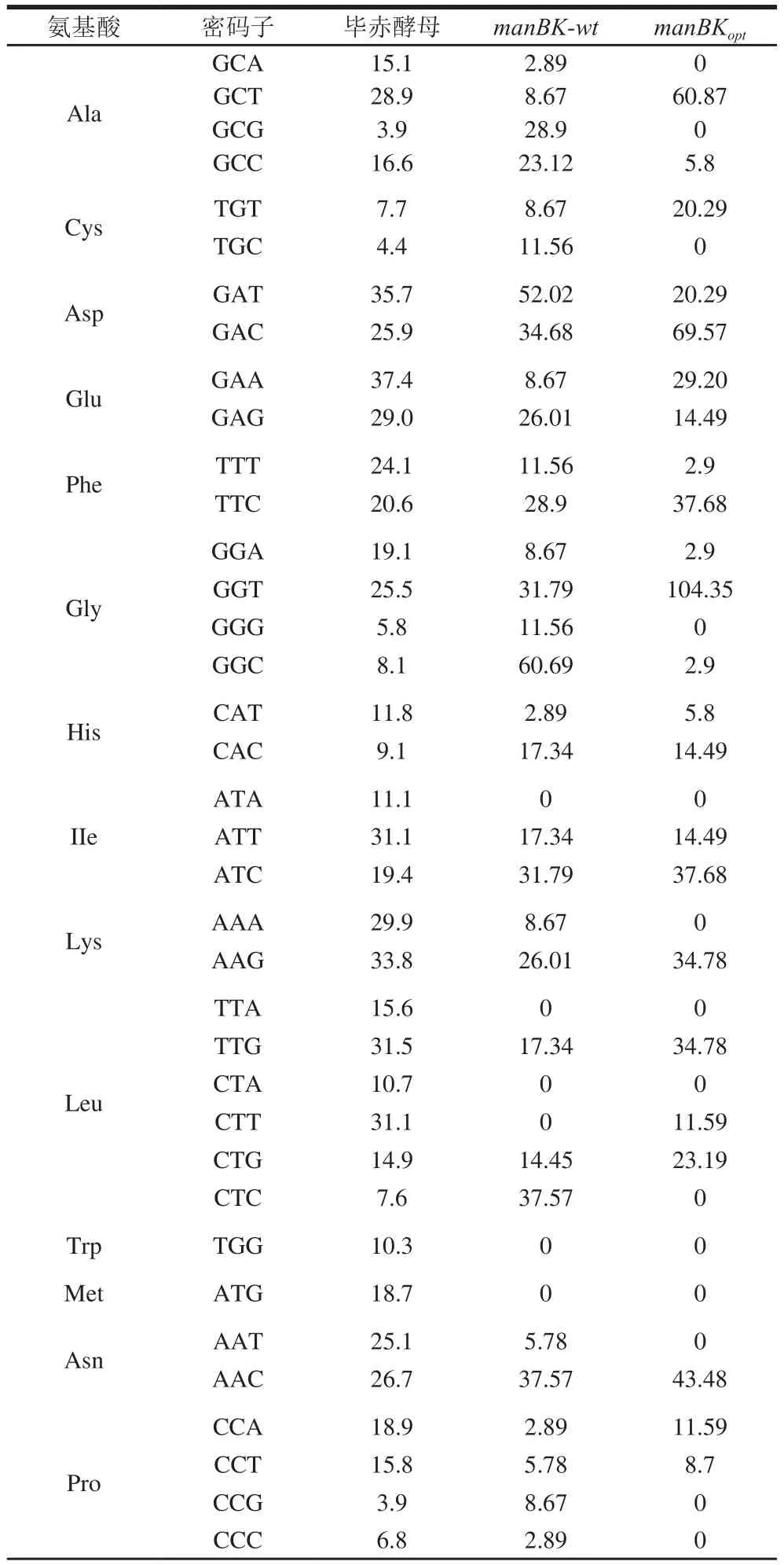
表1 野生型和优化型甘露聚糖酶manBK的密码子使用频率Table 1Codon usage frequencies for expression of wild-type and optimized manBK gene in P.pastoris%
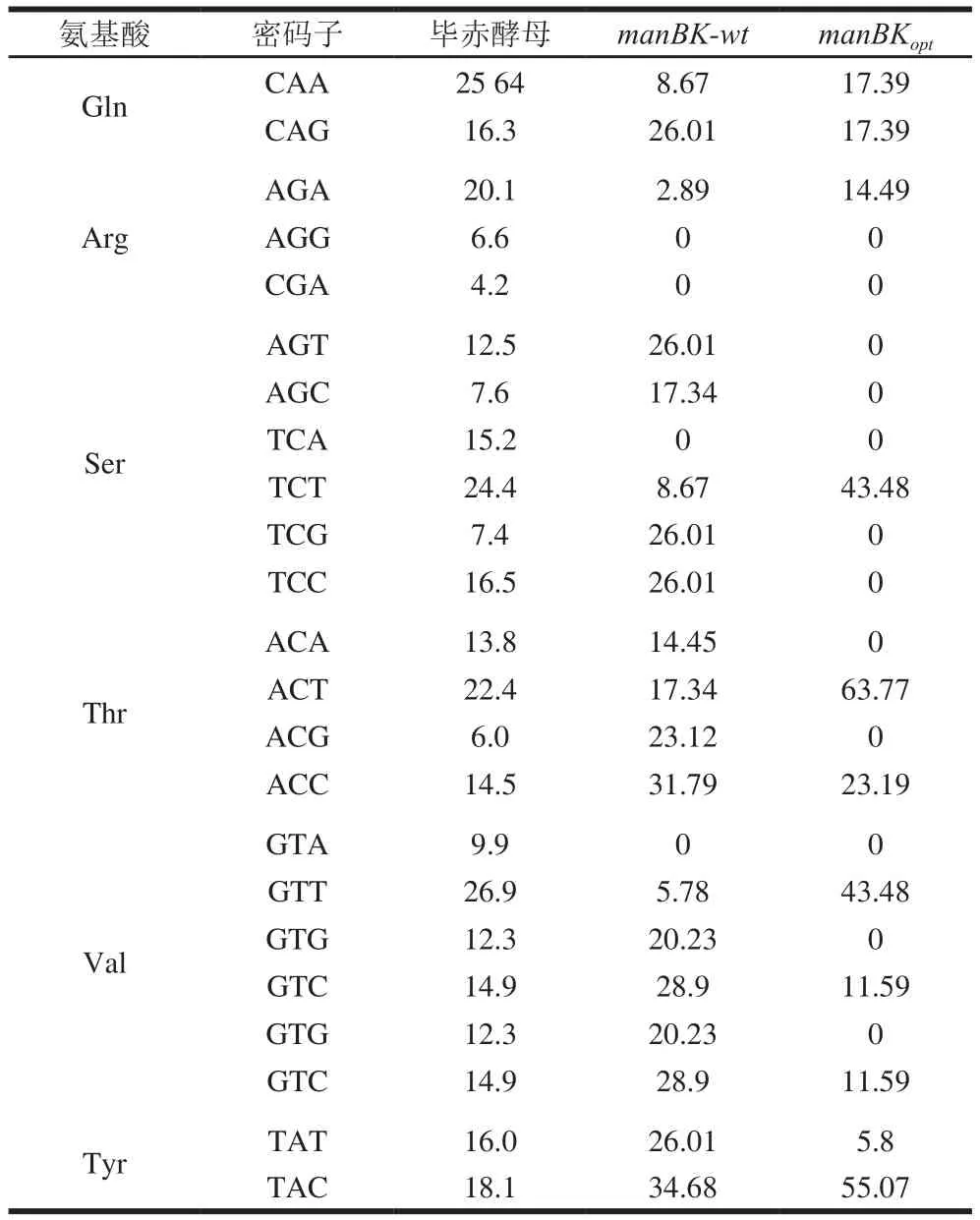
续表1
2.2 重组菌manBK基因诱导表达及产物的SDS-PAGE分析
用EcoRI和NotI消化manBKopt基因并克隆至pPIC9K中构建表达质粒。经双酶切检测及DNA测序结果分析,正确构建pPIC9k-manBKopt。经BglII限制酶线性化后,电击转入到GS115感受态细胞并在MD平板上培养,参照毕赤酵母表达手册方法,挑取48 个转化子进行阳性筛选,以角豆胶为底物,测定甘露聚糖酶的活性,共获得阳性转化子37 个。选取酶活力最高的30号转化子进行摇瓶水平诱导表达研究。将重组菌株在30 ℃条件下摇床培养并进行诱导表达,对细胞发酵液进行酶活力检测,为11.4 U/mL。
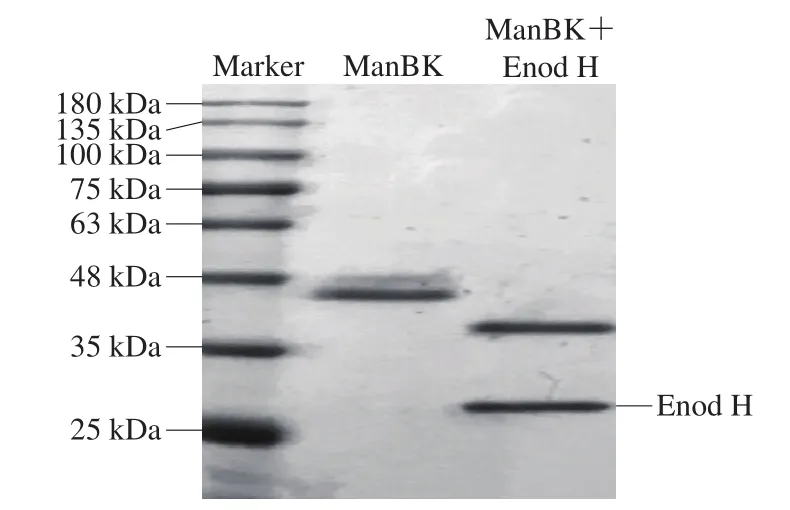
图1 ManBK酶的SDS-PAGE分析Fig.1 SDS-PAGE analysis of the ManBK enzyme
经SDS-PAGE分析得出重组酶ManBK表观分子质量为45 kDa,比理论分子质量37.7 kDa偏大,经糖基化预测软件NetNGlyc 1.0(http://www.cbs.dtu.dk/services/NetNGlyc/)分析发现该蛋白ManBK氨基酸中存在2 个潜在的N-糖基化位点(Asn-Xaa-Ser/Thr)分别是N156和N225位点。经脱糖基酶Endo H消化后,形成1 条约为38 kDa的蛋白条带(图1),结果表明ManBK酶在表达的过程中进行了N-糖基化加工修饰。
2.3 工程菌生产ManBK酶条件及酶学特性分析
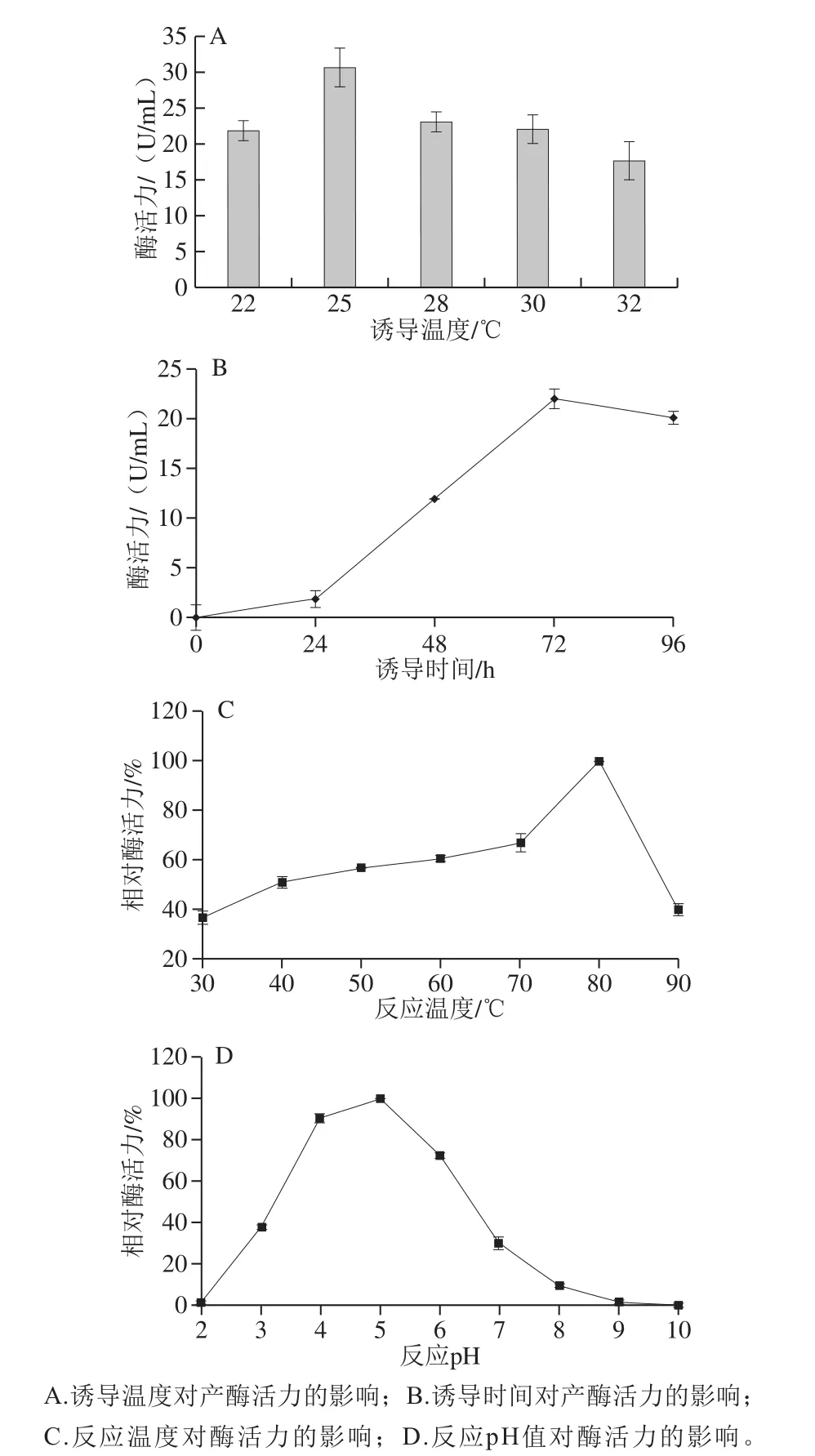
图2 ManBK酶的产酶条件和酶学性质Fig.2 Production conditions and enzymatic properties of ManBK
由于不同蛋白一级结构、蛋白稳定性及宿主蛋白酶的差异,使得诱导温度对工程菌的产酶情况有较大影响。经测定,菌株在不同的诱导温度下产酶量差异明显,该菌株在25 ℃诱导条件下产酶量最高(图2A)。诱导温度过高或过低都不利于酶的正确折叠和后修饰,在22 ℃和32 ℃诱导条件下仅能维持70%的产酶量。如图2B所示,以1%(V/V)接种量接菌至BMGY培养基中进行甲醇诱导产酶,随着诱导时间的延长,菌株的产酶量也随之增加,当诱导培养72 h后,产酶量达到最大值,其酶活力为22 U/mL,随后的24 h产酶量略有减少,这很可能是由于酶受到内源蛋白酶或其他因素造成的降解。
ManBK酶在pH 5.0、30~90 ℃条件下的酶活力如图2C所示。该酶最适反应温度为80 ℃,即使在90 ℃的高温下仍存在40%的酶活力,属于嗜热酶。该酶的反应温度范围较广,在30~90 ℃之间具有40%以上的酶活力,能够满足不同的反应温度需求。重组酶ManBK的最适pH值为5.0,属于酸性甘露聚糖酶,该酶在pH 4.0~6.0范围内酶促反应时能够维持其70%以上的酶活力(图2D)。
2.4 金属离子和化学试剂对重组酶活力的影响及动力学参数
金属离子种类和浓度对酶活力的影响差异较大,如表2所示,在低离子浓度条件下(1 mmol/L),K+、Li+、Mn2+和Cu2+对ManBK酶有不同程度的抑制作用,其中Li+的抑制最为强烈(抑制率100%),其他抑制率为20%左右,而金属离子Fe3+、Mg2+、Ca2+和Zn2+则极显著促进了重组酶ManBK活性,分别使酶活力提高了32%、23%、52%和459%。进一步对高离子浓度(5 mmol/L)下酶活性的研究结果表明,Mg2+和Ca2+依然对酶活力有较强烈的促进,分别使酶活力提高了234%和263%,而Zn2+则未表现出酶活力的促进作用。
酶的热稳定性直接影响酶使用效果和应用范围,通过测定Mg2+、Ca2+和Zn2+作用下,ManBK酶在70、85 ℃和95 ℃条件下的半衰期,探究这3 种离子对酶热稳定性的影响,结果如表2所示。经分析发现,Zn2+能延长酶的半衰期,使得ManBK酶在70、85 ℃和95 ℃条件下的半衰期分别延长了116、3.2 min和1.5 min。Mg2+和Ca2+并没有起到增强酶热稳定性效果。
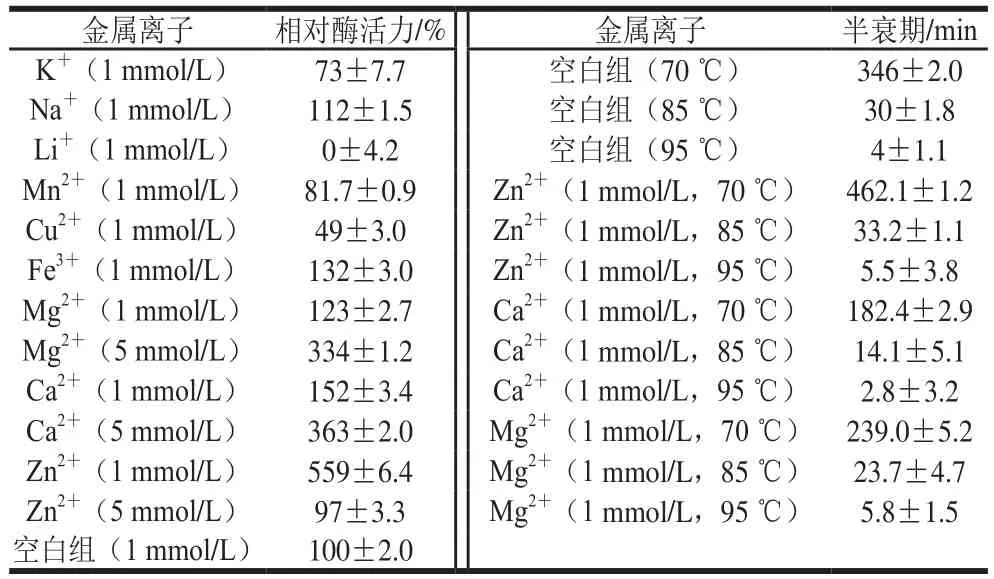
表2 金属离子对ManBK相对酶活力及热稳定性的影响Table 2 Effect of metal ions and chemical reagents on the activity and thermal stability of ManBK
按照双倒数法绘制以角豆胶为底物的ManBK酶的动力学曲线,计算得出ManBK酶的Km值为1.40 mg/mL,Vmax值为149.25 μmol/(min·mg)。
2.5 ManBK酶在魔芋聚糖降解中应用
为更好探究ManBK酶在魔芋聚糖降解中的应用特性,进行魔芋聚糖酶解过程中平均聚合度及降解产物抗氧化特性测定,结果如图3所示。
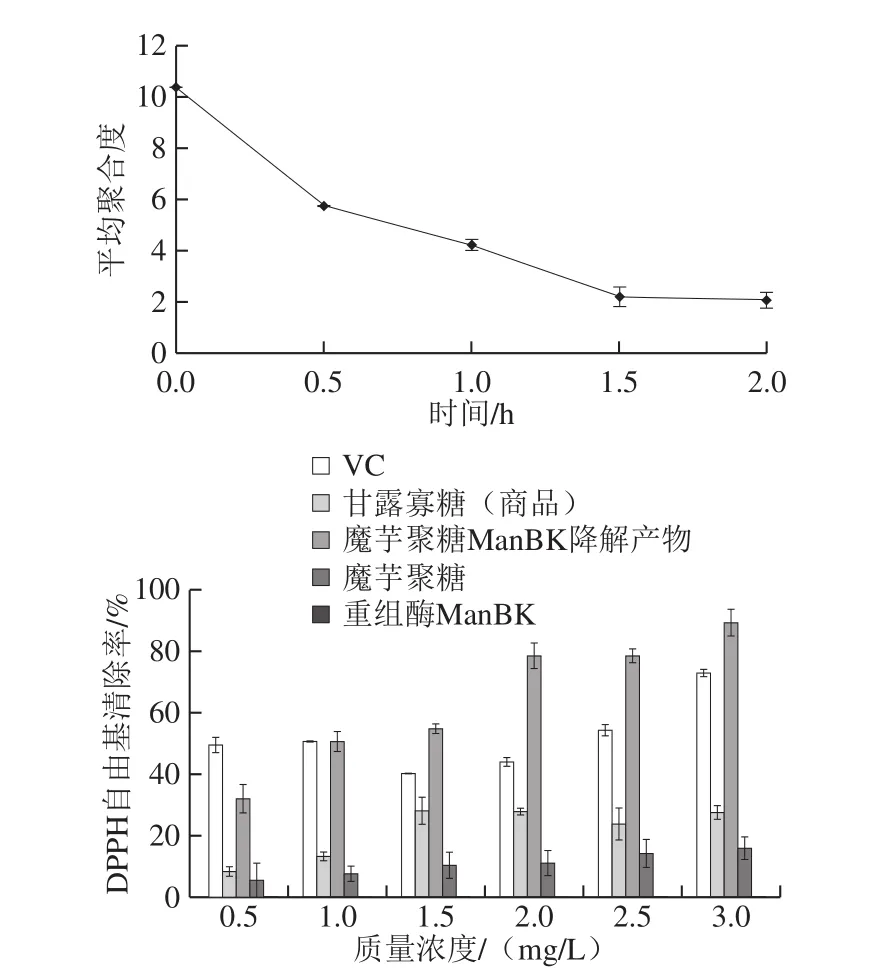
图3 魔芋聚糖降解产物平均聚合度和抗氧化活性的变化规律Fig.3 Change in polymerization degree and antioxidant activity of konjac glycan degradation products
随着酶作用时间的延长,魔芋聚糖降解产物的平均聚合度逐渐降低,表明ManBK酶将魔芋聚糖进行了有效降解,释放出大量的还原糖。当反应进行到1.5 h后,糖含量基本增加不明显,平均聚合度基本保持不变(图3),因此当酶底比为1∶1时,反应温度70 ℃,pH 5.0反应进行1.5 h,即可达到较合适的酶解时间。
魔芋聚糖经ManBK酶降解后的产物进行冷冻干燥处理,与VC、甘露寡糖(商品)和魔芋聚糖同时测定不同质量浓度下的DPPH自由基清除率,上述物质都表现出不同程度的抗氧化性,且基本上随着质量浓度的增加抗氧化性能逐渐增强,而重组酶ManBK并未表现出抗氧化性能。魔芋聚糖虽然也表现一定的抗氧化性能,但是经ManBK酶降解后制备的魔芋聚糖降解产物的抗氧化性能大幅度增强,两者抗氧化性能最高相差5.6 倍左右。商品化寡糖与ManBK酶制备的魔芋聚糖降解物的抗氧化性能比较发现,后者的抗氧化性能是前者的2~4 倍。综上,ManBK酶在魔芋聚糖深加工中具有极强的应用潜力。
3 讨 论
较高的催化反应速率、较低的杂菌污染、简化的生产工艺等优点,使得嗜热酶在在工业领域应用潜力巨大[22]。甘露聚糖酶依据氨基酸序列和结构的一致性差异可被分为GH5、GH26、GH113和GH134家族,其中GH5和GH134主要分布在真菌微生物中,而GH26和GH113家族主要来自细菌[23]。目前,嗜热甘露聚糖酶主要来自GH5家族,ManBK01就是其中之一,是唯一完成解析晶体结构的嗜热甘露聚糖酶。据报道,嗜热酶基因可以在中温的大肠杆菌和酵母菌表达宿主中按照正确构象进行折叠表达,这使得嗜热酶可以从中温表达宿主中获得,极大方便对嗜热酶的研究。嗜热甘露聚糖酶manBK在毕赤酵母中异源表达,经性质测定分析发现异源表达酶蛋白与野生型酶蛋白酶学性质一致。如何进一步提高嗜热酶ManBK的表达量、提高酶的热稳定性、探究酶在食品加工中的应用潜力成为该酶研究的新阶段内容。
manBKopt基因密码子的使用频率与毕赤酵母差异较大,其中CTC(Leu)密码子使用偏好性差异最大,CGC(Arg)密码子在毕赤酵母中的使用频率仅为10%。本研究获得的转化子酶活力为11 U/mL,远低于Do等[11]的报道(660 U/mL)。分析其原因可能是两者的表达系统的差异所致,本研究采用pPIC9K和毕赤酵母GS115表达系统,而Do等[11]采用pPICZαA和毕赤酵母X33表达系统;另外一个可能原因是manBK基因在两者宿主中的拷贝数的差异,在一定程度上基因拷贝数与基因表达量呈正相关,两者最终子宿主的拷贝数差异还需进一步研究。经SDS-PAGE分析发现,本研究所表达蛋白纯度较高,电泳纯能达到99%,能够大幅度简化对酶下游的提取工艺。
研究发现重组酶ManBK的表观分子质量远大于其理论分子质量,经分析酶蛋白分子的N-糖基化是造成分子质量增大的主要原因。产物的糖基化修饰作为真核表达系统重要的加工修饰方式,影响产物的分子质量、稳定性和催化效率等诸多属性,真核生物的糖基化对于维持蛋白的功能及稳定性方面有重大的作用[24-25],而对其他性质(如催化效率和热稳定性)的影响并不清楚,还需进一步分析。 重组酶ManBK最适pH值为5.0,这与大多数GH5家族甘露聚糖酶的性质相类似(最适pH 1.5~7.0)[26]。ManBK酶最适反应温度为80 ℃,并且该酶即使在90 ℃时依然能够维持40%的酶活力,属于典型嗜热酶,这与Do等[11]研究结果一致。
Zn2+、Ca2+和Cu2+对该酶活力性有较大促进,Do等[11]也得出了一致的结论,但是上述金属离子对甘露聚糖酶的影响效应并不适用于所有甘露聚糖酶,如Yin等[27]报道Zn2+和Cu2+对甘露聚糖酶具有强烈抑制作用,使活性降低了60%和80%。Regmi等[28]研究发现,Ca2+抑制了20%左右的酶活力。对于提升酶的热稳定性普遍采用酶的定向改造、包被处理,而采用金属离子辅助提升酶稳定性的相关研究较少[29-30]。本研究除探究金属离子对酶活力性的影响之外,经Zn2+、Ca2+和Cu2+对酶热稳定性分析发现,1 mmol/L浓度下,Zn2+对酶的热稳定性表现出了较好的促进作用,使得重组酶ManBK在70 ℃的半衰期增长116 min。本研究对采用嗜热甘露聚糖酶制备甘露寡糖进行了聚合度及抗氧化性分析,嗜热酶能够将魔芋聚糖高效降解成低聚糖,且产物表现出较好的抗氧化性能。
猜你喜欢
林产化学与工业(2022年4期)2022-09-13
中老年保健(2021年1期)2021-12-04
生物学通报(2020年11期)2020-10-22
湖南饲料(2019年5期)2019-10-15
今日农业(2019年14期)2019-09-18
发明与创新·中学生(2019年6期)2019-06-26
中成药(2018年7期)2018-08-04
农民致富之友(2017年23期)2018-01-02
中国酿造(2016年12期)2016-03-01
中国酿造(2016年12期)2016-03-01
