
杜仲EuREF1基因表达水平与橡胶积累的关系
2020-09-09 07:16:08赵德刚
林业科学研究 2020年4期
冉 昕,赵德刚,2*
(1. 贵州大学农业生物工程研究院/生命科学学院/山地植物资源保护与种质创新教育部重点实验室,贵州 贵阳 550025;2. 贵州省农业科学院贵州特色植物资源保育工程研究中心,贵州 贵阳 550006)
杜仲(Eucommia ulmoidesOliver)是我国二级珍稀保护树种,是单科单属单种的多年生落叶乔木,分布于我国的热带至暖温带地区,有27个省广泛种植[1]。杜仲既是重要的药用植物,也是优质的胶源植物,在其叶片、果皮、树皮和根皮中均有白色杜仲胶丝[2-3]。杜仲胶与天然橡胶(产自巴西橡胶树)均为聚异戊二烯,二者互为同分异构体,天然橡胶为顺式聚异戊二烯(cis-polyisoprene),杜仲胶为反式聚异戊二烯(trans-polyisoprene)。与顺式橡胶相比,反式杜仲胶的绝缘性更高,热胀冷缩系数更低,酸碱耐受性更强,并具有形状记忆功能和独特的“橡胶-塑料”二重性[4-6]。
橡胶基本单元为异戊烯基焦磷酸(IPP)。在三叶橡胶中,异戊稀基焦磷酸通过细胞中的甲瓦龙酸(MVA)途径产生,在杜仲实生叶片含胶细胞中,IPP通过MVA和甲基赤磷酸(MEP)2种途径产生[7]。IPP通过2种途径产出后,在异戊烯基焦磷酸异构酶作用下部分转化为其同分异构体二甲基丙烯基焦磷酸(DMAPP),这2种物质通过一系列异戊烯基焦磷酸合酶,包括牻牛儿基焦磷酸合酶(GPPS)、法尼基焦磷酸合酶(FPPS)和牻牛儿基牻牛儿基焦磷酸合酶(GGPPS)合成相应的牻牛儿基焦磷酸、法尼基焦磷酸、牻牛儿基牻牛儿基焦磷酸,最后在橡胶延长因子(Rubber Elongation Factor,REF)、小橡胶颗粒蛋白(SRPP)、橡胶转移酶(RT)等作用下合成长链橡胶[6]。杜仲中的反式聚异戊二烯分子,由多个反式单元的二甲基烯丙基和末端脂肪族脂肪酸的酯基或二磷酸基排列组成[8]。
在长链橡胶的生物合成过程中,橡胶延长因子(Rubber Elongation Factor,REF)发挥重要功能并受到广泛关注。1989年,Dennis等从三叶橡胶树中分离出一种分子量为14 KD的胶粒蛋白(HbREF),制备了特异性免疫球蛋白IgG抗体,或用丝氨酸蛋白胰蛋白酶(serine protease trypsin)处理该蛋白后,在体外均很大程度抑制了橡胶的生物合成,被称为橡胶延长因子[9]。之后,Attanyaka等获得了编码HbREF蛋白的cDNA序列,编码的蛋白可以与橡胶粒子紧密结合,是异戊烯基转移酶将异戊二烯单元添加到橡胶分子中不可缺少的成分[10]。此后不久,Cornish等发现,橡胶延长因子在橡胶粒子的聚合中起到控制作用[11]。目前的研究发现,除根组织外,三叶橡胶树的乳胶细胞、皮、叶中均检测到HbREF mRNA转录本,但组织间表达水平不同,通过对产胶量不同的无性系橡胶树HbREF的表达水平比较分析,发现HbREF基因表达模式与乳胶产量呈正相关[12]。本项目组通过转录数据库结合RACE技术扩增得到全长1 075 bp的序列,分析比对后命名为EuREF1[13],Wuyun等将该基因命名为EuSRPP1[14]。但EuREF对杜仲对橡胶积累及胶分子延长的关系和作用尚不明确,本论文初步分析了EuREF1基因在不同器官组织中的表达水平与橡胶积累和橡胶分子量的关系,试图探明EuREF1与胶分子量和含胶率的关系,进一步深入探究杜仲胶合成机制。
1 材料与方法
1.1 材料
实验材料取自贵州大学农业生物工程研究院试验示范基地,16年生杜仲树。选择典型样株雌、雄株各3株,从2019年4月15日开始,每15 d取样1次,至6月15日共5次。每株选择树冠中部外围当年生新梢同时期生长的叶片、茎皮和当年生果进行采摘,用自封袋封上后放于液氮中保存防止RNA降解,带回室内并保存于-80℃,用于mRNA提取和RT-PCR分析以及杜仲胶提取分析。
1.2 方法
1.2.1 杜仲胶提取与含量计算 参照Spence等的方法[15],将不同时期采摘叶片、茎皮和果皮分别用电子天平(北京丹佛仪器有限公司)称取10 g样品,电热鼓风干燥箱(上海一恒科学仪器有限公司)中120℃ 30 min,温度调至80℃,烘至恒质量,称取烘干后的样品质量,精确至小数点后3位,将方法中溶剂苯替换为石油醚(天津市致远化学试剂有限公司,沸程60~90℃)。对杜仲胶进行提取获得精胶。橡胶含量=精胶/组织干质量×100%。
1.2.2 RT-PCR分析 根据康为世纪公司的Plant RNA Kit试剂盒说明书,提取杜仲叶、茎皮、雌果中的总RNA,并利用Reverse Transcriptase MMLV试剂盒(TAKARA)反转录试剂盒反转录为cDNA。设计的EuREF1基因qPCR引物为,F:5′-CGCACATTGGTCGGAGAACTACAAC-3′;R:5′-GCAAAATAAGCAGCCGCCTCTAGTC -3′,以杜仲EuActin基因为内参基因[16],以cDNA为模板,采用RT-PCR试剂盒(Novoprotein)对EuREF1基因进行实时定量PCR分析(BIO-RAD)。反应体系Primer F/R各0.2 μL,cDNA 1 μL,Premix Mix 5 μL,ddH2O 3.6 μL,共10 μl,做3次重复。
1.2.3 杜仲胶分子量检测 杜仲胶烘干后,按照1 mg·mL-1用四氢呋喃(Merck)溶解,置于28℃摇床(Thermo)溶解3 h,用0.22 μm的有机滤头过滤溶液;装入安捷伦排阻色谱仪(1 260 Infinity II)配套小棕瓶中进行检测;流动相:四氢呋喃;样品注入量: 35 μL(1 mg·mL-1);柱温箱:40℃。得到的图谱再用GPC/SEC(gel permeation chromatography/Size-exclusion chromatography)配套软件直接处理。
2 结果与分析
2.1 杜仲不同组织的橡胶积累变化
2.1.1 杜仲叶片含胶量变化 在初春杜仲新芽长出以后,于4月15日开始至6月15日,每15 d采摘1次叶片测定含胶量。结果表明:从4月15日至6月15日,雌、雄株叶片中杜仲胶含量均不断增加;在4月15日,雌、雄株叶片含胶量分别为0.51%和0.33%,二者差异不显著;5月2日,雌、雄株叶片含胶量分别为0.84%和0.36%,雌株叶片含胶量显著高于雄株叶片;到5月15日,雌、雄株叶片含胶量分别为0.89%和0.76%,二者差异不显著;6月2日,雌、雄株叶片含胶量分别为0.95%和1.36%,雄株叶片含胶量显著高于雌株叶片;6月15日,雄株叶片含胶量增加更明显,达1.83%,雌株增长相对较少,为1.24%,二者差异显著(图1)。
2.1.2 杜仲茎皮含胶量变化 初春新生嫩茎长出后,于4月中旬开始,至6月15日,每隔15 d采摘1次,将皮剥下测定含胶量。结果表明:在4月15日至6月15日期间,雌、雄株茎皮中杜仲胶含量均不断增加;4月15日,雌、雄株茎皮含胶量分别为0.76%和0.74%,二者差异不显著;5月2日,雌、雄株茎皮含胶量分别为0.98%和0.84%,此时雌株茎皮含胶量显著高于雄株;5月15日,雌、雄株茎皮含胶量分别为1.03%和1.40%,此时雄株显著高于雌株,与叶片中含胶量相反;至6月15日时,雌、雄株含量分别为1.54%和2.01%,二者差异显著(图2)。
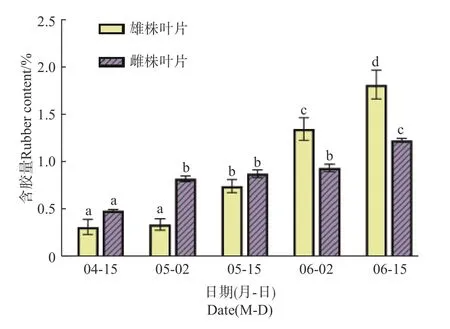
图1 雌雄株叶片含胶量积累变化Fig. 1 Accumulation of rubber content in leaf of male and female
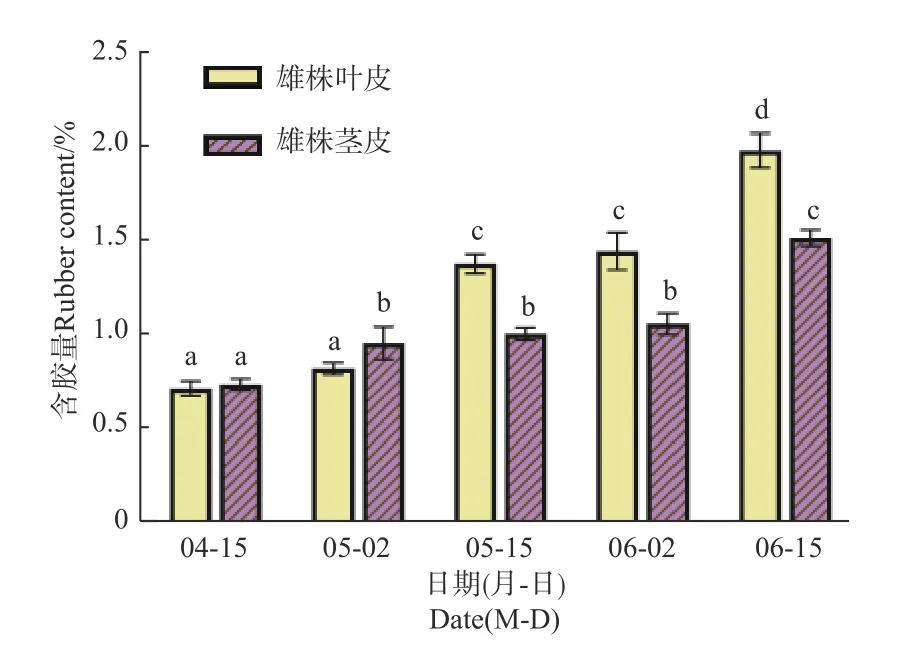
图2 雌雄株茎皮含胶量积累变化Fig. 2 Accumulation of rubber content in stem of male and female
2.2 杜仲胶分子量变化
2.2.1 叶中杜仲胶分子量变化 雌株叶片中橡胶平均分子量呈上升趋势,4月15日、5月2日、5月15日、6月2日、6月15日时平均分子量(Mw)分 别 为3.59×106、6.69×106、7.83×106、8.14×106、9.05×106Da。分子量分布随着叶片增长而变窄,4月15日至6月15日分子量分布(数均分子量/重均分子量)依次为4.14、2.73、2.64、2.48、2.01。杜仲雄株叶片中橡胶平均分子量从4月15日至6月15日同样呈现上升趋势,平均分子量(Mw)依次为7.38×106、9.94×106、11.24×106、12.06×106、14.78×106Da。雄株叶片中胶平均分子量均大于雌株。雄叶分子量分布同样随着叶片增长而变窄,4月15日至6月15日依次为5.30、4.06、3.31、3.3、2.65。说明雌、雄株叶片中胶分子量分布均随着叶片增大越来越集中。雌雄株叶片中橡胶不同分子量大小所占比列随叶片增长而发生变化,在4月15日至6月15日之间,雌雄株叶片中分子量在1.0×106Da以下所占比例分别从27.86%、17.08%降至1.10%、0.00%;分子量在1.3×107Da以上所占比例分别从3.60%、17.10%上升至20.20%、33.54%;此期间,雌、雄株叶片,分子量在1.0×106~5.0×106Da间所占比例最多(表1)。
2.2.2 茎皮中杜仲胶分子量变化 雌株茎皮中橡胶分子量从4月15日至6月15日呈上升趋势,平均分子量(Mw)分别为2.51×106、3.68×106、4.18×106、4.53×106、4.71×106Da。分子量分布从4月15至6月5日在1.95至2.73之间,说明雌株茎皮中橡胶分子量分布不随茎皮增长而变化。雄株茎皮中橡胶分子质量从4月15日至6月15日同样呈现上升趋势,平均分子量(Mw)分别为0.84×106、2.15×106、4.74×106、5.07×106、5.45×106Da。雄株茎皮中橡胶分子量分布从4月15至6月5日在3.00至3.50之间,说明雄株茎皮中橡胶分子量分布同样不随茎皮增长而变化。此期间,雌株橡胶分子量分布整体小于雄株茎皮。雌、雄株茎皮中橡胶不同分子量大小所占比列随茎皮增长而变化,在4月15日至6月15日之间,雌、雄株茎皮中分子量在1.0×106Da以下所占比例分别从44.61%、75.15%降至2.18%、15.74%;分子量在1.3×107Da以上所占比例分别从1.97%、0.93%上升至4.62%、12.76%;此期间,除4月15日时雄株茎皮中分子量小于1.0×106Da所占比例最多,其余均为分子量在1.0×106~5.0×106Da间所占比例最多(表1)。
2.2.3 果皮中杜仲胶分子量变化 果皮中橡胶分子量大小从4月15日至6月15日呈现上升趋势,平均分子量(Mw)分别为2.53×106、4.11×106、5.65×106、6.88×106、7.72×106Da。果皮中分子量分布随果增长而变窄,依次为4.88、4.49、2.61、2.33、2.25, 说明随着果增长,果皮中胶分子量分布越来越集中。果皮中橡胶不同分子量大小所占比列随果增长同样发生变化,在4月15日至6月15日之间,果皮中分子量在1.0×106Da以下所占比例从38.86%降至5.65%,分子量在1.3×107Da以上所占比例从4.76%上升至22.99%,分子量在1.0×106~5.0×106Da间所占比例最多(表2)。

表1 雌雄株叶片和茎皮橡胶分子量分布比例Table 1 Molecular weight distribution of rubber in leaf and stem of male and female %
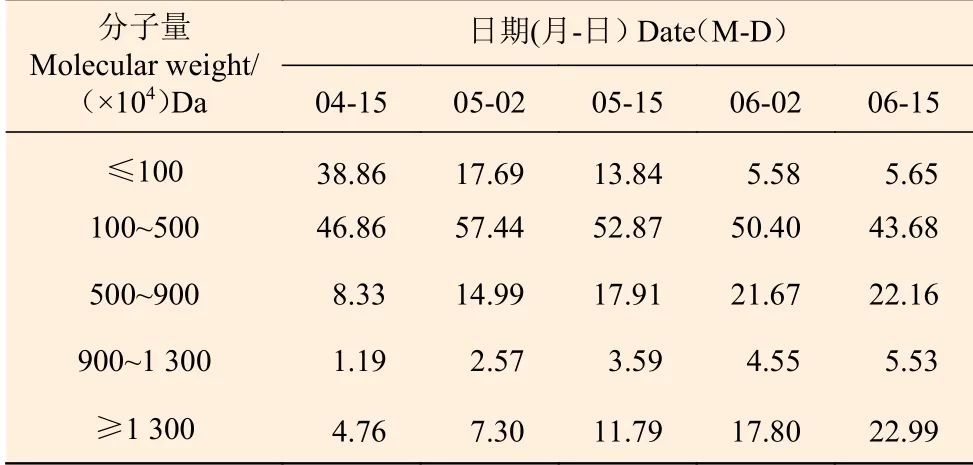
表2 果皮橡胶分子量分布比例Table 2 Molecular weight distribution of rubber in fruit%
2.3 杜仲EuREF1基因表达特征与含胶量的关系
2.3.1 叶片中EuREF1基因表达特征与含胶量的关系 提取雌、雄株叶片mRNA反转录成cDNA后,以杜仲Actin为内参基因,对4月15日至6月15日的杜仲雌雄株叶EuREF1基因相对表达量进行分析,结果表明:随着叶片增长,雌雄株叶片的EuREF1基因表达量不断增加;6月2日时,雄株叶中EuREF1表达量是4月15日的6倍;到6月15日时,雌株叶片中EuREF1表达量继续升高,而雄株叶片表达量与6月2日相比差异不显著(图3)。对雌、雄株叶片中含胶量与EuREF1表达量进行相关分析表明:雌株叶中的EuREF1基因表达量与其胶含量呈极显著正相关(P<0.01),R值为0.969 3;杜仲雄株叶中的EuREF1基因表达量与其胶含量呈显著正相关(P<0.05),R值为0.910 5(表3)。
2.3.2 茎皮中EuREF1基因表达特征与含胶量的关系 以当年发生的杜仲嫩枝分离树皮组织的mRNA反转录成cDNA,以Actin作为内参基因,对4月15日至6月16日雌雄株茎皮中的EuREF1基因表达量进行分析,结果表明:雌雄株的茎皮组织中,EuREF1基因表达量均呈上升趋势。雌株茎皮6月15日EuREF1相对表达量是4月15日的2.2倍;雄株茎皮6月15日EuREF1相对表达量是4月15日的1.7倍(图4)。对雌、雄株茎皮中含胶量与EuREF1表达量进行相关分析表明:杜仲雌株茎皮中EuREF1基因表达量与其胶含量呈显著正相关(P<0.05),R值为0.952 2;雄株茎皮中EuREF1基因表达量与其胶含量呈极显著正相关(P<0.01),R值为0.988 0(表1)。
2.4 杜仲EuREF1基因表达特征与分子量的关系
2.4.1 叶片中EuREF1基因表达特征与分子量的关系 雌、雄株叶片中分子量与EuREF1基因表达量在4月15至6月15期间均呈上升趋势,通过对二者相关性分析表明:雌、雄株叶片中平均分子量与EuREF1基因表达量均呈显著正相关(P<0.05),R值分别为0.942 8、0.898 2。
2.4.2 茎皮中EuREF1基因表达特征与分子量的关系 雌、雄株茎皮中分子量与EuREF1基因表达量在4月15至6月15期间均呈上升趋势,通过对二者相关性分析表明:雌、雄株茎皮中橡胶平均分子量与EuREF1基因表达量均呈显著正相关(P<0.05),R值分别为0.937 4、0.905 1(表4)。
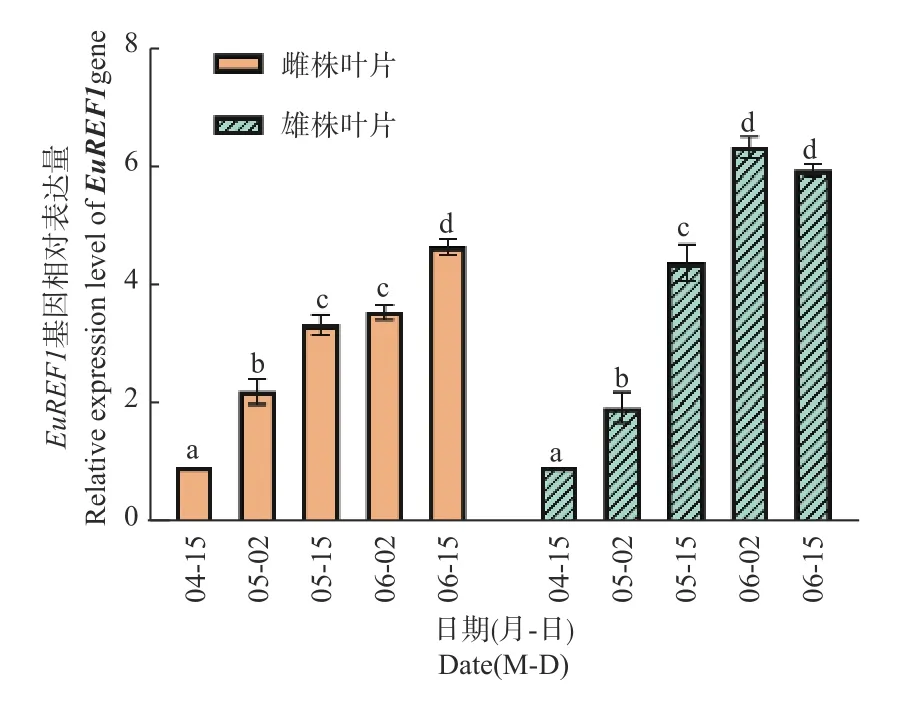
图3 叶片EuREF1基因相对表达水平Fig. 3 Relative expression level of EuREF1gene in leaf

图4 杜仲茎皮EuREF1基因相对表达水平Fig. 4 Relative expression level of EuREF1gene in stem

表3 EuREF1表达量与含胶量相关性分析Table 3 Correlation analysis of EuREF1 expression and rubber content

表4 EuREF1表达量与橡胶分子量相关性分析Table 4 Correlation analysis between EuREF1 expression level and molecular weight of rubber
3 讨论
3.1 杜仲橡胶积累规律
杜仲所产的反式橡胶具有独特的“橡胶-塑料”二重性[4]。杜仲的茎皮、叶片以及果皮中均含有杜仲胶,但随着季节的不同,不同器官含胶量也不同。雌、雄株叶片或茎皮中含胶量均随着器官生长而增加;在杜仲生长初期,叶片和茎皮中橡胶积累较缓慢,到5-6月时,橡胶积累量快速增加,与莫俊等[17]结果一致。研究发现,含有杜仲橡胶的胶颗粒粒径随器官成熟而增大[18-19],含胶细胞内的细胞核与细胞器随着器官的发育而解体,并被橡胶颗粒充满[20]。推测5月后环境条件适宜杜仲生长发育,加速胶颗粒中橡胶合成相关基因转录和翻译,使得橡胶积累量增加。研究发现,雌、雄株茎皮中的含胶量均大于叶片,与杜红岩结果一致[21]。杜仲叶中小胶颗粒、中等胶颗粒、大胶颗粒所占比例较平均,而皮中小胶颗粒所占比例为98.38%[19],推测雌、雄株不同器官中含胶量虽都有增加,但增加量和总含量有差异可能与器官中所含胶颗粒大小有关。
3.2 杜仲胶分子量变化规律
杜仲胶是三倍半萜烯醇的一个高分子量同系物,主要由反式聚异戊二烯单体构成,对杜仲雌、雄株不同时期叶片、茎皮与果皮中橡胶分子量进行测定,在4月15至6月15期间,不同器官分子量均随时间的延长呈上升趋势,但增加量有差异。Bamba等[22]在杜仲不同组织中检测到低分子量聚异戊二烯的分布有组织特异性,认为不同组织间聚异戊二烯伸长的调节机制可能不同。对于雌雄株不同器官中不同时期各分子量大小所占比列而言,随着器官生长发育,小分子量橡胶占比逐步降低,而大分子量橡胶占比逐步增加,说明各器官组织中橡胶分子量随着器官生长而增大。雌、雄株叶片、茎皮和果皮中,分子量在1×106~5×106KD间所占比例几乎为最多,异戊二烯分子量为68.12 KD,说明长度在1.5万至7.4万个异戊二烯单体的橡胶最多。
3.3 EuREF1基因表达特征与分子量及胶含量的关系
对杜仲雌、雄株叶片和茎皮中EuREF1表达量与分子量大小和含胶量进行比较,EuREF1在雌、雄株不同时期的叶片和茎皮中均有表达,且表达量均呈上升趋势,与分子量大小变化规律和含胶量增长规律呈正相关关系,叶片和茎皮中高分子量橡胶占比也逐步增加。
4 结论
通过对不同时期杜仲雌、雄株各器官EuREF1基因表达量与含胶量、分子量比较分析,得出以下结论:EuREF1表达量与含胶量增长规律呈正相关关系,推测EuREF1基因产物与杜仲胶积累有密切关系;EuREF1表达量和分子量大小变化规律呈正相关关系,推测EuREF1基因与橡胶链的延伸有关系,但仍需要直接证据。
猜你喜欢
安徽农业科学(2024年5期)2024-04-20 11:38:27
西北园艺·果树(2024年1期)2024-04-01 09:10:14
磷肥与复肥(2022年5期)2022-06-18 07:25:54
光源与照明(2021年1期)2021-09-26 16:44:16
中小企业管理与科技·上旬刊(2020年5期)2020-07-14 15:31:41
园林科技(2020年1期)2020-01-18 05:13:00
铁道学报(2019年10期)2019-11-08 08:21:14
现代农业科技(2018年17期)2018-10-13 12:31:24
中国医药科学(2015年21期)2016-01-20 20:50:39
食品与生物技术学报(2014年11期)2014-12-25 02:28:58
