
中年女性,右下肢无力3年,左下肢无力5个月
——远端型遗传性运动神经病
2020-09-02 09:56:18周美鸿朱敏洪道俊
中国神经精神疾病杂志 2020年6期
周美鸿朱敏洪道俊
1 资料
患者女性,37岁,因“右下肢无力3年,左下肢无力5个月”于2019年8月3日就诊我院。
患者3年前顺产1女婴后,逐步出现右下肢无力,开始时仅有右足上抬无力,随后发展为行走时足尖先着地,后演变为右下肢上楼不利,上抬受限,并出现右下肢远端部位肌肉萎缩,左侧下肢无明显无力不适,无肢体皮疹、疼痛、麻木、烧灼感或刺痛等不适。5个月前患者顺产二胎男孩后,逐步出现左下肢无力,症状发展类似于3年前右侧下肢受累情况。下肢无力进行性加重,行走需人搀扶,坐起下床困难。自发病以来,精神萎靡,食欲不振,睡眠差,无发热,无头痛,无肌肉痉挛或跳动,上肢运动无明显异常,无肢体麻木,无体位性头晕或黑朦,无视物重影及视物旋转,自产后体质量下降约5 kg,无大小便功能障碍。既往史无其他特殊疾病,自幼按计划进行免疫接种,无特殊药物史,也无特殊毒物接触史,家族内无特殊遗传病史,也无类似家系患者。
体格检查:体温36.4℃,心率78次/min,呼吸18次/min,血压110 mmHg/80 mmHg,颈静脉无怒张,心肺腹未见异常,双下肢无水肿。神志清楚,言语清晰流利,摇摆跨域步态,记忆力、计算力、定向力、判断力及理解力等高级皮层功能检查未见明显异常。双眼视力粗测正常。双侧瞳孔等大正圆,直径约3 mm,直接及间接对光反射灵敏。双眼睑闭合力可,双侧眼球向各个方向运动正常,未及复视和眼震。角膜反射正常,双侧面部痛温觉无减退。双侧颞肌、咀嚼肌对称、有力,双侧额纹、鼻唇沟对称。悬雍垂居中,双侧软腭上抬有力,咽反射存在。双侧转颈、耸肩对称有力。伸舌居中,无舌肌萎缩及震颤。双上肢无明显肌萎缩,双足部肌肉、双侧胫前肌、腓肠肌萎缩,右侧较左侧著,双上肢肌张力正常,双下肢肌张力减低(右侧著)。双上肢肌力5级,右上肢轻瘫试验阳性(右侧小指外展),左下肢远端踝背屈1级,踝跖伸2级,左下肢近端4-级,右下肢远端踝背屈0级,踝跖伸1级,右下肢近端3级。双侧指鼻试验正常,跟膝胫试验无法配合检查。双侧肢体及躯干痛温觉、位置觉、振动觉及运动觉正常。双侧肱二头肌腱反射、肱三头肌腱反射、桡骨膜反射活跃,双侧膝腱反射减低,跟腱反射未引出。双侧Hoffmann′s 征阳性,双侧 Rossolimo′s 征阳性,掌颌反射和吸吮反射阴性, 双侧 Babinski′s征和 Chaddock′s征均阴性。脑膜刺激征阴性。自主神经功能检查正常。
辅助检查:血常规、大小便常规、肝肾功能、凝血功能、血糖、血脂、电解质、同型半胱氨酸、甲状腺功能、铜蓝蛋白、C反应蛋白、抗CCP抗体、抗核抗体、风湿抗体未见明显异常。肌酸激酶189 U/L(正常值43~165 U/L),叶酸2.50 ng/mL(4.2~19.8 ng/mL),维生素 B12>2000 pg/mL。血尿λ和κ轻链水平正常,血清免疫固定电泳IgM/A/G未见异常条带,血清蛋白电泳无异常,莱姆抗体阴性。脑脊液初压100 mmH2O,无色清亮透明,无凝块,潘氏试验阴性,白细胞总数 20×106/L(正常值:0~8×106/L),脑脊液蛋白、糖、氯均正常,脑脑脊液革兰染色、改良抗酸染色、墨汁染色、脱落细胞学均阴性。血和脑脊液神经节苷脂抗体谱及副肿瘤综合征抗体谱均阴性。头颅和颈胸腰脊髓核磁成像未见明显异常。运动神经传导速度显示左侧腓总神经未记录到动作电位,右侧腓总神经波幅显著降低(踝点0.2 mv),双侧胫神经运动传导波幅降低(4.1~5.8 mv),右尺神经波幅轻度降低,其他上肢神经传导速度和波幅均正常。双侧正中神经和尺神经inching检测均未见传导阻滞。感觉神经传导速度检查示所检感觉神经传导未见异常改变。针刺肌电图示左侧胫前肌未见运动单位电位,静息时见多量自发电位;右腓肠肌偶见运动单位电位;左侧股四头肌运动神经电位时限增宽,电压增高,大力收缩募集电位呈单项相,静息时见少量自发电位及束颤电位;左第一骨间肌静息时见多量自发电位,大力收缩募集电位呈单混相;胸锁乳突肌未见异常。H反射显示双侧胫神经潜伏期在正常范围,右侧波幅低。超声检查显示双侧腓肠肌、比目鱼肌结构紊乱,血流不规则丰富,有斑片状的高回声,右侧相对明显。双侧腓肠神经外膜增厚,神经干下段增粗,筛网状结构消失,硬度稍增大。双下肢大腿肌肉核磁显示双侧股四头肌和腘绳肌弥漫散点状脂肪浸润,伴轻微水肿样改变 (图1A~C);双下肢小腿肌肉核磁显示右侧小腿较左侧小腿萎缩,双侧胫前肌和腓肠肌出现斑片状脂肪浸润,伴随水肿样改变(图 1D~F)。
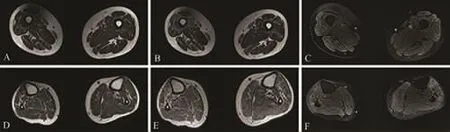
图1 患者下肢磁共振图像 双下肢大腿肌肉磁共振显示双侧股四头肌和腘绳肌弥漫散点状脂肪浸润(A:T1加权,B:T2加权),伴轻微水肿样改变(C:T2压脂)。双下肢小腿肌肉磁共振显示右侧小腿较左侧小腿萎缩(D:T1加权),双侧胫前肌和腓肠肌出现斑片状脂肪浸润(E:T2加权),伴随水肿样改变(F:T2压脂)。
病理检查:为明确诊断行左侧腓肠神经及腓肠肌活检。腓肠神经活检的苏木精-伊红(HE)染色显示神经外衣结构正常,血管壁未见坏死和炎细胞浸润,神经束衣内结构正常,未见明显水肿和吞噬现象(图2A)。髓鞘碱性蛋白(myelin basic protein,MBP)免疫组化染色显示有髓神经纤维密度大致正常,个别神经纤维的髓鞘膨大(图2B)。神经丝蛋白(neurofilament,NF)免疫组化显示神经纤维密度基本正常(图2C)。甲苯胺蓝半薄染色显示神经束内神经轴索密度基本正常,分布均匀,可见个别有髓神经纤维的髓鞘变薄,个别大有髓神经纤维直径肥大(图2D)。腓肠肌肉活检HE染色显示肌束内的小角状萎缩肌纤维成组分布,少数肌纤维肥大 (图3A),NADH染色显示萎缩肌纤维深染,少数肌纤维内出现靶或靶样肌纤维(图3B),SDH/COX复染再次证实靶样肌纤维存在,未见COX阴性肌纤维(图3C),ATP酶碱性(PH值10.4)染色显示两性肌纤维群组化改变,萎缩肌纤维累及两型(图3D)。
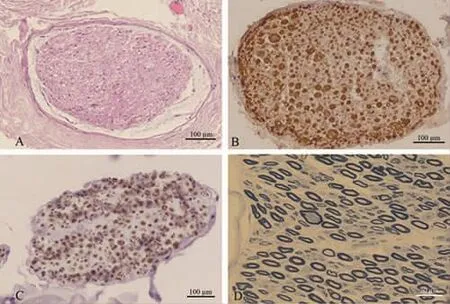
图2 左侧腓肠神经活检 HE染色显示神经外衣结构正常,血管壁未见坏死和炎细胞浸润,神经束衣内结构正常,未见明显水肿和吞噬现象(A)。髓鞘碱性蛋白免疫组化染色显示有髓神经纤维密度大致正常,个别神经纤维的髓鞘膨大(B)。神经丝蛋白免疫组化显示神经纤维密度基本正常(C)。甲苯胺蓝半薄染色显示神经束内神经轴索密度基本正常,可见个别有髓神经纤维的髓鞘变薄,个别大有髓神经纤维直径肥大(D)。

图3 腓肠肌肉活检 HE染色显示肌束内的小角状萎缩肌纤维成组分布,少数肌纤维肥大(A),NADH染色显示萎缩肌纤维深染,少数肌纤维内出现靶或靶样肌纤维(B),SDH/COX复染再次证实靶样肌纤维存在(C),ATP酶碱性(PH值10.4)染色显示两性肌纤维群组化改变,萎缩肌纤维累及两型(D)。
基因检测:患者血DNA标本二代测序显示丙氨酰tRNA合成酶基因(AARS)存在一处新的杂合突变c.2177+1G>A。一代Sanger测序进一步证实了该位点的存在,且患者父母并无该位点突变。该位点位于15号内含子的剪接受体第一位核苷酸,具有高度的保守性,根据核酸剪切软件分析可能影响导致15号外显子的剪切异常,进而导致AARS蛋白的移码截断突变。根据临床-病理-基因检测结果,该患者诊断为AARS基因突变导致的远端型遗传性运动神经病(distal hereditary motor neuropathy,dHMN)。
2 讨论
远端型遗传性运动神经病是一种少见的遗传性神经系统疾病,1966年由Nelson和Amick首次报告[1]。目前国内尚无其发病率的准确统计,在英国的发病率为2.14/100000,爱尔兰的发病率为1/100000[2-3]。尽管dHMN没有进入第一批121种罕见病目录中,但是有着更高发病率的其姊妹病运动神经元病 (motor neuron disease,MND)和Charcot-Marie-Tooth病(CMT)都属于经典的罕见病。
dHMN是一组隐袭性发展的,仅累及运动神经或者显著累及运动神经的长度依赖性周围神经病[4]。其临床具有显著的临床异质性,发病年龄跨度较大,多数患者在青少年期发病,但是成年期发病患者并不少见,本例患者即为成年期发病患者。dHMN临床上主要表现为肢体远端的肌无力和肌萎缩,多数患者下肢远端首先受累,也有部分患者首发上肢远端肌无力,但球部肌肉受累症状在dHMN患者中罕见。多数dHMN患者均表现为对称性的临床症状,但文献也有个别不对称发病患者的报告[5]。本例患者首先变现为单侧下肢远端肌无力和萎缩症状,3年后才发展到对侧下肢,进一步提示dHMN可以存在显著的不对称性起病,临床诊疗中不应过分强调不对称症状作为排外遗传性疾病的权重。
本例患者dHMN临床症状和妊娠生产存在密切的时间轴关系,然而目前没有文献报道dHMN和妊娠之间的关系,故其是否有因果关系,或者两者之间存在怎样的联系尚不清楚。已有研究显示妊娠或生产后女性可以出现多种周围神经的病变,其中个别CMT患者可以表现为病情加重进展,其机制可能和激素水平、免疫调节、营养变化、应急事件等相关[6]。在我们神经肌肉病患者库中,也观察到个别遗传性神经肌病患者在妊娠过程中出现显著加重的案例。本例患者文化素养较高,其明确表示病情启动和发展分别开始于两次妊娠产后,通过实验室检查基本排外了免疫性、代谢性、损伤性等因素,但是脑脊液细胞数轻度升高的意义不明,是否提示有炎性因素参与病程发展有待商榷。总之,妊娠和dHMN之间的关系值得进一步探究,对于该病的预防和治疗可能存在价值。
电生理检查对dHMN诊断至为重要[7]。首先,神经传导速度测定提示运动神经轴索受累,感觉神经不受累或者非常轻微受累,如本例患者就为选择性运动神经受累,纯感觉神经腓肠神经的病理改变轻微而无特异性。其次,神经电生理检查提示为长度依赖性受累,H反射或F波等反应神经根或前角细胞的指标无明显异常,本例患者双侧腓总神经受累明显,和临床上显著的足下垂可以对应,而无明显前角细胞受累证据,而且肌肉活检病理改变也符合神经干受累表现,而缺乏前角细胞受累的病理改变。再次,神经电生理一定要排外多灶性运动神经病的可能,本例患者Inching法没有发现神经走形上存在节段性传导阻滞,且GM1抗体阴性[8]。最后,dHMN患者的肌电图检查和ALS患者的肌电图都提示广泛神经源性损害,通常超过三个节段,因此临床上不能单纯分析肌电图的结果就盲目下ALS的诊断,而应该结合神经电生理其他检查以及临床表现,注意dHMN的鉴别诊断[9]。
1993年Harding依据临床表型及遗传模式等相关特征将其分为7个类型(dHMN I-VII),该疾病分类仍然广泛地在临床中运用[10]。然而,随着临床表型和基因表型的深入研究,逐步认识到Harding分型存在一定的不足,目前倾向于将dHMN分为单纯性dHMN、dHMN伴轻微感觉受累型(即motor predominant CMT2)、dHMN伴其他神经系统受累(即dHMN plus)三个亚型[11]。dHMN伴其他神经系统受累亚型患者常合并小脑共济失调 (spinocerebellar ataxia,SCA)、痉挛性截瘫(spastic paraplegia,SPG)、肌萎缩侧索硬化(amyotrophic lateral sclerosis,ALS),甚至远端型肌病,在临床上有时很难与脊髓前角病变导致的运动神经元病相鉴别[12-14]。例如HSPB1基因突变既可以导致经典的单纯型dHMN,也可以导致CMT2,甚至可以导致ALS,因此在一些dHMN患者选择性运动神经受累可能只是疾病的一部分表型,或者是疾病过程的某一个阶段[15]。从整个病变机制分析,dHMN 和 CMT2、ALS、SPG、SCA 都属于轴索病(axonapathy)的范畴[16]。本例患者临床上表现为下肢远端显著受累的肌无力、肌萎缩,肌电图提示下肢运动神经波幅显著下降伴传导速度下降,而感觉神经不受累,临床电生理符合远端型运动神经病的诊断,但是其临床查体存在上肢的锥体束受累体征,提示AARS基因突变导致的dHMN也可以存在皮质脊髓束的轴索受累症状[4],此类患者病程后期如果进一步出现皮质脑干束的受累,以及肢体近端受累,可能与经典的ALS鉴别困难。
dHMN是一组单基因遗传病,可以是常染色体显性遗传,也可以是常染色体隐性遗传,甚至有X连锁隐形遗传,目前仅有大概15%的dHMN患者发现了致病基因[17-18]。到目前为止已发现30多种dHMN致病基因,这些蛋白的异常可能影响RNA合成、细胞骨架形成、神经轴突运输及线粒体功能等,进而主要导致神经轴索功能障碍[19]。已有研究显示HSPB1基因突变的dHMN最常见的原因,HSPB1是一种分子伴侣蛋白,类似的蛋白还包括HSPB3、HSPB8、FBXO38、DNAJB2等,这些基因突变都可以导致dHMN,因而又被总称为分子伴侣蛋白病相关性dhMN[15]。除此之外,tRNA合成酶相关基因突变导致的dHMN是第二常见的致病基因种类,目前已知 AARS、GARS、HARS、WARS都可以导致dHMN[20-21]。其中AARS负责丙氨酰tRNA的合成,本例患者在保守的剪切位点存在一个错义突变,将导致整个蛋白质的截断,最终导致蛋白功能缺失或者降解,因而该突变为致病突变的可能性较大。AARS首先在中国dHMN家系中被发现证实[21],目前研究发现AARS显性遗传突变可以导致CTM2N或dHMN,而隐性遗传突变可以导致早发性癫痫脑病伴髓鞘缺陷。相信依托国内优势的患者资源,中国学者对dHMN相关基因的克隆研究将会做出更多贡献。
目前对于dHMN的治疗主要包括物理治疗、职业治疗、矫形器和辅助设备,以及支持性护理[22]。随着我们对dHMN的分子病理生理学理解的深入,新的生物标志物和治疗策略正在进行积极的临床试验。尽管大剂量抗坏血酸的临床研究没有显示对疾病严重程度以及预后有积极影响,但是更多的有希望的药物仍在路上[14]。期待进一步完善我国dHMN患者常见的致病基因的筛查,增加对dHMN症状前诊断和产前诊断率,并为治疗方法的优化提供帮助。
3 点评
随着遗传学和免疫学的进展,谱系病的概念在当下神经系统疾病的诊断中运用越来越广泛。就本病例代表的dHMN而言,很好的诠释了谱系病的概念,具体表现在如下两方面。
第一方面,经典的遗传性周围神经病称为Charcot-Marie-Tooth病,又称为遗传性运动感觉神经病(hereditary motor sensory neuropathy,HMSN),是指运动神经和感觉神经都受累的一大组周围神经病,大概包括80多种类型[23],但是在临床中也存在一些选择性累及运动神经的遗传性周围神经病,就是本文所讲述的dHMN,大概有30多种类型[1]。此外还有一组遗传性感觉自主神经病(hereditary sensory autonomic neuropathy,HSAN),其临床主要为选择性累及感觉神经和自主神经病,大概也有30多种类型[24]。HMSN、dHMN、HSAN这三大类遗传性周围神经病的临床和基因之间有一定的重叠性,特别是HMSN轴索型 (即CMT2)的运动为主型和dHMN之间存在很大的重叠[25]。此外,部分遗传性周围神经病患者,在病程早起阶段表现为dHMN或HSMAN,但是随着病情发展为HMSN。因此,尽管根据临床特点可以将遗传性周围神经病分为HMSN、dHMN、HSAN,但是在临床实践中要重视该类患者亚临床的损害特点和长期随访。
第二方面,dHMN的主要致病机制影响了运动神经轴索的功能,导致轴索性周围神经病。部分维持轴索功能的蛋白,对运动神经元同等重要,因此可能同时导致运动神经元病变。锥体束本质上也是一束轴索纤维构成,因此可以造成周围神经轴索功能障碍的蛋白,理论上也可能导致锥体束的轴索功能。基于上述原理,同一种基因突变既可以导致dHMN,也可以导致运动神经元病、痉挛性截瘫、脊髓小脑共济失调等整个大有髓轴索行程的范围,因而目前临床上提出了轴索病(axonapathy)的概念[16]。
猜你喜欢
中国现代医生(2022年19期)2022-11-04 10:13:29
昆明医科大学学报(2022年4期)2022-05-23 13:04:50
基层中医药(2021年8期)2021-11-02 06:24:54
健康之家(2021年19期)2021-05-23 09:10:44
成都信息工程大学学报(2020年5期)2020-07-29 08:50:16
中国生物医学工程学报(2019年6期)2019-07-16 07:53:00
中国医药指南(2016年12期)2016-07-09 08:16:03
中国卫生标准管理(2015年4期)2016-01-14 05:16:45
山东医药(2015年16期)2016-01-12 00:40:07
护士进修杂志(2015年17期)2015-03-19 06:34:06
