
MiR-92b-3p通过靶向RAD21增强非小细胞肺癌对顺铂的化学敏感性
2020-08-08 03:30潘有光傅文凡莫益俊赵健陈岗东
实用医学杂志 2020年14期
潘有光 傅文凡 莫益俊 赵健 陈岗东
1广州医科大学附属第三医院(广州510150);2广州医科大学附属肿瘤医院(广州510095);3南方医科大学深圳医院(广东深圳518000)
肺癌是人类最常见的恶性肿瘤之一,其中非小细胞肺癌(non-small cell lung cancer,NSCLC)约占所有原发性肺癌病例的80%[1]。在我国城市人口中,肺癌的发病率及病死率均居首位,且大多数NSCLC 患者确诊时已是晚期,失去手术机会。因此,化疗药物如顺铂(cisplatin,DDP)等广泛应用于一线治疗晚期NSCLC 患者[2-3]。然而,多数患者在经过化疗后会对DDP 产生耐药[4-7],使治疗效果减弱甚至失效,导致预后不良[8]。因此,迫切需要了解NSCLC 产生DDP 耐药的分子机制,寻找新的治疗策略以克服DDP 耐药。
microRNAs(miRNAs)是一类单链、小分子非编码RNA(长度为19~25 个核苷酸),通过与靶基因的互补结合,在转录后负向调控靶基因蛋白的表达水平[9]。已经证明,大约60%的细胞蛋白编码基因是由miRNA 调控的[10]。同时,miRNA 也参与了NSCLC 的化疗耐药[11-12]。研究[13]表明,过表达miR-146a 可促进细胞凋亡,增加NSCLC 对DDP 的敏感性。miR-181c 在NSCLC 组织中上调,促进了肺癌细胞对DDP 的耐药[14]。然而,miR-92b-3p 是否在NSCLC 中介导耐药尚未明确。本研究拟阐明miR-92b-3p 在NSCLC 细胞中对DDP 耐药的生物学功能,初步发现了miR-92b-3p 可能通过调控RAD21基因的表达来介导细胞对DDP的药物敏感性。
1 材料与方法
1.1 主要试剂顺铂(DDP,江苏豪森药业);Cell Counting Kit-8(日本化学同仁研究所);Trizol(美国invitrogen 公司);miRNA 逆转录试剂盒(美国ABI公司);microRNA 荧光定量PCR 试剂盒(美国ABI公司);miRNAmimics(上海吉玛公司);miRNAinhibitors(美国Dharmacon 公司);双荧光素酶检测试剂盒(美国promega 公司);一抗anti-RAD21(美国cell signaling techonology 公司);一抗anti-β-actin(美国ProteintechGroup 公司);二抗goatanti-mouse(美国invitrogen 公司);cDNA 合成试剂盒及PCR 试剂盒(大连宝生物有限公司)。
1.2 细胞培养及体外顺铂耐药株的建立NSCLC亲本细胞A549由中山大学医学部提供。以A549细胞系为亲本,采用顺铂富集法逐步诱导形成A549/DDP(顺铂耐药)细胞。最终诱导药物浓度达到6 μg/mL。当细胞在6 μg/mL的顺铂中存活2个月且活性正常时,证实为耐顺铂细胞株,命名为A549/DDP。A549/DDP 培养基含有2 μg/mL 的顺铂以维持细胞的耐药性。取对数生长期细胞为实验对象。
1.3 miRNA表达的微阵列检测用总RNA纯化试剂盒分离A549 细胞和A549/DDP 细胞的总RNA。使用服务提供商(LCSciences)进行微阵列分析。使用激光扫描仪采集杂交图像,然后用Array-Pro图像分析软件进行数字化处理,比较A549 和A549/DDP的表达谱,用红色(高表达)到绿色(低表达)表示两组表达差异的比值,计算t检验的P值,P<0.01的信号为差异检测信号。
1.4 细胞转染及药物敏感性分析miR-92b-3p mimic(miR-92b-3p)和miRNA 阴性对照(miR-NC)、miR-92b-3p inhibitor(anti-miR-92b-3p)、inhibitor 阴性对照(anti-miR-NC)分别转染至细胞株中,转染步骤按操作指南进行。24 h 后换液。然后用DDP(浓度梯度为0、3、6、9、12、15 μg/mL)继续培养48 h。加10%CCK8 培养液培养1 h 后,酶标仪测定吸收值。计算不同药物浓度下的半数抑制浓度(half inhibition concentration,IC50)值。
1.5 实时荧光定量PCR(Real time-PCR)用Trizol试剂(Invitrogen,USA)从培养细胞中提取总RNA。使用逆转录试剂盒从总RNA 中获得cDNA,合成的cDNA 在Real-timePCR 仪 检 测miRNA 和mRNA的表达水平。
1.6 荧光素酶报告系统检测利用pGL3-MCScontrol质粒进行基因3′UTR区域荧光报告基因的克隆构建。利用Primer Premier 5.0 分别设计RAD21的3′UTR 引物,克隆到报告基因载体pGL3-MCScontrol 中,构建成双荧光报告系统pGL3-RAD21-3′UTR-WT,pGL3-RAD21-3′UTR-MUT。A549 接种24 h 后,共转染miRNA mimics 或NC 对照,继续培养48 h,双荧光素酶试剂盒进行检测。
1.7 Western Blot 检测转染后,收集各组细胞,裂解液裂解细胞后收集蛋白,测量蛋白浓度后,通过SDS-PAGE凝胶电泳分离后转移至PVDF膜上,封闭后2.5%脱脂奶粉的PBST 及一定比例的RAD21及β-actin抗体结合,4 ℃过夜。室温下二抗孵育1 h,最后进行曝光显影。
1.8 统计学方法统计分析采用GraphPad Prism 5 Demo 以及SPSS 18.0 软件,实验数据以表示,组间差异用one-wayANOVA 检验,以P<0.05 为差异具有统计学意义。所有实验重复3 次。
2 结果
2.1 miRNA 基因芯片筛选靶基因本研究使用miRNA 芯片分析比较了A549/DDP 及A549 细胞株的miRNA表达谱,发现有33个miRNA在A549/DDP细胞与A549 细胞表达上差异存在统计学意义(图1A)。图1B 列出了差异表达水平相对较高的miRNAs。值得注意的是,几个差异表达明显的miRNA(miR-17、miR-20a、miR-20b、miR-92a、miR-92b 及miR-106a)包含相同的种子序列,在A549/DDP 细胞中显著下调,通过前期的一些预实验后,最终筛选出miR-92b-3p作为进一步研究的对象。
2.2 miR-92b-3p在A549/DDP细胞系中下调qRTPCR 结果显示,与A549 相比,A549/DDP 中miR-92b-3p 的表达显著下调(P<0.01,图2A)。此外,药物敏感性试验表明,与A549/DDP 相比,A549 的耐药性明显下降。这些结果表明,异常表达的miR-92b-3p 可能与NSCLC 细胞对DDP 的耐药有关(图2B、C)。
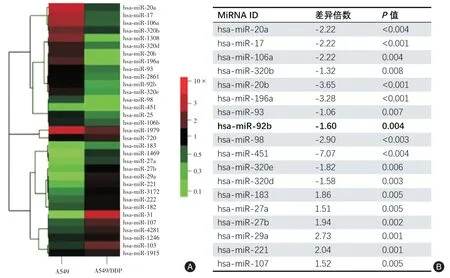
图1 miRNA 基因芯片筛选靶基因Fig.1 Screening target genes by miRNA microarray
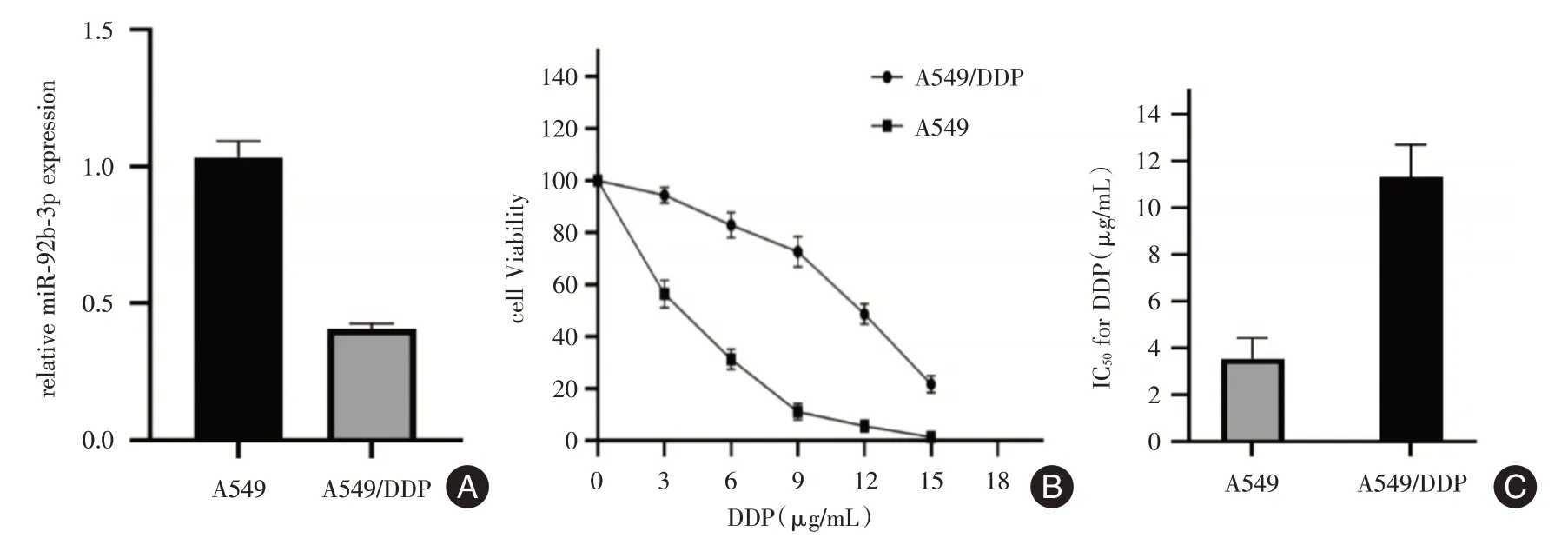
图2 A549 及A549/DDP 中的miR-92b-3p 相对表达量及DDP 浓度梯度下各自的IC50Fig.2 The relative expression of mir-92b-3p in A549 and A549/DDP and the respective IC50 under the concentration gradient of DDP
2.3 miR-92b-3p 可上调A549/DDP 细胞对DDP 的敏感性将A549/DDP 细胞株同时转染miR-92b-3p mimics 及miR-NC,CCK-8 结果显示,转染了miR-92b-3p mimics 的A549/DDP 细胞株IC50值显著降低,对顺铂敏感性增加。见图3。
2.4 miR-92b-3p 的下调可降低A549 细胞对DDP的敏感性将anti-miR-92b-3p 及anti-miR-NC 分别转染亲本A549 细胞,qRT-PCR 验证miR-92b-3p 在A549 细胞中的下调(P<0.01,图4A)。CCK-8 结果显示:下调A549 细胞株中miR-92b-3p 将显著提升其对顺铂的耐药性,见图4B、4C。
2.5 RAD21 是A549/DDP 细胞中miR-92b-3p 的直接靶点本研究通过生物信息学网站对miR-92b-3p 的靶基因进行了预测和鉴定。RAD21 被预测为miR-92b-3p 的结合位点(图5A)。双荧光素酶报告基因测定显示:与共转染阴性对照相比,报告基因载体pGL3-RAD21-3′UTR 共转染miR-92b-3p mimics的荧光素酶活性显著降低;RAD21是miR-92b-3p 的一个新的靶基因(图5B)。
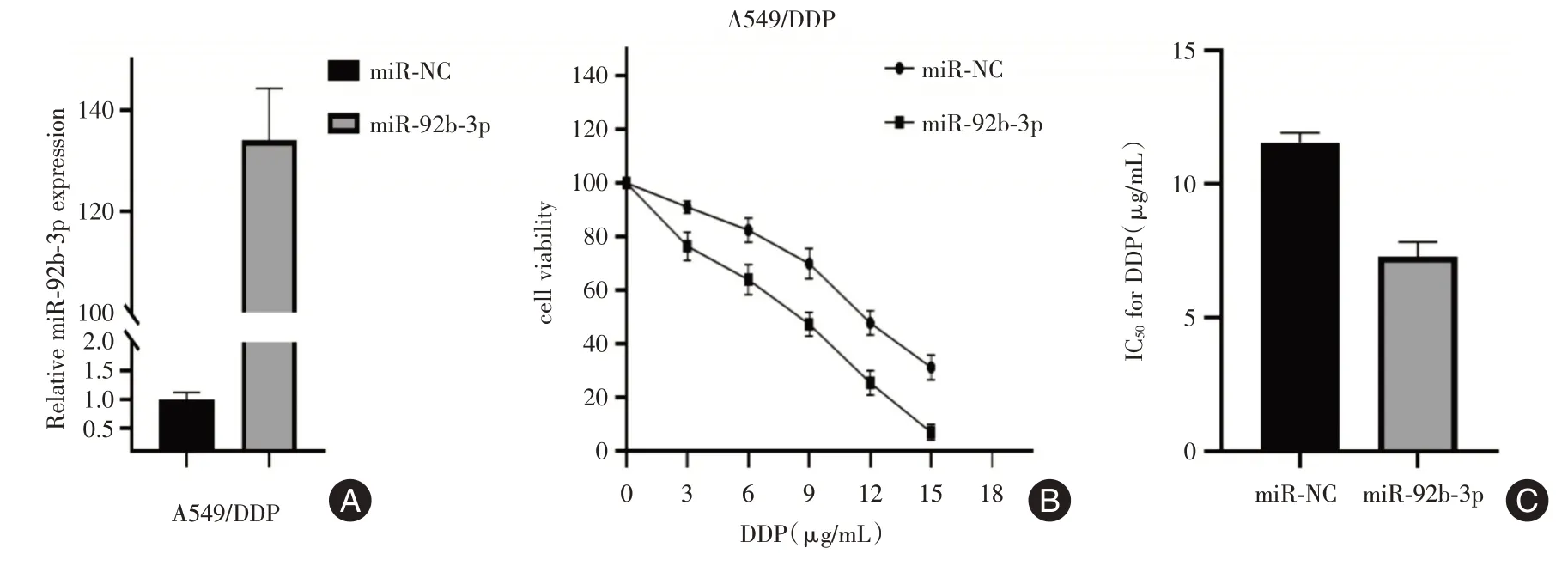
图3 miR-92b-3p mimics 的转染效率及转染后实验组与对照组对顺铂的敏感性Fig.3 The transfection efficiency of miR-92b-3p mimics and the sensitivity of the experimental group and the control group to cisplatin after transfection
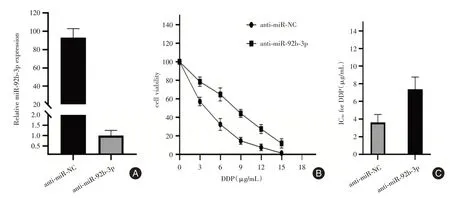
图4 miR-92b-3p inhibitor 转染效率及实验组与对照组对顺铂的敏感性Fig.4 The transfection efficiency of miR-92b-3p inhibitor and the sensitivity of the experimental group and the control group to cisplatin after transfection
2.6 Western Blot在A549/DDP 细胞中,与miRNC组相比,过表达miR-92b-3p可抑制RAD21mRNA和蛋白质含量。反之,如果抑制A549细胞中的miR-92b-3p,可上调RAD21mRNA 和蛋白质含量。因此,这些发现表明,RAD21 可能是miR-92b-3p 的直接靶点(图6)。
3 讨论
化疗耐药性仍然是目前治疗NSCLC 遇到的瓶颈。然而,NSCLC 治疗中耐药的分子机制在很大程度上仍有待于阐明。近年来,人们认为多种分子通路参与了耐药性的形成,包括异常的药物摄取和外排、DNA 修复功能的增强、凋亡通路的中断、细胞解毒的激活等[15]。此外,有证据表明,基因和表观遗传变化,如扩增、易位、突变和组蛋白翻译后修饰也有助于化疗期间的耐药性[16-17]。最近,有研究报道,miRNAs 在获得性DDP 耐药方面发挥了关键作用,这为开发新的基于miRNA 的治疗药物克服NSCLC 的耐药带来了希望[18]。MA 等[19]发现,在前列腺癌细胞株PC-3M-2B4 中,用顺铂处理过的PC-3M-2B4 细胞株,其miR-92b-3p 的表达是下调的,而GSTM3 的表达却是上调的,表明miR-92b-3p 可以通过负向调控GSTM3 的表达而改变前列腺癌对顺铂的敏感性。
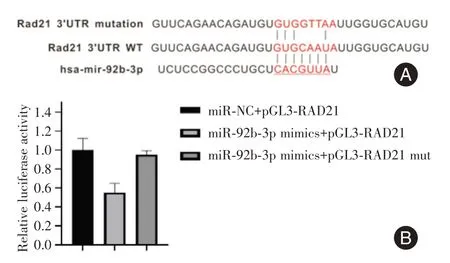
图5 miR-92b-3p 与RAD21 的3′UTR 的潜在结合点及荧光素酶活性Fig.5 Potential binding site and luciferase activity of 3′UTR of mir-92b-3p and RAD21
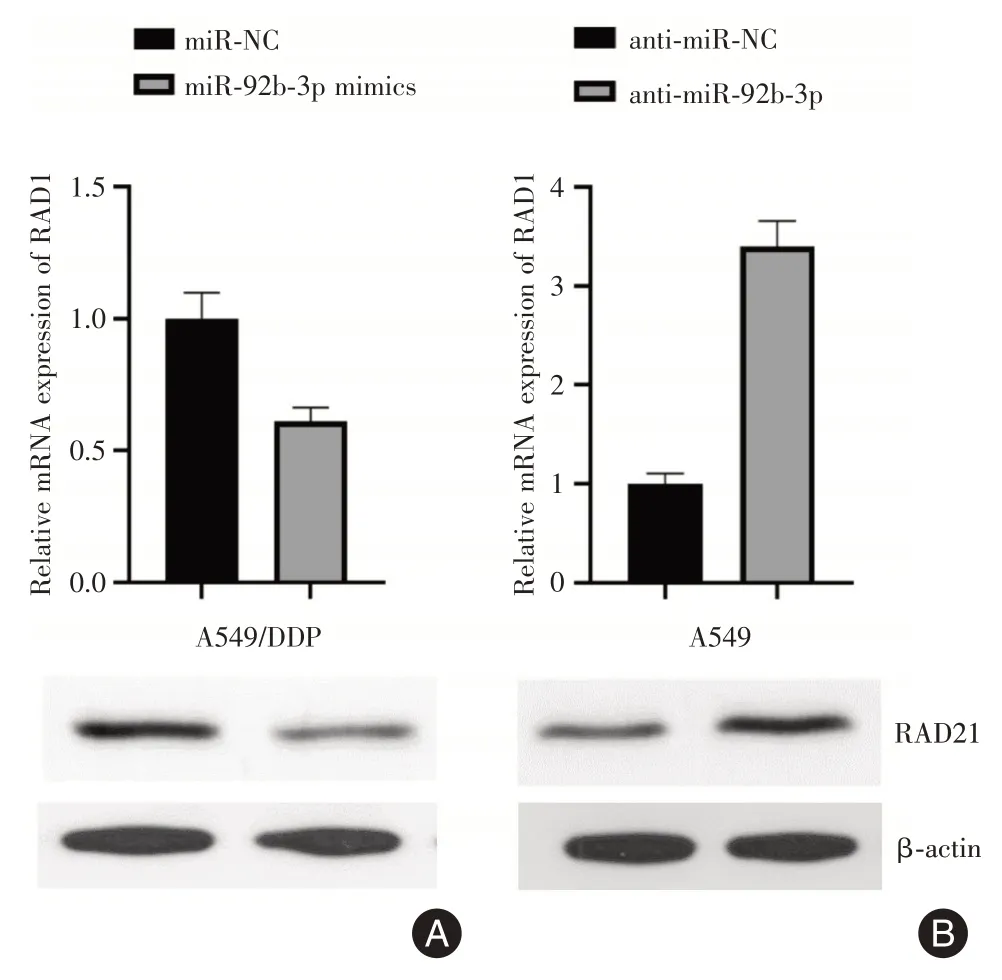
图6 miR-92b-3p 对A549 及A549/DDP 中RDA21 mRNA 及蛋白的影响Fig.6 Effects of miR-92b-3p on the expression of RDA21 mRNA and protein in A549 and A549/DDP
本研究中,qPCR 结果显示miR-92b-3p在A549/DDP 细胞株中的表达明显下调,表明miR-92b-3p可能参与NSCLC 的耐药。通过TargetScan、Pictar和miRBaseTargets 等生物信息学网站预测及双荧光素酶报告基因验证miR-92b-3p 的下游靶基因是RAD21。RAD21基因位于人染色体8q24.11上,其编码一种人类同源的非洲酒酵母RAD21 蛋白,是内聚蛋白复合体的关键核心成分[20],主要功能是在有丝分裂的中后期保证染色体的正确分离[21]。同时,它还参与了染色体的同源重组及DNA 双链修复等过程[22]。研究发现,RAD21 基因的敲除可显著增强乳腺癌细胞对5-FU、环磷酰胺和依托泊苷的敏感性[23],说明其可修复化疗药物介导的DNA 损伤。miR-92b-3p 的低表达可增强G2 期RAD21 介导的DNA 修复,减少S 期顺铂诱导的DNA 损伤,导致顺铂耐药。在本研究中,荧光素酶报告基因实验表明,在DDP 耐药的NSCLC 细胞中,miR-92b-3p 可以与RAD21 的3′-UTR 结合。过表达miR-92b-3p 可抑制DDP 耐药NSCLC 细胞株中RAD21 的mRNA 和蛋白表达。这些数据表明,miR-92b-3p 通过直接靶向NSCLC 细胞中的RAD21,高表达的RAD21 可以帮助细胞在S 期有更多的机会修复顺铂诱导的DNA 损伤,使肿瘤细胞化疗期间保持顺铂耐药性。
本研究结果显示,与DDP 敏感的亲代A549 细胞相比,A549/DDP 细胞中miR-92b-3p 的表达明显下调。恢复miR-92b-3p 可通过靶向RAD21 增加A549/DDP 细胞对DDP 的敏感性,这些发现可能为克服DDP 耐药NSCLC 的治疗提供一个有希望的生物标志物。然而必须承认,耐药是一个复杂的调控体系,本研究只是其中的冰山一角,并且验证药物单一,本课题组后续实验将进一步阐明下游的信号通路及上游与长非编码RNA 的调控关系,并在临床大样本中验证,从而为肺癌耐药机制提供新的实验依据。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
昆明医科大学学报(2021年8期)2021-08-13
西安交通大学学报(医学版)(2021年3期)2021-05-15
广东蚕业(2021年1期)2021-03-18
临床超声医学杂志(2020年12期)2020-12-20
中西医结合肝病杂志(2020年2期)2020-10-27
世界华人消化杂志(2020年2期)2020-02-14
江苏农业科学(2019年23期)2019-03-03
医学信息(2016年29期)2016-11-28
热带农业科学(2015年9期)2015-10-14
