
BnaBZR1和BnaPIF4基因调控甘蓝型油菜弱光光效的机制
2020-07-21 08:29官春云
作物学报 2020年8期
冯 韬 谭 晖 官 梅 官春云,*
和基因调控甘蓝型油菜弱光光效的机制
冯 韬1,2谭 晖1,2官 梅2官春云2,*
1遵义师范学院生物与农业科技学院, 贵州遵义 563000;2湖南农业大学农学院/ 国家油料改良中心湖南分中心, 湖南长沙 410128
甘蓝型油菜品系XY881和XY883是湘油15辐照诱变后连续自交筛选的2个种子含油量、光合效率和弱光敏感性等有明显差别的子代品系。分别从XY881和XY883中克隆了芸薹素唑抗性因子1 (brassinazole-resistant 1,) 和光敏色素互作因子4 (phytochrome interacting factor 4,)基因并进行了序列结构、表达和功能分析。结果表明, XY883的和基因存在结构变异, 引起表达和调控模式的差异。XY883中的启动子具有124 bp的富含A / T的插入序列, 且XY883具有比XY881高的表达, 并且在弱光和2,4-表油菜素内酯(2,4-BL)诱导下具有较少的表达变化。XY883中的5'-UTR区域存在可变剪接, 形成长度分别为424 bp (U01)、239 bp (U02)和332 bp (U03)的3种5'-UTR, 在弱光和2,4-BL诱导下, XY883中3种可变剪接的转录产物的变化不一致。将的3个5'-UTR与CDS分别组合转化拟南芥后其表达在转录水平无明显差异, 但蛋白翻译存在明显差异, 表明的5'-UTR变异影响其翻译过程。转基因拟南芥出现株高增加、叶片狭长且光合作用下降的表型, 共转化能减弱造成的光合作用下降; 转和基因对油菜的影响与拟南芥相似, 但表型不如拟南芥明显, 表明BnaPIF4是油菜光合作用的负调控因子, 而BnaBZR1可对BnaPIF4的光合负调控产生拮抗; 这与XY881和XY883中两基因表达调控模式及其光合表型相吻合。
甘蓝型油菜;;; 可变剪接; 插入突变
植物中转录因子PIF4和BZR1互作是植物油菜素内酯信号与光信号互作的核心节点[1], 同时二者互作也是介导植物对弱光照等非生物胁迫响应的节点[2]。油菜素内酯通过一系列信号转导调节BZR1和BZR2 (BES1)的表达和磷酸化[3], 并通过磷酸化等方式串联与之互作的其他因子调控下游基因表达, 调控植物生长, 最后形成完整的反馈回路平衡植物内源油菜素内酯的合成[4]。拟南芥和水稻中BZR1/2以单体或复合体形式与PIF4互作, 介导下游的基因转录调控[5]。植物中PIF4蛋白在光照条件下稳定性较弱, 极易发生泛素化降解, 因此植物PIF4在转录水平的调控也是相关信号互作中重要的调节方式。
甘蓝型油菜是世界范围内最重要的油料作物之一[6], 但其具有极为复杂的异源二倍体基因组[7], 不同甘蓝型油菜对低光等非生物胁迫的抗性有极大的多态性[8], 但油菜相关和的表达调控与功能均不清楚。前期研究发现, 从甘蓝型油菜湘油15中克隆基因发现其A03染色体上的同源拷贝相较测序品种中双11号存在着内含子突变[9], 这表明在甘蓝型油菜中基因可能存在着更多的突变。甘蓝型油菜品系XY881和XY883由相同亲本湘油15经辐照诱变后连续自交筛选而得, 其中XY881基本保持了湘油15的原有特征, XY883则表现为更高的光合效率, 更高的种子含油量和油酸含量, 但同时种子含油量的稳定性较低、对低光寡照胁迫更为敏感。在XY881和XY883中进行和基因克隆、序列特征、表达和功能分析, 对探明油菜中和的表达调控规律及其在低光照胁迫中扮演的角色具有重要意义, 同时也将为阐明甘蓝型油菜对低光照胁迫响应存在差异的分子机制提供一些新的理论支持。
本文克隆了两品系甘蓝型油菜中和基因并进行了完整的比较分析, 确定了甘蓝型油菜品系XY883中和基因存在的可变剪接和启动子插入突变, 并通过酵母杂交、表达分析和植物遗传转化等方式确认了XY883]中和基因突变对基因表达和蛋白翻译的影响, 并初步确定了和在拟南芥和油菜中的功能。
1 材料与方法
1.1 植物材料与培养
甘蓝型油菜品种(系)湘油15、XY881、XY883以及野生型拟南芥由国家油料改良中心(湖南)提供。分别于湖南农业大学耘园试验基地和室内植物光照培养箱中(KBW240, BINDER, 美国) (16 h/8 h光周期)种植受试材料, 以常规水肥管理。在植物培养箱中设置光源100 µmol m–2s–1模拟强光照, 20 µmol m–2s–1模拟弱光照, 试验基地中以遮光网遮光模拟弱光胁迫。
1.2 取样和测试
分别于甘蓝型油菜品系XY881和XY883种子萌发后45、120、200和220 d对根、茎、叶、花、角果及完成遗传转化后的甘蓝型油菜和T2代拟南芥叶片随机取样, 以液氮速冻后储存于-80℃冰箱, 用于RNA和蛋白提取。对经不同处理的甘蓝型油菜品系XY881和XY883完全成熟的种子随机取样, 以索氏抽提测试含油量, 以气相色谱法[10]测试脂肪酸组成。
1.3 DNA提取、RNA提取与cDNA合成
使用CTAB-PVP提取液从甘蓝型油菜叶片中提取基因组DNA。使用TRIzol RNA Extraction Kit (TransGen Biotech Co., Ltd.)从甘蓝型油菜和拟南芥样品中提取总RNA。使用Easy Script First-Strand cDNA Synthesis SuperMix Kit (TransGen Biotech Co., Ltd.)进行第1链cDNA合成。
1.4 基因克隆
从BRAD数据库(http://brassicadb.org/brad/)获取甘蓝型油菜和基因及转录本序列并进行序列染色体步移分析。截取和起始密码子上游1 kb序列为启动子克隆区域, 根据序列特征设计用于和启动子及全长mRNA克隆的引物(表1), 以PrimeSTAR GXL DNA聚合酶(TaKaRa, 日本)进行PCR。
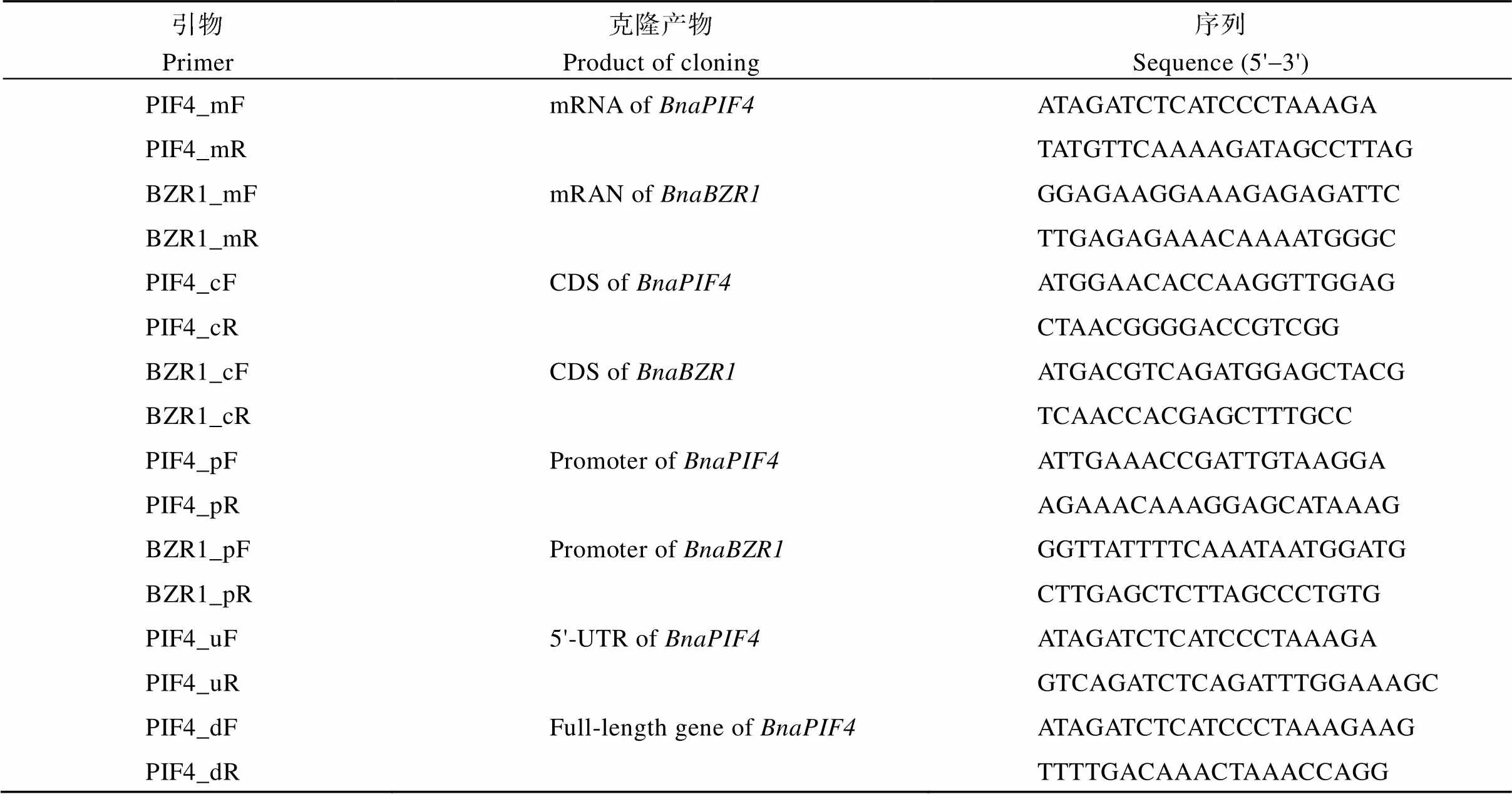
表1 基因克隆引物
1.5 RT-qPCR与WB
以RT-qPCR检测和基因表达, 以WB检测BnaPIF4、BnaBZR1及1,5-二磷酸核酮糖羧化酶(Ribulose-1,5-bisphosphate carboxylase, RuBPCase)等蛋白表达。以为内参基因, 采用2–ΔΔCt法以RT-qPCR检测和各拷贝表达, 根据序列特征设计所需引物(表2), 各试验均3次技术重复。
分别以HA-tag (ab18181, Abcam, Inc., 美国)(1∶1000)和RbcL抗体(AS03037, Agrisera, Ins., 瑞典) (1∶2000)为一抗进行WB, 检测连接HA标签的BnaPIF4蛋白和RuBPCase蛋白, 以ECL发光法进行印迹, 随后以GS800扫描仪(Bio-Rad Company, 美国)扫描蛋白光密度。
1.6 载体构建与遗传转化
用于拟南芥和甘蓝型油菜和遗传转化的载体来源于国家油料改良中心(湖南)。分别以3种基因的5¢-UTR替换载体中的的5'-UTR序列, 随后将携带HA标签的CDS序列插入载体。以农杆菌侵染介导进行拟南芥和甘蓝型油菜的遗传转化。
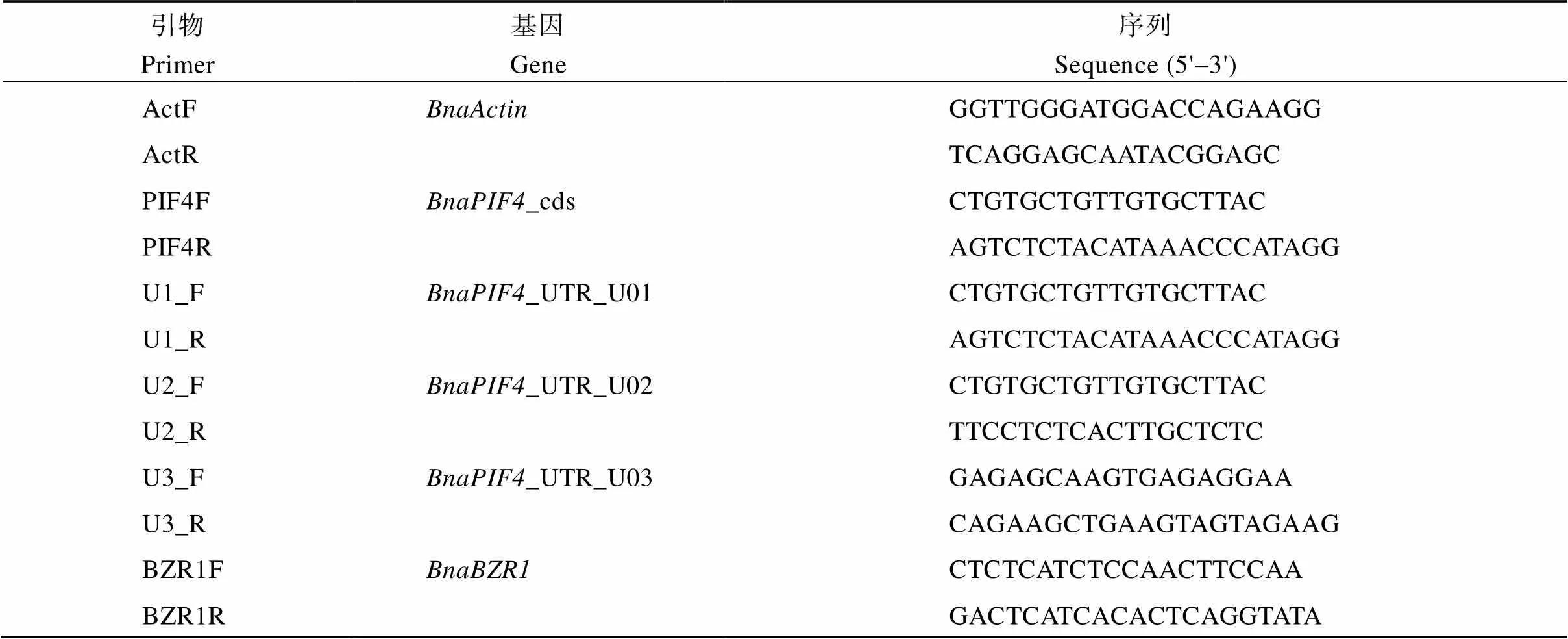
表2 RT-qPCR引物
1.7 酵母杂交实验
分别以作为酵母单杂交和双杂交的AD质粒, 以为酵母双杂交的BD质粒, 以为酵母单杂交的BD质粒, 共转化酵母菌株用于鉴定BnaPIF4和BnaBZR1之间的互作关系。
2 结果与分析
2.1 弱光和2,4-BL处理下XY881与XY883重要农艺性状
对大田栽培的XY881和XY883分别进行遮光和2,4-BL处理, 并统计其有效光合叶RuBPCase活性、全株总角果数、每角果种子粒数、成熟种子含油量、油酸含量及种子千粒重(图1)。
遮光处理下XY881和XY883表现出明显的敏感性差异, XY883叶片中RuBPCase含量下降较XY881更明显, 与之对应的XY883每角果中种子粒数和种子含油量也由明显下降, 而XY881每角果粒数和种子含油量均无明显下降。2,4-BL处理对XY881与XY883的影响规律相似, 油菜叶片中RuBPCase含量明显上升, 光合作用加强, 与之对应的全株总角果数和每角果种子粒数明显增多, 但整体而言XY883受影响更明显。无论弱光照胁迫或2,4-BL处理均未对XY881和XY883种子千粒重及种子油脂中油酸比例。由此推测弱光和2,4-BL处理下主要通过影响光合作用水平, 调控XY881和XY883种子油脂合成的底物, 该过程中XY881和XY883存在油菜素内酯信号响应因子表达水平差异。
2.2 两品系甘蓝型油菜(XY881和XY883)中BnaBZR1和BnaPIF4基因克隆和结构分析
为进一步理解和在XY881和XY883对弱光和2,4-BL响应差异中的作用, 本文从XY881和XY883中分别克隆和的全长mRNA和全长基因, 并从中亚克隆启动子、CDS和5¢-UTR等元件, 并对两品系中和基因结构进行分析(图2)。
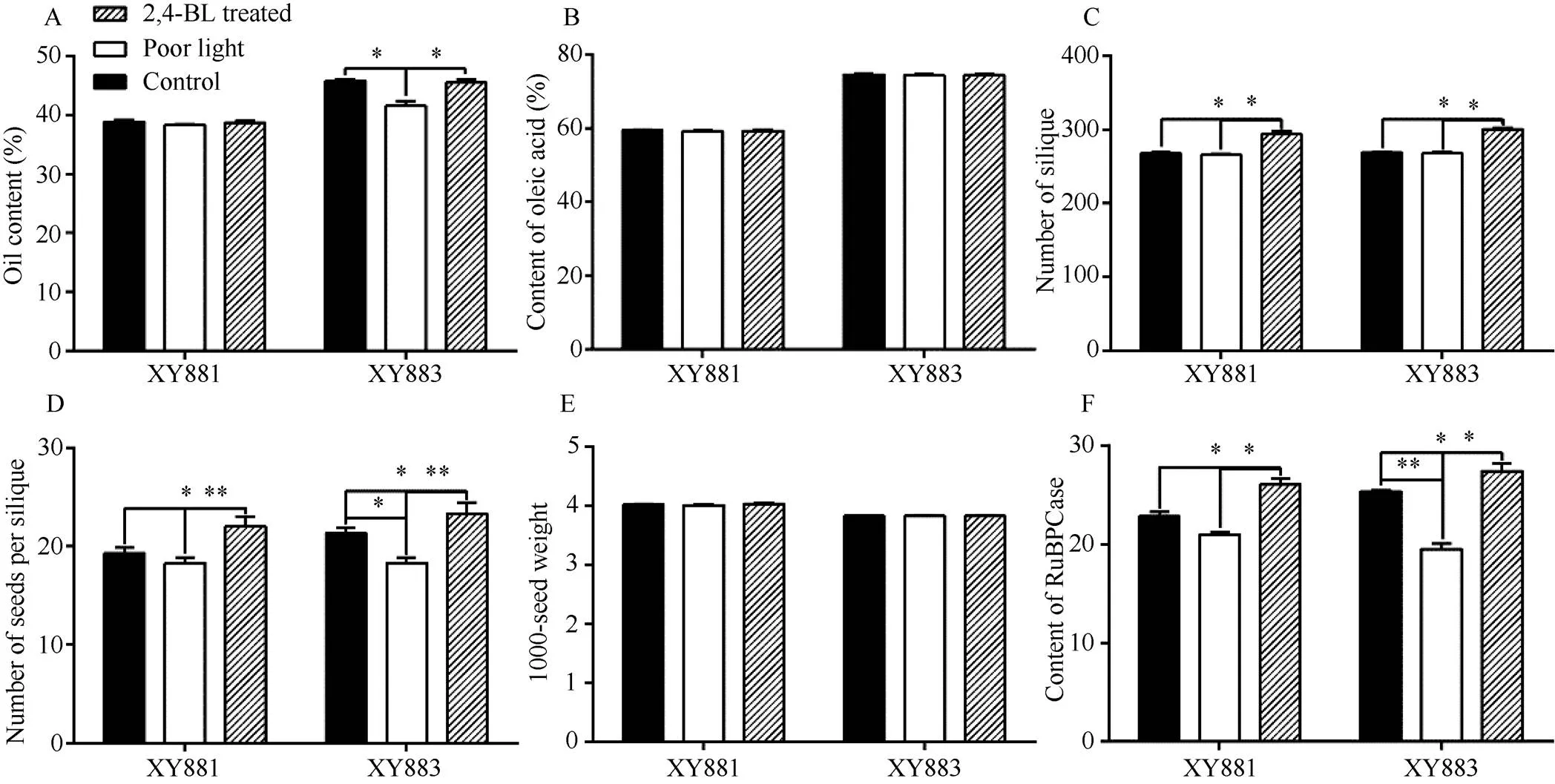
图1 弱光和2,4-BL处理下XY881和XY883重要农艺性状
A: 种子含油量; B: 种子油酸含量; C: 单株总角果数; D: 每角果种子粒数; E: 成熟种子千粒重; F: 叶片RuBPCase含量。*< 0.05, **< 0.01。
A: oil content of seed; B: oleic acid content of seed; C: total number of silique per plant; D: number of seeds per silique; E:1000-seed weight of mature seeds; F: RuBPCase content in leaves. *< 0.05, **< 0.01.
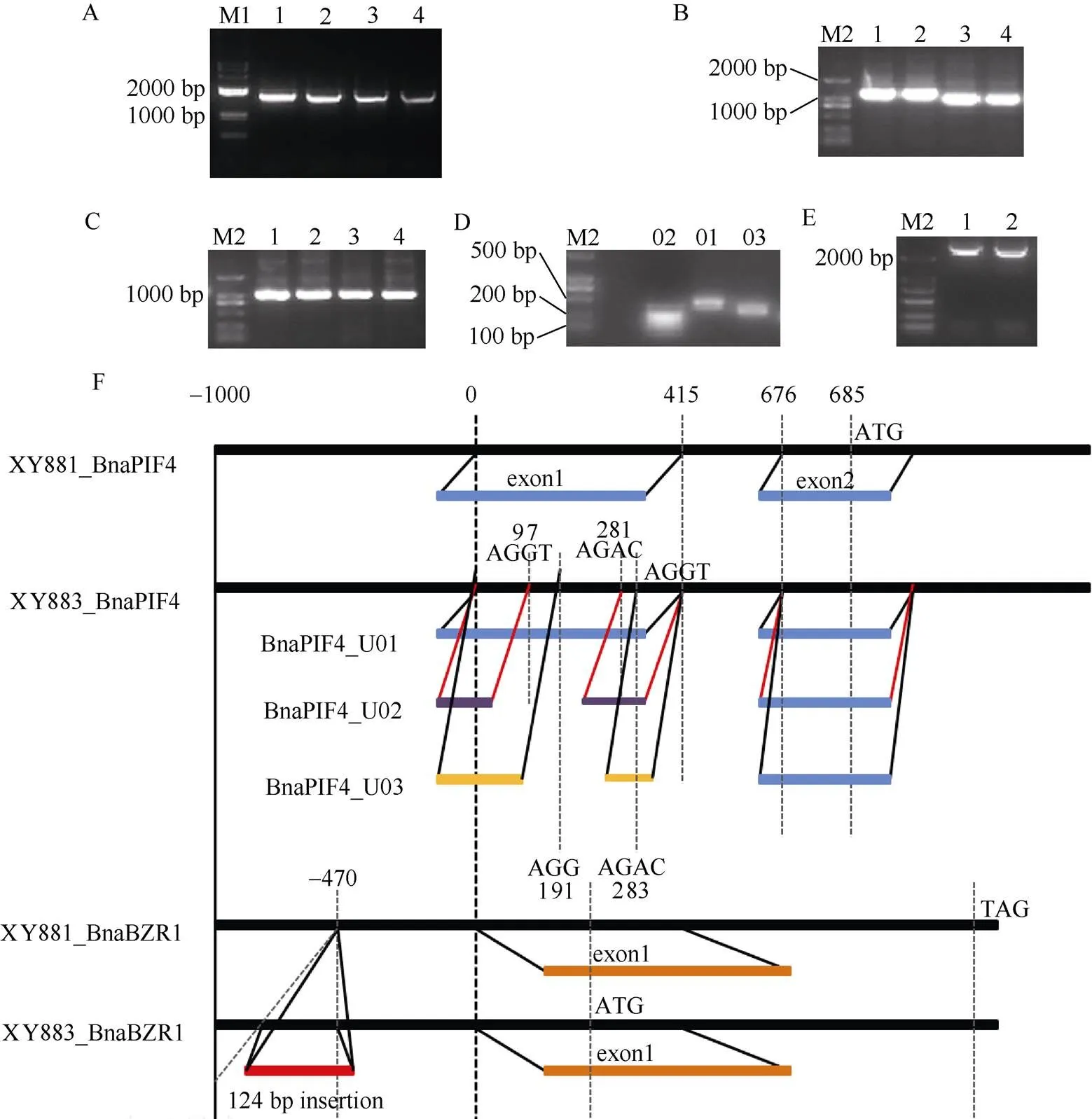
图2 XY881和XY883中BnaPIF4和BnaBZR1基因结构
A:和全长mRNA; B:和全长CDS; C:和启动子; D:三种5'-UTR; E:全长基因; F: XY881和XY883中和突变结构示意。1: XY881中; 2: XY883中; 3: XY881中; 4: XY883中; 01~03: XY883中三种5'-UTRs; M1: 2K plus DNA marker; M2: 2K DNA marker。
A: cloning of mRNA ofand; B: cloning of CDS ofand; C: cloning of promoter ofand; D: cloning of 5'-UTR of; E: cloning of full-lengthgene; F: the mutation structure ofandin XY881 and XY883. 1:of XY881; 2:of XY883; 3:of XY881; 4:of XY883; 01–03: three 5'-UTRs offrom XY883; M1: 2K plus DNA marker; M2: 2K DNA marker.
基因结构分析显示, XY881中和基因与亲本湘油15一致, 而XY883中和基因分别存在可变剪接和启动子插入突变。XY883中可变剪接共形成3种含有不同5¢-UTR的转录产物, 分别为U01 (424 bp)、U02 (239 bp)和U03 (332 bp) (图2-D); 该可变剪接均发生于亲本基因的第一外显子内, 3个5¢-UTR拷贝具有相同的剪接边界序列(AGGT和AGAC), 第一剪接点分别位于+97 bp、+191 bp和+415 bp位置。XY883中基因在启动子-470 bp位置内存在一个124 bp长度的富A/T碱基插入突变。
2.3 BnaBZR1启动子插入突变功能
为进一步确认XY883中基因启动子区域的插入突变有何功能, 分别以PlantCARE数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)鉴定插入序列中的启动子元件(图3-A); 将和的CDS克隆到载体(AD-PIF4和AD-BZR1), 将的CDS克隆到载体(pGBK-PIF4), 将和启动子克隆到载体(pHIS2-pPIF4、 pHIS2-pBZR1.1和pHIS2-pBZR1.2) , 分别进行酵母杂交, 验证BnaPIF4与BnaBZR1之间的互作关系是否受启动子突变影响(图3-B); 以RT-qPCR检测XY883和XY881中基因表达调控规律的异同点(图3-C, D)。
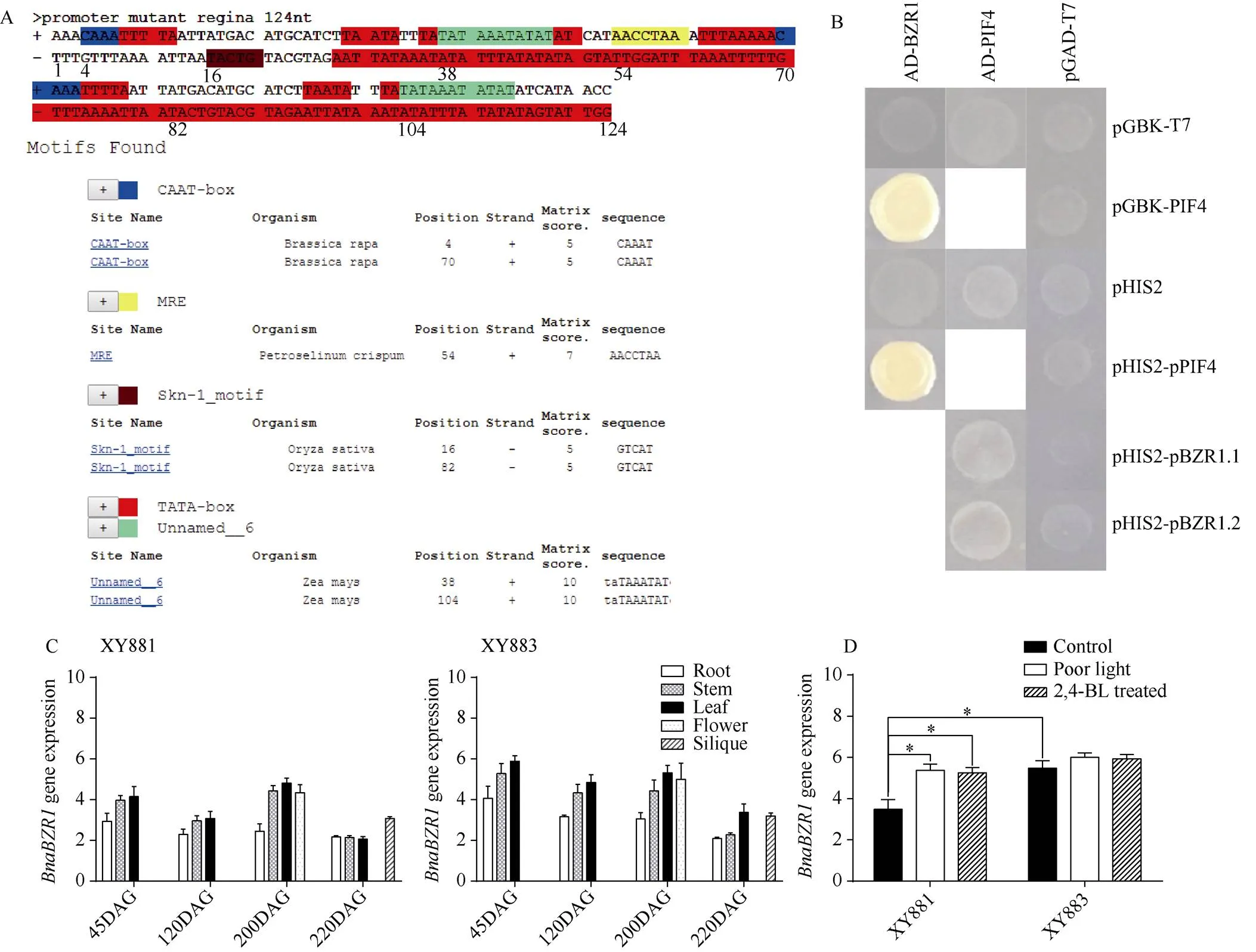
图3 BnaBZR1启动子插入突变、基因表达和BnaBZR1与BnaPIF4互作
A:启动子插入突变序列分析; B:和酵母杂交分析; C: XY881和XY883中时空表达; D: 弱光和2,4-BL处理下XY881和XY883中表达。DAG表示种子萌发后天数。
A: sequence analysis of insertion mutation regions in the promoter of; B: analysis of the interaction betweenandby yeast hybridization; C: the temporal and spatial gene expression ofin XY881 and XY883; D: the expression ofin XY881 and XY883 under poor light stress and 2,4-BL treatment. DAG means days after seed germination.
分析发现,基因启动子区124 bp的插入突变中含有大量的启动子元件, 包含多个TATA盒、2个CAAT盒、1个MRE 结构域及2个Skn-1结构域。酵母杂交结果表明, BnaBZR1与BnaPIF4蛋白可相互结合, BnaBZR1能结合启动子, 而BnaPIF4不能结合启动子,基因启动子插入突变不影响BnaPIF4与启动子的结合关系。XY883中启动子插入突变会明显提高其表达量, 苗期和苔期XY883中表达明显高于XY881, 叶片和茎等光合作用部位XY883中表达明显高于XY881。XY883中表达稳定, 不受低光和2,4-BL的明显诱导, 而XY881中表达受到低光和2,4-BL明显诱导, 刺激后XY881中表达明显提高, 这表明启动子插入突变可能提高本底表达水平, 同时造成表达对环境的响应减弱, 插入突变序列影响表达具体通过何种方式尚需进一步研究, 由插入突变中大量启动子元件来看, 该插入突变可能引入了上游转录因子的结合位点, 加强了启动, 但同时由于启动子元件的增加造成光响应和油菜素内酯响应的转录因子结合位点竞争抑制, 降低了表达对低光和2,4-BL的响应。
2.4 可变剪接影响BnaPIF4基因表达和翻译
通过RT-qPCR检测XY881和XY883中基因的时空表达规律以及在弱光胁迫和2,4-BL处理下不同转录产物表达的响应(图4-A, B)。将BnaPIF4可变剪接形成的不同转录产物分别克隆到载体上, 重组载体结构如图4-C, 随后将重组载体转化拟南芥, 观察表型并检测转基因拟南芥中表达情况及光合酶RuBPCase含量(图4-D~F)。
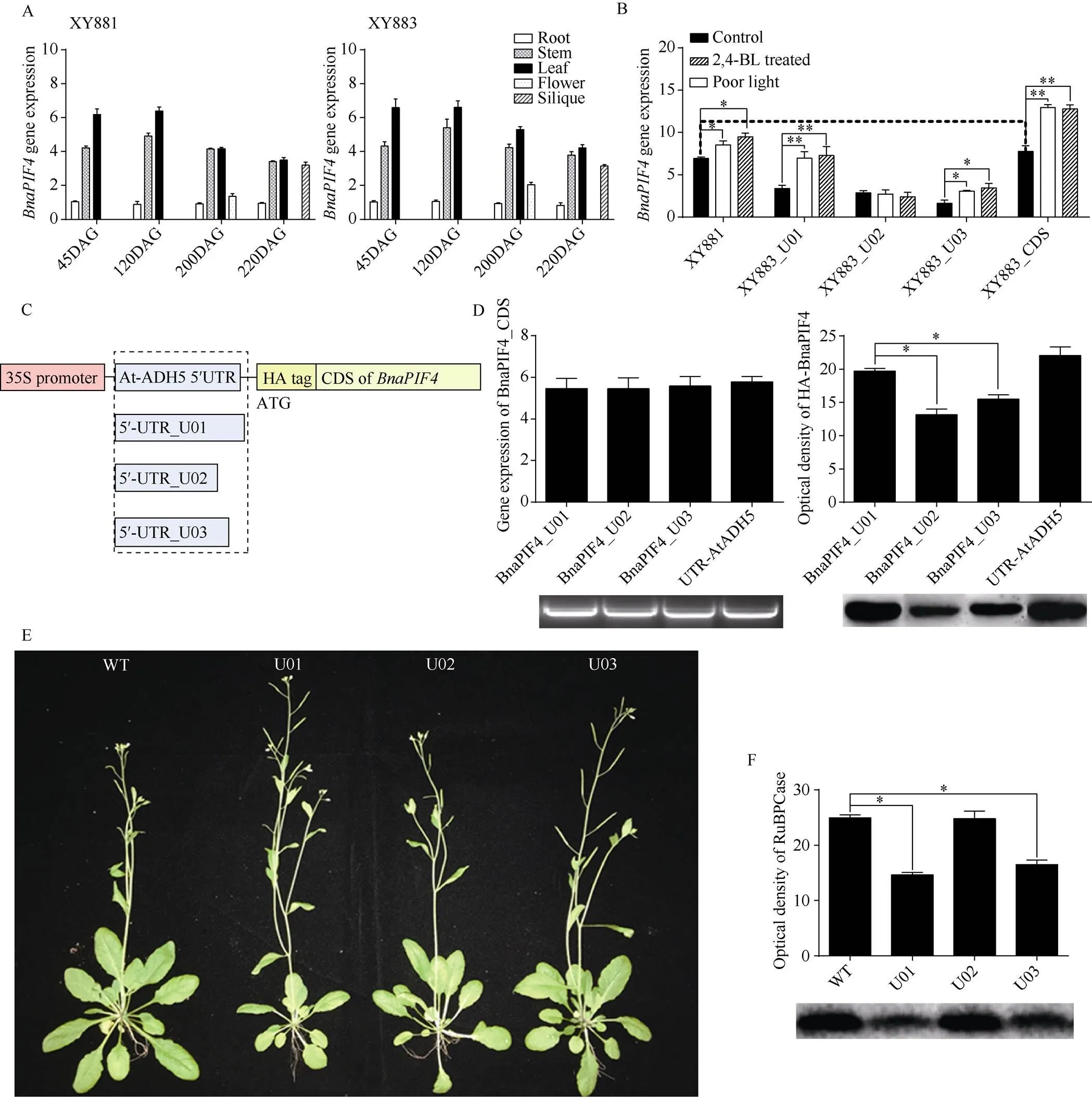
图4 BnaPIF4可变剪接对基因表达和翻译的影响
A: XY881和XY883中时空表达; B: 弱光和2,4-BL处理下XY881和XY883中表达; C: 含不同5'-UTRs重组载体结构; D: 转基因拟南芥中mRNA和蛋白表达; E: 转拟南芥表型; F: 转拟南芥RuBPCase含量。DAG表示种子萌发后天数。
A: temporal and spatial gene expression ofin XY881 and XY883; B: the expression ofin XY881 and XY883 under poor light stress and 2,4-BL treatment; C: construction ofvector with different 5'-UTRs; D: the mRNA expression ofand the protein expression of BnaPIF4; E: the phenotype of transgenic; F: the RuBPCase protein content of transgenic. DAG means days after seed germination.
时空表达检测发现XY881和XY883中总表达量模式无明显差异,表达均为随生育进程表达量逐渐降低, 有明显的组织特异性, 主要表达于地上部分, 叶中表达量最高。低光照和2,4-BL处理诱导表达, 且XY883中表达量增幅远大于XY881, 进一步分析发现不同转录产物对弱光和2,4-BL的响应具有明显差别, 其中转录本U01 (具有U01结构5¢- UTR, 即亲本无突变转录产物)响应最明显, U03转录产物次之(具有U03结构5¢-UTR), U02转录产物(具有U02结构5¢-UTR)无明显响应。拟南芥遗传转化发现不同的5¢-UTR在35S启动子驱动下不影响的CDS转录, 但不同的5¢-UTR对BnaPIF4蛋白合成具有显著影响, 其中U01与对照AtADH5具有相近的高水平BnaPIF4蛋白合成, U02和U03具有明显偏低的BnaPIF4蛋白合成量。转基因拟南芥表现出叶片狭长、茎伸长、早花及结实减少等表型, 转不同的转录产物, 效应具有差异, U01转录产物影响最大, U03次之, U02影响最小, 叶片光合关键酶RuBPCase含量的下降也表现U01最明显, U03次之, U02相对野生型无明显下降。BnaPIF4本身是bHLH家族重要的转录因子, 其下游存在大量的响应基因,不同转录产物的蛋白合成能力具有差异, 由此推测, 环境因素可能影响XY883中的剪接过程, 调控不同转录产物的比例, 一方面在转录水平响应上游因子的调控, 一方面在蛋白水平传导和改变对下游响应基因的调控水平, 通过自身在转录和翻译上平衡的调节来调控XY883对环境的综合响应, 这可能是XY883相对亲本湘油15在正常光照条件下具有更高的光合效能且对弱光等环境因素更敏感的原因之一。
2.5 甘蓝型油菜和拟南芥中BnaPIF4和BnaBZR1功能
为进一步验证和的功能, 分别对拟南芥和甘蓝型油菜亲本湘油15进行遗传转化, 观察过表达和后的表型(图5)。
转基因拟南芥相较野生型表现出叶片狭长、茎伸长、早花且结实率降低, 叶片中RuBPCase含量明显下降的表型, 成熟种子的含油量也明显下降,不同转录产物的效应存在差异, 转U01和U03拟南芥种子含油量下降明显, 转U02拟南芥种子含油量则无明显下降, 这与3种转录物对拟南芥叶片RuBPCase含量的影响一致。转基因拟南芥相较野生型具有明显更大的生物量, 生育期适度缩短, 总结实量上升, 但叶片RuBPCase含量和种子含油量无明显变化。共转化和基因拟南芥仍保持转基因拟南芥叶片狭长和早花的特征, 但植株生物量明显提升, 叶片RuBPCase含量和种子含油量相对野生型无明显变化。综合来看, 在拟南芥中表现为光合作用的负调控因子;表现生长促进作用, 提升拟南芥生物量;对的光合作用负效应具有拮抗作用。在亲本湘油15中过表达会让其主茎相对更细长, 叶片也更狭长, 开花略有提前; 在湘油15中过表达会提高其生物量, 提早花期, 增加开花量, 但均无统计学差异; 湘油15过表达和后叶片RuPBCase含量亦无明显变化。整体而言,和过表达对甘蓝型油菜的影响和拟南芥基本一致, 但对甘蓝型油菜的影响远小于拟南芥, 这可能与甘蓝型油菜复杂的双二倍体基因组中存在大量同源拷贝有关, 单一基因过表达影响远不如拟南芥明显。
3 讨论
甘蓝型油菜品系XY883和XY881均来源于亲本湘油15, XY881基本保持了亲本特性, XY883的RuBPCase含量、种子含油量、种子油酸含量和结实量相较亲本均明显提升, 弱光胁迫对XY881和湘油15影响较小, 而对XY883影响十分明显, 这表明XY883和XY881的表型差异可能源于光响应元件的差异。2,4-BL对XY881和XY883的调控规律基本一致, 均提升RuBPCase含量, 促进光合, 但影响幅度差异明显, 2,4-BL对和基因具有一致的诱导作用, 诱导效应存在品系差异[11], 这表明和的表达调控介导油菜素甾类信号和光信号通路的互作可能是XY881和XY883光合作用和弱光敏感性等差异的原因。
植物中BZR1是油菜素内酯信号通路中的关键转录因子, 其通过核质穿梭调控下游靶基因表达, 并通过反馈回路影响植物油菜素甾醇合成来调节植物的生长[12], 可见油菜中自身的表达调控同样可以影响油菜素内酯信号途径。XY883中启动子中的插入突变引入了多个具有转录因子特征的元件, 本底表达水平明显高于XY881, 但弱光胁迫和2,4-BL处理对XY883中的诱导效应较弱, 造成在胁迫和2,4-BL处理下两品系油菜中表达水平接近, 这可能是由于启动子插入突变之后增强了启动, 同时对上游转录调控因子产生竞争性结合, 抑制了弱光和2,4-BL的响应元件工作, 但具体的分子机制尚需进一步探索, 另一方面, XY883中对弱光和2,4-BL钝感, 导致内源油菜素内酯合成的反馈调节失效, 造成XY883对弱光照等不利环境因素的调节能力下降。
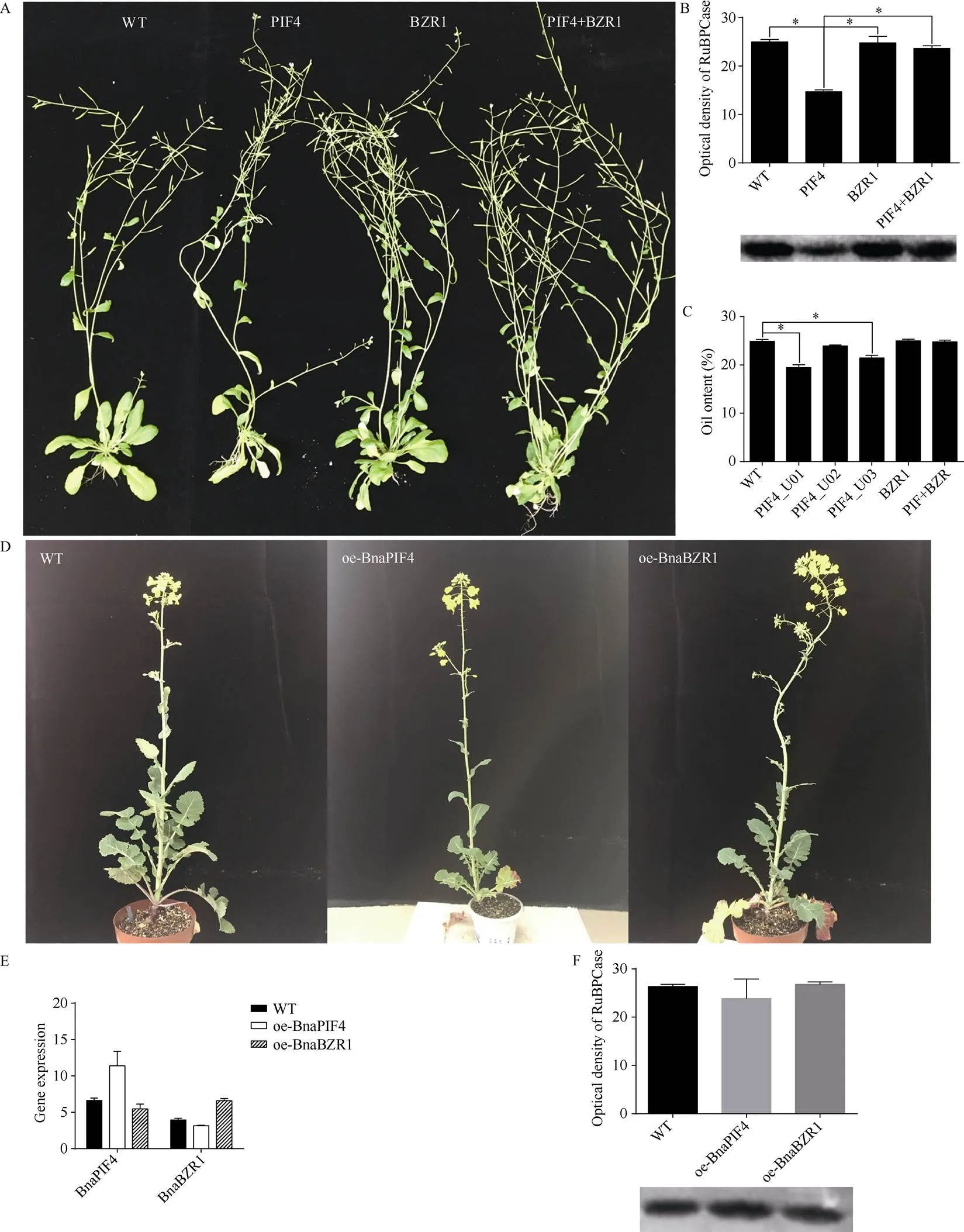
图5 转基因拟南芥和转基因油菜表型
A: 转基因拟南芥; B: 转基因拟南芥RuBPCase含量; C: 转基因拟南芥种子含油量; D: 转基因油菜; E: 转基因油菜中和表达; F: 转基因油菜RuBPCase含量。
A: the phenotype of transgenicand; B: the RuBPCase protein content of transgenicand; C: the oil content of transgenicseed; D: the plant of transgenicL.; E: the gene expression ofandin transgenicL.; F: the RuBPCase content of leaves of transgenicL.
PIF4是植物光信号通路中的核心转录因子之一,它通过响应光质变化[13]和热胁迫[14]等调节植物植物开花和生长等生物学过程。PIF4是介导光信号与植物激素途径相互作用的关键转录因子, 如与赤霉素信号[15]、油菜素内酯信号[16]和生长素信号[17]等产生互作。在XY883中选择性剪接形成3种具有不同5'-UTR的转录物, 弱光和2,4-BL对这3种转录物的诱导效应具有明显差别, 在转基因拟南芥中这3种转录物也表现明显不一致的翻译效率, 并导致转基因拟南芥产生不同的RuBPCase含量, 这表明XY883中的剪接调控可能是其与XY881存在明显光合水平和弱光敏感性差异的原因。XY883中通过可变剪接形成不同转录物, 造成转录水平和翻译水平的变化量存在明显差异, 在正常光照下XY883中低翻译水平的U02和U03转录物相对比例较高, 弱光下高翻译水平的U01转录物比例明显上升, 而BnaPIF4在拟南芥和油菜中都表现出光效负调控因子的特征, 这可能是XY883在常光下表现明显高于亲本光合效能, 而弱光下光合效能明显降低的原因。
酵母杂交实验表明, BnaBZR1蛋白与BnaPIF4相互作用, BnaBZR1与的启动子相互作用, 但BnaPIF4不与的启动子相互作用, 这表明BnaBZR1可能调节的表达, 而BnaPIF4可能不直接调控的表达。PIF4和BZR的互作是植物油菜素内酯信号和光信号互作的关键节点, BnaBZR1与BnaPIF4的互作模式可能意味着油菜中油菜素内酯信号与光信号的互作存在方向性偏好。前期研究显示在甘蓝型油菜中和均存在大量同源拷贝[9,18], 因此在功能上可能存在大量的冗余, 二者在甘蓝型油菜光效调节中可能存在更复杂的互作网络, 需要进一步对二者同源拷贝及上下游调控因子进行发掘和探索。
总体来看, 甘蓝型油菜中涉及油菜素内酯和光信号互作因子存在着基因多态性, 由此介导的油菜素内酯信号途径和光信号途径互作等方面差异可能是不同品系甘蓝型油菜光合效能差异的原因, 两信号途径之间互作的平衡可能是筛选稳定的高产油油菜品种的重要指标。
4 结论
甘蓝型油菜品系XY883相对亲本湘油15, 分别存在的可变剪接和启动子的插入突变, 导致两基因在不同条件下的表达出现不同于亲本的变化规律。在拟南芥和油菜中表现光合负效应因子的特点, 而可对此负效应形成拮抗, 拟南芥受和遗传转化影响较甘蓝型油菜更大。
[1] Oh E, Zhu J Y, Wang Z Y. Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses., 2012, 14: 802–809.
[2] Bai M Y, Shang J X, Oh E, Fan M, Bai Y, Zentella R, Sun T P, Wang Z Y. Brassinosteroid, gibberellin and phytochrome impinge on a common transcription module in., 2012, 14: 810–817.
[3] Gudesblat G E, Russinova E. Plants grow on brassinosteroids., 2011, 14: 530–537.
[4] Ryu H, Hwang I. Brassinosteroids in plant developmental signaling networks., 2013, 56: 267–273.
[5] Wang W, Bai M Y, Wang Z Y. The brassinosteroid signaling network—a paradigm of signal integration., 2014, 21: 147–153.
[6] USDA FAS (Foreign Agricultural Service), 2016. Oilseeds: World Markets and Trade. https://www.fas.usda.gov/data/oilseeds-world-markets-and-trade.
[7] Chalhoub B, Denoeud F, Liu S, Parkin I A, Tang H, Wang X, Corréa M. Early allopolyploid evolution in the post-Neolithicoilseed genome., 2014, 345: 950–953.
[8] Mølmann J A, Hagen S F, Bengtsson G B, Johansen T J. Influence of high latitude light conditions on sensory quality and contents of health and sensory-related compounds in swede roots (L. ssp.Metzg.), 2018, 98: 1117–1123.
[9] 冯韬, 官春云. 甘蓝型油菜光敏色素互作因子4 (BnaPIF4)基因克隆和功能分析. 作物学报, 2019, 45: 204–213. Feng T, Guan C Y. Cloning and characterization of phytochrome interacting factor 4 (BnaPIF4) gene fromL., 2019, 45: 204–213 (in Chinese with English abstract).
[10] Wei F, Gao G Z, Wang X F, Dong X Y, Li P P, Hua W, Wang X, Wu X M, Chen H. Quantitative determination of oil content in small quantity of oilseed rape by ultrasound-assisted extraction combined with gas chromatography., 2008, 15: 938–942.
[11] 冯韬, 谭晖, 徐江林, 官春云. 油菜素内酯在不同生育期对两品系甘蓝型油菜的生长调控. 中国油料作物学报, 2019, 41: 904–913. Feng T, Tan H, Xu J L, Guan C Y. Epibrassinolide regulation on oilseed rape (L.) in different period., 2019, 41: 904–913 (in Chinese with English abstract).
[12] Wang Z Y, Nakano T, Gendron J, He J X, Chen M, Vafeados D, Yang Y L, Fujioka S, Yoshida S, Asami T, Chory J. Nuclear- localized BZR1 m llklediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis., 2002, 2: 505–513.
[13] Casson S A, Franklin K A, Gray J E, Grierson C S, Whitelam G C, Hetherington A M. Phytochrome B and PIF4 regulate stomatal development in response to light quantity., 2009, 19: 229–234.
[14] Kumar S V, Lucyshyn D, Jaeger K E, Alós E, Alvey E, Harberd N P, Wigge P A. Transcription factor PIF4 controls the thermosensory activation of flowering., 2012, 484: 242–245.
[15] Lucas M D, Davière J M, Mariana R F, Pontin M, Manuel J I P, Lorrain S, Fankhauser C, Blázquez M A, Titarenko E, Prat S. A molecular framework for light and gibberellin control of cell elongation., 2008, 451: 480–484.
[16] Stella B G, Miguel D L, Cristina M, Ana E R, Davière J M, Prat S. BR-dependent phosphorylation modulates PIF4 transcriptional activity and shapes diurnal hypocotyl growth., 2014, 28: 1681–1694.
[17] Franklin K A, Lee S H, Patel D, Kumar S V, Spartz A K, Gu C, Ye S Q, Yu P, Breen G, Cohen J D, Wigge P A, Gray W M. PHYTOCHROME-NTERACTING FACTOR 4 (PIF4) regulates auxin biosynthesis at high temperature., 2011, 108: 20231–20235.
[18] 冯韬, 官春云. 甘蓝型油菜芸薹素唑耐受因子(BnaBZR1/ BnaBES1)全长CDS克隆与生物信息学分析. 作物学报, 2018, 44: 1793–1801. Feng T, Guan C Y. Cloning and characterization of brassinazole-resistant (BnaBZR1 and BnaBES1) CDS fromL., 2018, 44: 1793–1801 (in Chinese with English abstract).
Mechanism ofandregulating photosynthetic efficiency in oilseed rape (L.) under poor light
FENG Tao1,2, TAN Hui1,2, GUAN Mei2, and GUAN Chun-Yun2,*
1College of Biology and Agriculture, Zunyi Normal College, Zunyi 563000, Guizhou, China;2College of Agronomy, Hunan Agricultural University / National Oilseed Crops Improvement Center in Hunan, Changsha 410128, Hunan, China
Double-low seed rapeL. varieties XY881 and XY883 screened from the same parent Xiangyou 15 have significant differences in seed oil content, photosynthetic efficiency, and poor light sensitivity. Brassinazole-resistant 1 () and phytochrome interacting factor 4 ()genes were cloned from XY881 and XY883, respectively, and their sequence structure, gene expression and gene function were analyzed. There were structural mutations in theandof XY883, causing differences in gene expression and regulation. The promoter ofin XY883 had a 124 bp A/T-rich insertion mutation, and XY883 had a higher expression ofthan XY881, and fewer expression changes under low light and 2,4-epibrassinolide(2,4-BL) treatment. Splicing differences ofin the 5'-UTR region were found in XY883, and the alternative splicing resulted in three 5'-UTRs of 424 bp (U01), 239 bp (U02), and 332 bp (U03). Under the induction of low light and 2,4-BL treatments the changed of three alternative splicedtranscripts in XY883 had significant differences. The three 5'-UTRs combined with the CDS ofwere transformed into. There was no significant difference ingene expression, but a significant difference in protein expression, indicating that the 5'-UTR mutation ofaffects protein synthesis.Transgenicshowed a phenotype with increased plant height, narrow leaves, and decreased photosynthesis. Co-transformation whitcould attenuate the decrease in photosynthesis caused by. The effects ofandgenes onL. were similar to those on, while the phenotype was not as obvious as that of, suggesting thatis a negative regulator of photosynthesis in rapeseed, andcan antagonize the negative regulation of photosynthesis by, while is consistent with the regulation pattern and photosynthetic phenotype of the two genes in XY881 and XY883.
L.;;; alternative splicing; insertion mutation
10.3724/SP.J.1006.2020.94198
本研究由国家重点基础研究发展计划(973计划)项目(2015CB150206)资助。
This study was supported by the National Basic Research Program of China (973 Program) (2015CB150206).
官春云, E-mail: guancy2011@aliyun.com
E-mail: 812298771@qq.com
2019-12-16;
2020-03-24;
2020-04-03.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20200403.0932.002.html
猜你喜欢
现代食品(2022年11期)2022-12-06
作物学报(2022年12期)2022-10-14
现代食品科技(2022年5期)2022-05-30
作物学报(2022年4期)2022-02-10
作物学报(2022年4期)2022-02-10
作物学报(2022年3期)2022-01-22
今日农业(2021年2期)2021-11-27
福建农林大学学报(自然科学版)(2021年5期)2021-10-08
今日农业(2020年23期)2020-12-31
福建农业科技(2016年10期)2016-03-07
