
利用CRISPR/Cas9技术编辑水稻温敏不育基因TMS5
2020-07-21 08:03陈日荣周延彪王黛君赵新辉唐晓丹许世冲唐倩莹符星学刘选明杨远柱1
作物学报 2020年8期
陈日荣 周延彪 王黛君 赵新辉 唐晓丹 许世冲 唐倩莹 符星学 王 凯 刘选明 杨远柱1,2,,,*
利用CRISPR/Cas9技术编辑水稻温敏不育基因
陈日荣1,**周延彪2,3,**王黛君5赵新辉3唐晓丹3许世冲4唐倩莹3符星学3王 凯3刘选明1,*杨远柱1,2,3,4,*
1湖南大学生物学院 / 植物功能基因组学与发育调控湖南省重点实验室, 湖南长沙 410082;2袁隆平农业高科技股份有限公司 / 农业农村部南方水稻品种创制重点实验室, 湖南长沙 410001;3湖南亚华种业科学研究院, 湖南长沙 410119;4华中农业大学植物科学技术学院, 湖北武汉 430070;5湖南师范大学生命科学学院, 湖南长沙 410081
是生产上应用最广泛的温敏不育基因。为创制新型水稻温敏核不育系, 利用CRISPR/Cas9基因编辑技术将6个优异的粳稻和4个优异的籼稻的基因敲除, 获得了温敏核不育系。比较发现, 粳稻温敏不育系ZG75S、CYGS、YG0618S、ZG07S、T0361S、7679S的不育起点温度在28~32°C之间, 籼稻温敏核不育系2537S、6150S、6379S的不育起点温度在24~28°C之间, 而籼稻温敏核不育系1109S的不育起点温度低于23.5°C。说明不同遗传背景材料获得的突变体的不育起点温度不一样, 通过的敲除可能获得不育起点温度较低的温敏核不育系。利用1109S与优质父本8048选配出优质高产杂交稻组合1109S/8048。大田试验表明, 1109S/8048比区试对照丰两优4号增产13.1%。温敏核不育系1109S及高产杂交稻组合1109S/8048的创制为高产育种提供了新的途径。
水稻; CRISPR/Cas9; 温敏不育;; 杂交组合
水稻是我国第一大粮食作物, 持续保持或提高水稻产量是关系国计民生的头等大事和保障国家粮食安全的迫切需求。杂交水稻的产量一般比常规水稻增产20%~30%, 为我国和世界粮食安全做出了重要贡献[1]。
两系法杂交水稻是我国发明的水稻杂种优势利用方法, 与三系法杂交稻相比, 两系法杂交水稻配组自由, 选育优质高产的组合几率大, 并且种子生产成本低、育种环节减少, 已经成为我国杂交水稻的主要栽培类型[2]。水稻两系不育系分为光敏核不育系(PGMS)和温敏核不育系(TGMS), 其中温敏核不育系是应用最广泛的两系不育系。2014年, Zhou等[3]克隆了安农S-1和株1S的温敏不育基因并阐明了其分子机理。编码具有前体tRNA 3'端内切酶活性的RNase ZS1蛋白, 通过调控受温度诱导的UbmRNA 的表达量, 控制水稻花粉发育。在温敏不育系中, 由于RNase ZS1功能缺失, 高温诱导表达的UbmRNA过度积累, 导致花粉败育。
CRISPR/Cas9基因编辑技术是指在基因组水平上对DNA序列进行定点改造的遗传操作技术, 由于操作简单、靶基因突变效率高等特点, 已广泛应用于基因功能研究和作物遗传改良方面[4-6]。Li等[7]以中花11为受体, 通过CRISPR/Cas9技术对负调控水稻产量基因(每穗实粒数)、(直立型密穗)、(粒长和粒重)和(穗粒数、分蘖相关)进行定点编辑, 发现突变体每穗实粒数增加,突变体每穗着粒密度增加的同时有半矮化现象,突变体在粒长增加的同时芒也增长。Sun等[8]利用CRISPR/Cas9技术对水稻淀粉分支酶IIb基因进行定点编辑, 与野生型相比,缺失突变体的粒形显著变小、直链淀粉和总淀粉含量都显著增加, 直链淀粉最高可达25%。周延彪等[2]利用CRISPR/Cas9基因编辑技术敲除粳稻基因, 获得了反光敏不育水稻新种质。
本研究以为靶基因, 运用CRISPR/Cas9技术进行基因的定点编辑, 分析了功能缺失突变体在不同温度处理下的花粉育性, 并对不育起点温度低于23.5°C的1109S进行测配, 为两系不育系和高产杂交水稻组合的选育提供新的思路。
1 材料与方法
1.1 水稻材料和CRISPR/Cas9载体构建
选用6个粳型常规稻浙粳75 (ZG75)、川引粳(CYG)、越粳0618 (YG0618)、ZG07、T0361、7679, 5个籼型常规稻2537、6150、6379、1109和8048, 2个杂交稻组合1109S/8048和区试对照丰两优4号。
(LOC_Os02g12290)靶点序列参照已有研究设计的靶位点[1], 在第1外显子上设计20 bp的靶点序列5′-CAGCGGCAAGTCATCGCCGG-3′, PAM序列为CGG (图1-A)。利用合成引物TMS5-target-F/ TMS5-target-R (表1)制成靶位点接头。18T-Cas9基因编辑中间载体为中国科学院上海植物生理生态研究所李来庚研究员馈赠。通过I酶切, 并将接头与酶切后的18T-Cas9通过T4连接酶连接, 转化大肠杆菌DH5a。M13F (表1)与TMS5-target-R进行菌液PCR 检测, 阳性菌液提取重组质粒18T-Cas9- TMS5。pCAMBIA1301和18T-Cas9-TMS5载体通过双酶切(I和d III)回收目的片段, 再通过T4DNA连接酶构建植物表达载体pCAMBIA1301- Cas9-TMS5-gRNA (图1-B), 并电击转化农杆菌EHA105。通过农杆菌介导法转化浙粳75 (ZG75)、川引粳(CYG)、越粳0618 (YG0618)、ZG07、T0361、7679、2537、6150、6379、1109成熟胚诱导的愈伤组织, 通过潮霉素抗性筛选T0代组培苗。
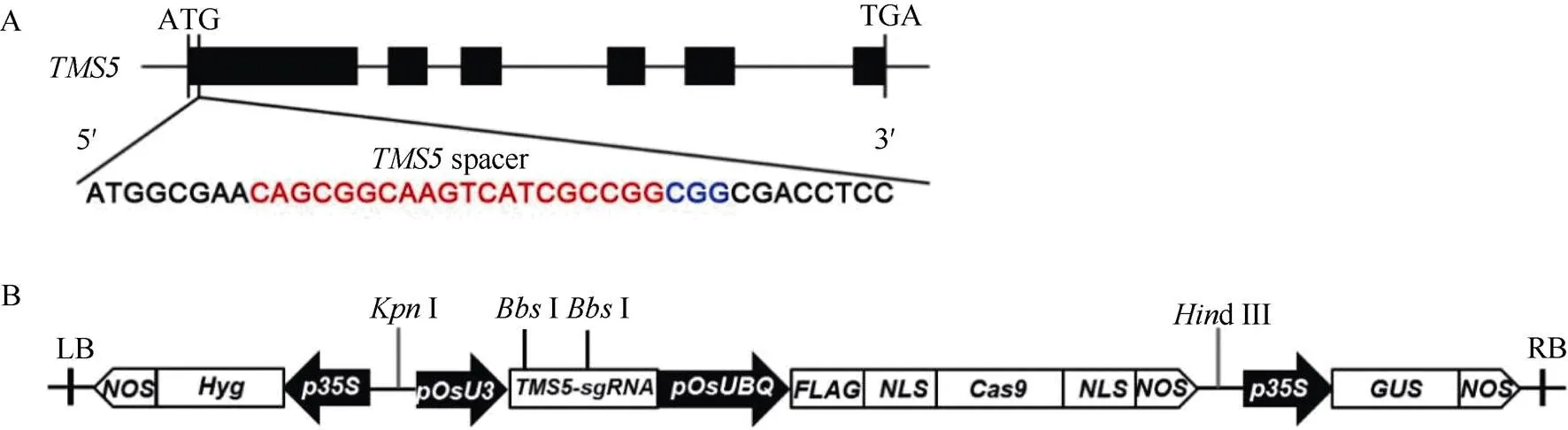
图1 CRISPR/Cas9-TMS5载体构建示意图
A:靶点位置。红色字母为靶点序列, 蓝色字母为PAM序列。B: pCAMBIA1301-Cas9-TMS5-gRNA重组载体结构示意图。LB: 左边界; RB: 右边界。
A: Schematic diagram of the targeted site in. The red letters are the target genome sequences. The blue letters are the protospacer adjacent motif (PAM) sequences. B:Schematic diagram of the pCAMBIA1301-Cas9-TMS5-gRNA construct. LB: left border; RB: right border.
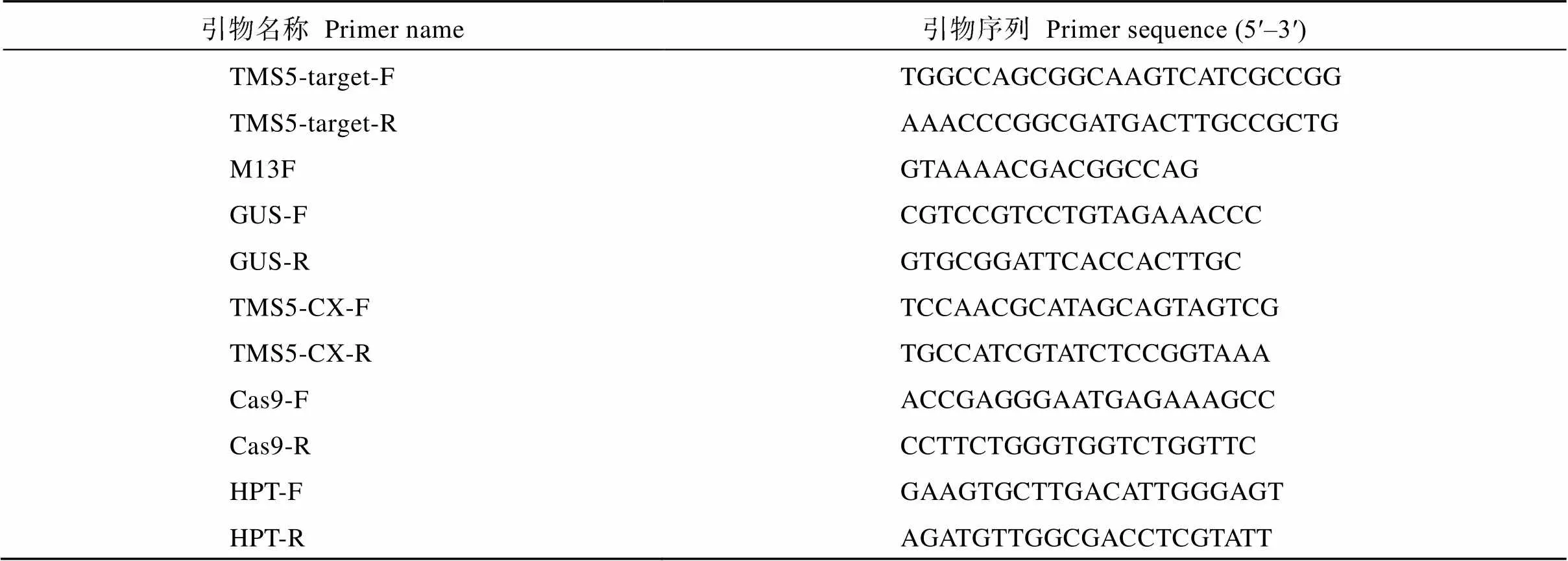
表1 本研究所用的引物
1.2 阳性转基因植株的获得及靶点测序分析
阳性农杆菌转化粳稻品种ZG75、CYG、YG0618、ZG07、T0361、7679以及籼稻品种2537、6150、6379、1109成熟胚的愈伤组织, 获得T0代组培苗。利用载体上GUS的特异性引物GUS-F/GUS-R进行T0代阳性转基因植株检测, 扩增产物718 bp的片段为阳性转基因植株。同时, 通过GUS组织化学染色[9]快速鉴定阳性转基因植株。为了检测靶位点的突变情况, 在靶点的5′端和3′端分别设计测序引物TMS5-CX-F/TMS5-CX-R (表1), 扩增目的片段后送测序公司测序。
1.3 无T-DNA元件tms5突变株的获得
T0代阳性转基因植株通过自交获得T1代种子。T1代种子播种成苗后, 取幼嫩的叶片进行GUS组织化学染色。为了鉴定无T-DNA元件的单株, 分别选取每个品种的亲本作为阴性对照, GUS染色呈蓝色的样品为阳性对照, 以及GUS染色不呈蓝色的5个单株, 通过CTAB法提取DNA[10], 用引物HPT-F/HPT-R、Cas9-F/Cas9-R以及GUS-F/GUS-R进行PCR扩增, 3对引物都不能扩增出目的条带的植株为不携带载体序列的突变株,基因为对照。筛选的不携带载体序列的突变株通过TMS5-CX-F/TMS5- CX-R引物进行PCR扩增, 将PCR产物送测序公司测序, 分析的突变类型。
1.4 tms5突变体育性光温处理与花粉育性鉴定
将T1代无T-DNA元件的突变体以及对照, 于2019年5月14日大田播种, 6月8日移栽, 参照丁颖的水稻幼穗分化八期划分标准[11], 当突变体主茎幼穗发育至六期(温敏核不育系育性敏感期为幼穗分化四至六期, 其中以五至六期最为敏感)的时候, 从大田移到人工培养箱进行光温处理, 分别设置24°C、28°C和32°C, 光照条件为12 h光照/12 h黑夜, 处理14 d后, 在自然条件下生长, 取主茎外的其他茎节上的花粉用碘-碘化钾染色[1-2], 调查花粉育性。
另外, 采用冷灌池鉴定法详细调查1109的突变体的不育起点温度。即当1109的突变体主茎幼穗发育至六期时, 移至人工冷灌池, 设定21.5°C、22.5°C和23.5°C三个梯度, 处理10 d后移至自然条件下生长, 取主茎外的其他茎节上的花粉进行碘-碘化钾染色[1-2], 调查花粉育性。
1.5 大田农艺性状调查
参考Zhou等[12]关于农艺性状的方法, 于2019年5月14日大田播种父本8048、杂交组合1109S/ 8048和丰两优4号, 6月8日移栽。在成熟期分别选择父本8048、杂交组合1109S/8048和丰两优4号各20株, 考察株高、分蘖数、千粒重、穗长、穗粒数、结实率和产量。采用检验统计分析数据。
2 结果与分析
2.1 转基因植株的鉴定及突变位点分析
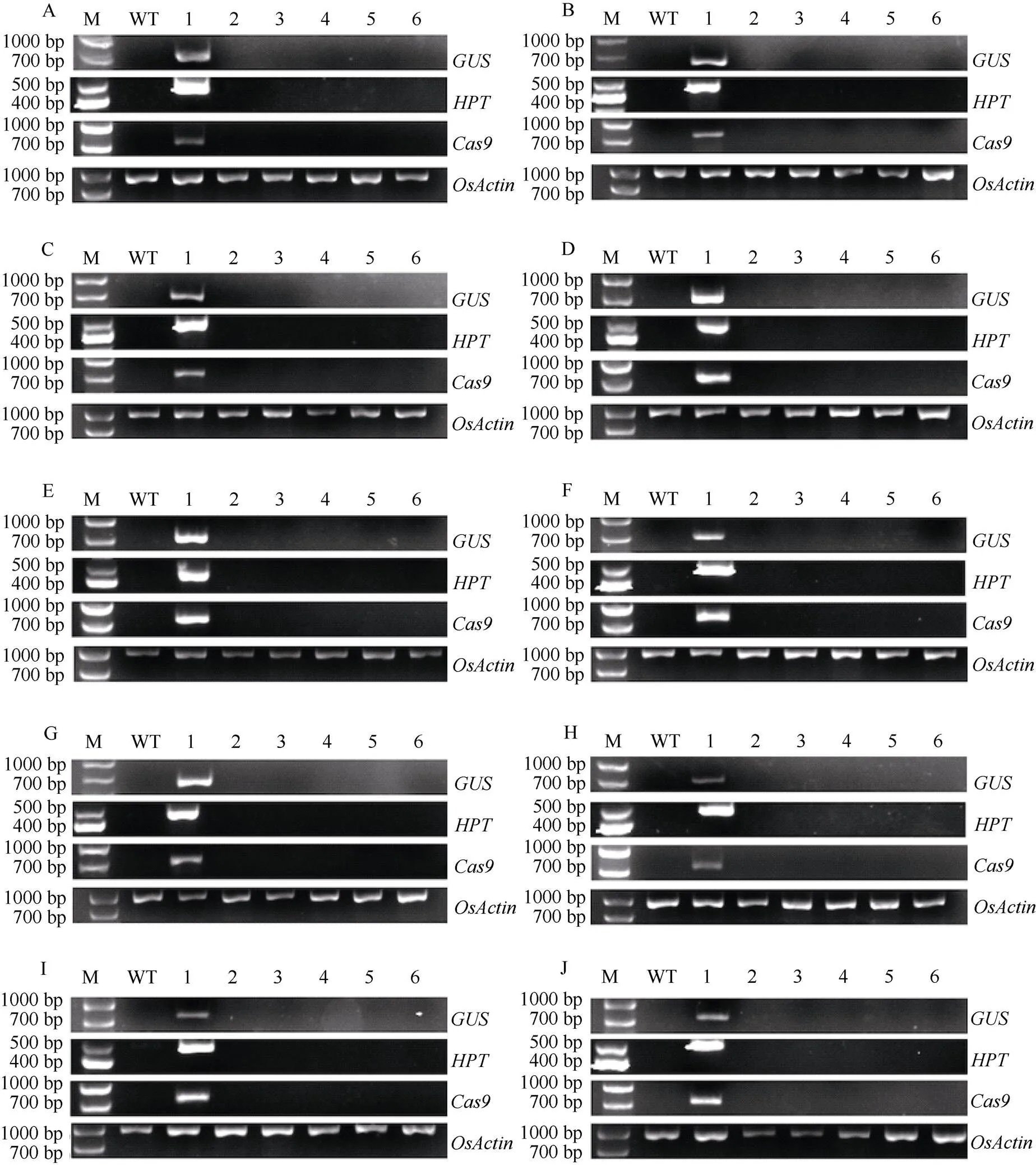
通过GUS的特异性引物GUS-F/GUS-R (表1)对T0代组培苗进行PCR扩增, 以及叶片进行GUS组织化学染色筛选阳性转基因植株(图2)。PCR能扩增出目的片段并且叶片呈蓝色的为阳性转基因植株。对阳性转基因植株的测序, 筛选纯合突变的T0代植株。纯合突变主要来自PAM前第3和第4碱基之间有单碱基A、T和G的插入, 以及单碱基和2个碱基的缺失(图3)。ZG75、CYG、YG0618、ZG07、T0361、7679、2537、6150、6379和1109的纯合突变率分别为25.0%、13.0%、11.8%、19.2%、18.8%、21.1%、8.0%、10.3%、9.4%和18.8%。

图2 T0代转基因植株的鉴定
ZG75(A)、CYG(B)、YG0618(C)、ZG07(D)、T0361(E)、7679(F)、2537(G)、6150(H)、6379(I)、1109(J)阳性转基因植株的PCR鉴定(上)和GUS组织化学染色(下)。M: DNA marker; 1~10: 转基因植株。
Identification of the positive transgenic plants by PCR (upper) and GUS histochemical staining (lower) in ZG75(A), CYG(B), YG0618(C), ZG07(D), T0361(E), 7679(F), 2537(G), 6150(H), 6379(I), and 1109(J). M: DNA marker; 1–10: transgenic plants.

图3 T0代纯合突变类型
红色字母为靶点序列; 蓝色字母为PAM序列; 绿色字母为插入的碱基; 横线表示碱基缺失; +表示插入; -表示缺失; WT表示野生型。
The red letters are the target genome sequences; the blue letters are PAM; the green letters are the insert base; the horizontal line indicates the missing base; +: insertion; -: deletion; WT: wild-type.
对T1代叶片GUS染色不呈蓝色的单株进行PCR检测, 筛选不含转基因元件的植株(附图1)。结果表明, 选取的叶片GUS染色不呈蓝色的单株都为不携带载体序列的突变株。测序发现, T1代非转基因植株的都为纯合突变, 且突变类型与T0代的纯合突变类型一致, 将这些非转且纯合突变的单株, 参考黄忠明等[1]的方法用于不育起点温度的鉴定。
2.2 tms5突变体不育起点温度的鉴定
将T1代纯合突变且不携带载体序列的突变株的植株用于不育起点温度的鉴定。当突变体的主茎幼穗发育到六期, 将突变体分别置24°C、28°C和32°C恒温培养箱培养14 d, 然后对主茎外的其他茎节上的花粉进行镜检。的粳稻突变体在32°C培养下, 花粉都表现为不育; 在28°C培养下, 花粉都表现为部分不育; 而在24°C培养下, 花粉都表现为可育(图4-A)。说明ZG75S、CYGS、YG0618S、ZG07S、T0361S、7679S的不育起点温度在28~32°C之间。的籼稻突变体在32°C和28°C培养下, 花粉都表现为不育, 而在24°C培养下,只有1109S的花粉表现为完全不育, 其他品种的花粉表现为可育(图4-B)。说明2537S、6150S、6379S的不育起点温度在24~28°C之间。进一步, 通过冷灌池设定21.5°C、22.5°C、23.5°C, 在1109S幼穗敏感期时分别处理, 处理后镜检花粉(图4-C)表明, 在21.5°C冷灌条件下, 1109S的花粉为完全可育; 22.5°C冷灌条件下, 1109S的花粉为部分可育; 23.5°C条件下, 花粉为完全不育。说明1109S的不育起点温度在22.5~23.5°C之间。
2.3 籼型温敏核不育系1109S的配组优势
由于1109S的不育起点温度较低, 在长沙具有明显的可育期和不育期, 育性转换明显, 不育性稳定, 具有较高的应用价值。为了鉴定1109S的配合力, 我们将1109S与不同父本测配, 筛选了一个具有较强杂种优势的组合1109S/8048。其生长量明显比8048大, 且具大穗表型(图5-A, B)。与8048相比, 1109S/8048在株高、有效穗、千粒重、穗长、穗粒数和产量等方面都显著增加, 其中产量增加25.6%, 具有典型的超亲优势(表2)。同时, 与区试对照丰两优4号相比, 1109S/8048的株高矮, 千粒重降低, 但有效穗、穗长、穗粒数和产量明显增加, 其中产量增加13.1% (表2), 表明1109S/8048杂交组合具有较强的杂种优势。
3 讨论
基因组编辑(Genome editing)技术是指在基因组水平上对DNA序列进行定点修饰和改造的遗传操作技术。现有的基因编辑系统主要包括锌指核酸酶 (Zinc finger nucleases, ZFNs) 系统、类转录激活因子效应物核酸酶(Transcription activator-like effector nucleases, TALENs)系统以及CRISPR/Cas (Clustered regularly interspaced short palindromic repeat-associated protein)系统[13]。其中, CRISPR/Cas9基因编辑技术由于载体构建过程简单、编辑效率高等优点, 成为当前主流的基因编辑系统, 并广泛应用于作物遗传改良方面[14-17]。2014年,基因的克隆及其控制育性分子机制的阐明, 为利用基因编辑技术创制温敏两用核不育系奠定了基础[3]。本研究通过CRISPR/Cas9基因编辑技术敲除粳稻和籼稻中基因, 比较粳稻和籼稻温敏不育系的不育起点温度, 筛选低不育起点温度的籼型温敏不育系, 并配制高产杂交稻组合, 为水稻的高产育种提供了新的途径。

图4 tms5突变体在不同温度处理条件下的花粉育性
A:粳稻突变体的花粉育性; B:籼稻突变体的花粉育性; C: 1109S在冷灌处理条件下的花粉育性。
A: Pollen fertility ofmutant ofrice; B: Pollen fertility ofmutant ofrice; C: Pollen fertility of 1109S under cold irrigation conditions.
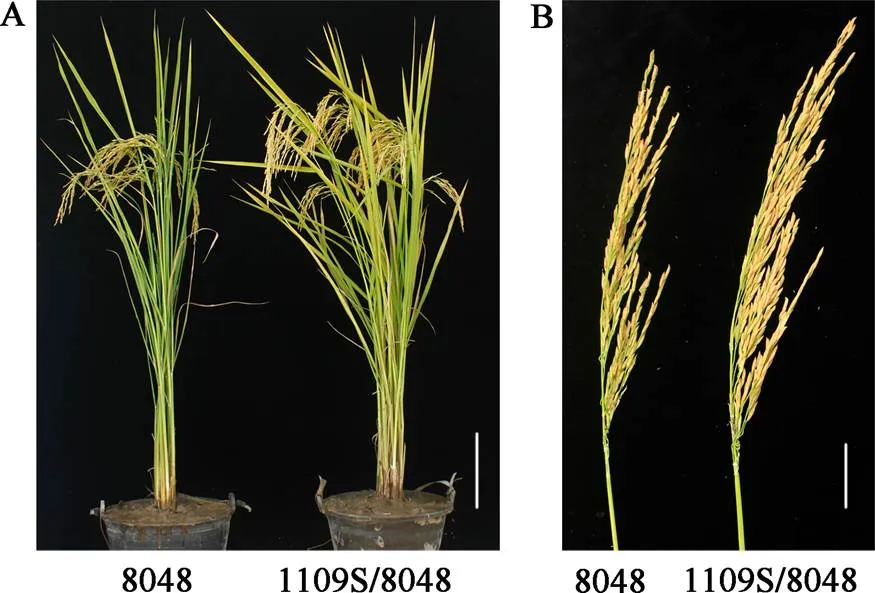
图5 1109S/8048杂交组合的表型鉴定
A: 1109S/8048杂交组合的表现, Bar = 20 cm; B: 1109S/8048杂交组合穗的表现, Bar = 5 cm。
A: Phenotype of 1109S/8048 hybrid combination, Bar = 20 cm; B: Panicle performance of 1109S/8048 hybrid combination, Bar = 5 cm.
是控制我国温敏型不育系的主要不育基因, 它编码一个保守的RNA酶ZS1, 具有前体tRNA 3′端内切酶活性, 通过抑制受温度诱导的UbmRNA 的表达量, 控制水稻花粉发育;基因CDS的71位碱基由C突变为A, 导致翻译提前终止, 使UbmRNA过度积累导致雄性不育[3-4]。Zhou等[14]通过CRISPR/Cas9基因编辑技术对11个水稻品种的进行敲除, 在1年内获得了不含转基因成分的温敏不育系, 这些不育系材料因遗传背景的差异表现出不同的温敏不育起点温度。中花11、GAZS、ZS97BS在28°C培养条件下花粉完全不育, ReBS、WSSMS、TFBS在26°C培养条件下花粉完全不育, 而YJSMS、ZZBS在24°C培养条件下花粉完全不育。本研究利用CRISPR/Cas9技术对6个粳稻和4个籼稻的基因进行敲除, 并对突变体进行不育起点温度鉴定, 发现粳稻温敏不育系ZG75S、CYGS、YG0618S、ZG07S、T0361S、7679S的不育起点温度在28~32°C之间, 籼稻温敏不育系2537S、6150S、6379S的不育起点温度在24~28°C之间, 而1109S的不育起点温度在22.5~23.5°C之间。Zhou等[14]通过CRISPR/Cas9基因编辑技术获得低不育起点温度的YJSMS、ZZBS, 在24°C培养条件下花粉完全不育, 本研究获得的1109S在23.5°C条件下完全不育, 因此1109S比YJSMS与ZZBS具有更低的不育起点温度。温敏核不育系转育起点温度的高低是决定两系杂交水稻制种安全的重要因素, 选育不育起点温度较低的温敏核不育系可以显著提高两系杂交水稻制种的纯度。育性转换起点温度23.5°C被认为是生产应用中比较安全的一个不育起点温度。因此, 通过基因编辑技术敲除基因, 筛选不育起点温度低的温敏不育系是决定能否应用的关键。由于1109S在23.5°C培养条件下完全不育, 因此在生产上具有较大的应用价值。以上这些结果说明不同遗传背景材料获得的突变体的不育起点温度不一样, 粳稻突变体的不育起点温度普遍比籼稻的要高。只是育性的一个开关, 而与不育起点温度无关, 不育起点温度受其他基因的调控。如何选取遗传背景材料敲除获得低不育起点温度的突变体有待进一步研究。
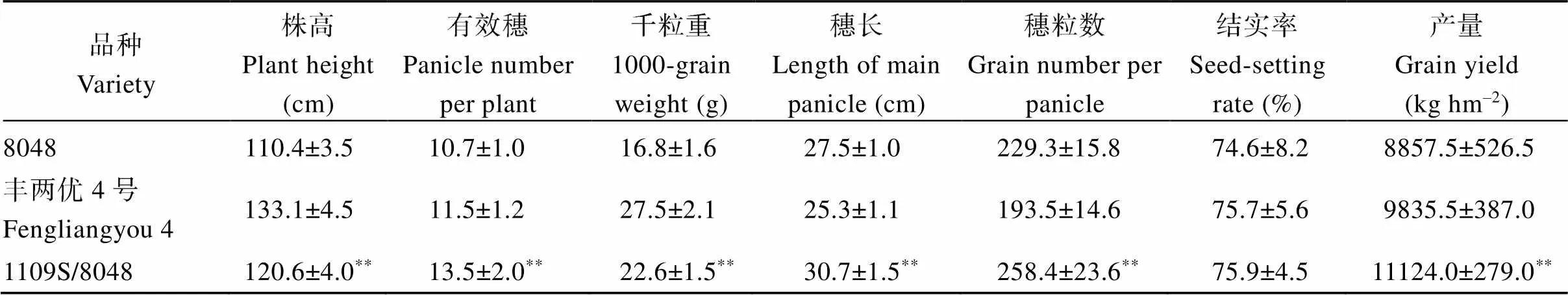
表2 杂交组合的农艺性状
数据表示平均值±标准误(= 20),**表示突变体与野生型之间存在显著差异(< 0.01)。
Data are presented as average values ± standard error (= 20).**, significant difference betweenmutant and wild-type (< 0.01).
杂种优势是指两个遗传基础有差异的亲本杂交产生的杂合子在生长势、生活力、生物量等方面优于两个亲本的现象。杂种优势现象在自然界普遍存在, 其最早于1717年被 Thomas Fairchild发现[18], Joseph Koelreuter于1761年利用[19]。我国是一个人口和农业大国, 粮食安全是关乎我国的长治久安和国计民生的重大战略性问题。杂交水稻的产量比常规水稻提高20%~30%, 在解决我国粮食短缺的问题上发挥了重要作用[1]。本研究通过CRISPR/Cas9技术敲除基因筛选到了一个低不育起点温度的温敏核不育系1109S, 并与优异父本8048配制了高产杂交稻组合1109S/8048。Zhou等[14]通过CRISPR/ Cas9基因编辑技术获得的温敏核不育系HNBS与华航1179配制的杂交稻组合的小区产量比父本增加14.4%, 而本研究获得的1109S/8048杂交稻组合的产量比父本8048增产25.6%, 同时比区试对照丰两优4号增产13.1%, 因此1109S/8048具有更高的超亲优势与杂种优势。
4 结论
利用CRISPR/Cas9基因编辑技术对6个粳稻和4个籼稻的基因进行敲除, 获得了粳稻和籼稻的温敏核不育系, 并比较了它们的不育起点温度。筛选到不育起点温度介于22.5~23.5°C之间的籼型温敏核不育系1109S, 并配制了高产杂交稻组合1109S/8048。不同遗传背景材料获得的突变体的不育起点温度不一样, 选取不同遗传背景的材料进行定点编辑可能获得不育起点温度较低的温敏核不育材料。
[1] 黄忠明, 周延彪, 唐晓丹, 赵新辉, 周在为, 符星学, 王凯, 史江伟, 李艳峰, 符辰建, 杨远柱. 基于CRISPR/Cas9技术的水稻温敏不育基因突变体的构建. 作物学报, 2018, 44: 844–851. Huang Z M, Zhou Y B, Tang X D, Zhao X H, Zhou Z W, Fu X X, Wang K, Shi J W, Li Y F, Fu C J, Yang Y Z. Construction ofmutants in rice based on CRISPR/Cas9 technology., 2018, 44: 844–851 (in Chinese with English abstract).
[2] 周延彪, 赵新辉, 唐晓丹, 周在为, 庄楚雄, 杨远柱. 基于CRISPR/Cas9技术的水稻反光敏不育基因突变体的获得. 杂交水稻, 2018, 33(6): 68–74. Zhou Y B, Zhao X H, Tang X D, Zhou Z W, Zhuang C X, Yang Y Z. Acquisition of mutants of the reverse photoperiod-sensitive genic male sterility genein rice based on CRISPR/Cas9 technology., 2018, 33(6): 68–74 (in Chinese with English abstract).
[3] Zhou H, Zhou M, Yang Y Z, Li J, Zhu L Y, Jiang D G, Dong J F, Liu Q J, Gu L F, Zhou L Y, Feng M J, Qin P, Hu X C, Song C L, Shi J F, Song X W, Ni E D, Wu X J, Deng Q Y, Liu Z L, Chen M S, Liu Y G, Cao X F, Zhuang C X. RNase ZS1processesUbmRNAs and controls thermosensitive genic male sterility in rice., 2014, 5: 1–9.
[4] 周海, 周明, 杨远柱, 曹晓风, 庄楚雄. RNase ZS1加工UbmRNA控制水稻温敏雄性核不育. 遗传, 2014, 36: 1274. Zhou H, Zhou M, Yang Y Z, Cao X F, Zhuang C X. RNase ZS1processes UbmRNA and controls thermosensitive genic male sterility in rice.(Beijing), 2014, 36: 1274 (in Chinese with English abstract).
[5] Belhaj K, Chaparro-Garcia A, Kamoun S, Patron N J, Nekrasov V. Editing plant genomes with CRISPR/Cas9., 2015, 32: 76–84.
[6] Baltes N J, Voytas D F. Enabling plant synthetic biology through genome engineering., 2015, 33: 120–131.
[7] Li M, Li X, Zhou Z, Wu P, Fang M, Pan X, Lin Q, Luo W, Wu G, Li H. Reassessment of the four yield-related genes,,andin rice using a CRISPR/Cas9 system., 2016, 7: 377.
[8] Sun Y, Jiao G, Liu Z, Zhang X, Li J, Guo X, Du W, Du J, Francis F, Zhao Y, Xia L. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes., 2017, 8: 298.
[9] Zhou Y B, Liu C, Tang D Y, Lu Y, Wang D, Yang Y Z, Gui J S, Zhao X Y, Li L G, Tang X D, Yu F, Li J L, Liu L L, Zhu Y H, Lin J Z, Liu X M. The receptor-like cytoplasmic kinase STRK1 phosphorylates and activates CatC, thereby regulating H2O2homeostasis and improving salt tolerance in rice., 2018, 30: 1100–1118.
[10] Wang H, Chu Z, Ma X, Li R, Liu Y. A high through-put protocol of plant genomic DNA preparation for PCR., 2013, 39: 1200–1205.
[11] 姜树坤, 张喜娟, 王嘉宇, 张凤鸣. 水稻幼穗-颖花发育的研究进展. 植物遗传资源学报, 2012, 13: 1018–1022. Jiang S K, Zhang X J, Wang J Y, Zhang F M. Research advancement on young panicle and spikelet development in rice (L.)., 2012, 13: 1018–1022 (in Chinese with English abstract).
[12] Zhou Y B, Liu H, Zhou X C, Yan Y Z, Du C Q, Li Y X, Liu D R, Zhang C S, Deng X L, Tang D Y, Zhao X Y, Zhu Y H, Lin J Z, Liu X M. Over-expression of a fungal NADP(H)-dependent glutamate dehydrogenaseGDH improves nitrogen assimilation and growth quality in rice., 2014, 34: 335–349.
[13] 李希陶, 刘耀光. 基因组编辑技术在水稻功能基因组和遗传改良中的应用. 生命科学, 2016, 28: 1243–1249. Li X T, Liu Y G. Genome editing technology for functional genomics and genetic improvement in rice., 2016, 28: 1243–1249 (in Chinese with English abstract).
[14] Zhou H, He M, Li J, Chen L, Huang Z F, Zheng S Y, Zhu L Y, Ni E D, Jiang D G, Zhao B R, Zhuang C X. Development of commercial thermo-sensitive genic male sterile rice accelerates hybrid rice breeding using the CRISPR/Cas9-mediatedediting system., 2016, 6: 1–12.
[15] Svitashev S, Schwartz C, Lenderts B, Young J K, Ciqan A M. Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes., 2016, 7: 13274.
[16] Wang Y P, Cheng X, Shan Q W, Zhang Y, Liu J X, Gao C X, Qiu J L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew., 2014, 32: 947–951.
[17] Jacobs T B, LaFayette P R, Schmitz R J, Parrott W A. Targeted genome modifications in soybean with CRISPR/Cas9., 2015, 15: 16.
[18] Zirkle C. Some forgotten records of hybridization and sex in plants 1716–1739., 1932, 23: 433–447.
[19] Stiles W. A short history of the plant sciences., 1942, 150: 672–673.
附图1 PCR鉴定T1代无选择标记基因的突变株
ZG75(A)、CYG(B)、YG0618(C)、ZG07(D)、T0361(E)、7679(F)、2537(G)、6150(H)、6379(I)、1109(J)非转基因植株的PCR鉴定。M: DNA marker; WT: 野生型; 1: 阳性对照; 2~6: GUS染色不呈蓝色的T1代植株。
Identification of the marker-free transgenic plants by PCR in ZG75(A), CYG(B), YG0618(C), ZG07(D), T0361(E), 7679(F), 2537(G), 6150(H), 6379(I), 1109(J). M: DNA marker; WT: wild-type; 1: positive control; 2–6: the leaf of T1plants did not show blue by GUS staining.
CRISPR/Cas9-mediated editing of the thermo-sensitive genic male-sterile genein rice
CHEN Ri-Rong1,**, ZHOU Yan-Biao2,3,**, WANG Dai-Jun5, ZHAO Xin-Hui3, TANG Xiao-Dan3, XU Shi-Chong4, TANG Qian-Ying3, FU Xing-Xue3, WANG Kai3, LIU Xuan-Ming1,*, and YANG Yuan-Zhu1,2,3,4,*
1Hunan Province Key Laboratory of Plant Functional Genomics and Developmental Regulation / College of Biology, Hunan University, Changsha 410082, Hunan, China;2Yuan Longping High-Tech Agriculture Co. Ltd. / Key Laboratory of Rice Germplasm Enhancement in Southern China, Ministry of Agriculture and Rural Affairs, Changsha 410001, Hunan, China;3Yahua Seeds Science Academy of Hunan, Changsha 410119, Hunan, China;4College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, Hubei, China;5College of Life Sciences, Hunan Normal University, Changsha 410081, Hunan, China
Thermo-sensitive genic male-sterile (TGMS) geneis most widely used in the two-line hybrid breeding system in China. To develop novel rice thermo-sensitive male sterile lines, we knocked out thegenes of six eliteand fourrice varieties by using CRISPR/Cas9 gene editing technology. By analyzing the critical sterility-inducing temperature (CIST) of the newly TGMS lines, it was found that the CIST ofTGMS lines ZG75S, CYGS, YG0618S, ZG07S, T0361S, and 7679S were between 28°C and 32°C, the CIST ofTGMS lines 2537S, 6150S and 6379S were between 24°C and 28°C, and the CIST ofTGMS line 1109S was lower than 23.5°C. These results indicated that the CIST ofmutant from different genetic background materials was different. The TGMS lines with lower CIST could be obtained by knocking out thefrom different genetic background materials. A hybrid rice combination 1109S / 8048 had high quality and high yield. The yield of 1109S/8048 was 13.1% higher than that of Fengliangyou 4. The creation of the TGMS 1109S and the high-yield cross combination 1109S/8048 provides a new way for high-yield breeding.
rice; CRISPR/Cas9; thermo-sensitive genic male sterile;;cross combination
10.3724/SP.J.1006.2020.92059
本研究由国家自然科学基金项目(31901516), 国家转基因生物新品种培育重大专项(2016ZX08001-004)和湖南省自然科学青年基金项目(2019JJ50414)资助。
This study was supported by the National Natural Science Foundation of China (31901516), the National Major Project for Developing New GM Crops (2016ZX08001-004), and the Project of Hunan Natural Science Youth Foundation (2019JJ50414).
杨远柱, E-mail: yzhuyah@163.com; 刘选明, E-mail: xml05@hnu.edu.cn
**同等贡献(Contributed equally to this work)
E-mail: 641605686@qq.com
2019-11-09;
2020-04-15;
2020-04-26.
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20200426.1442.012.html
猜你喜欢
哈哈画报(2022年8期)2022-11-23
亚热带农业研究(2022年1期)2022-08-08
蔬菜(2021年7期)2021-11-27
艺术启蒙(2020年6期)2020-07-16
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
小学生学习指导(低年级)(2018年10期)2018-10-13
儿童故事画报·发现号趣味百科(2017年4期)2017-06-30
文理导航·科普童话(2016年6期)2016-09-12
科技知识动漫(2016年4期)2016-04-20
