
基于多组学解析作物杂种优势机制及其利用展望
2020-07-09 02:57:12史关燕赵雄伟韩渊怀
山西农业大学学报(自然科学版) 2020年4期
史关燕,赵雄伟,韩渊怀
(1.山西省农业科学院经济作物研究所,山西汾阳032200;2.山西农业大学生命科学学院,山西太谷030801;3.山西农业大学农学院,山西太谷030801)
自然选择和人类选择促使了作物的不断驯化和生态适应性。随着测序技术不断完善和数量遗传学的发展,大量的优异变异位点被发掘。了解变异位点与表型的关系是实现作物高产、优质、抗逆等育种目标的重要措施之一。基于对杂种优势的基本认识而培育的杂交种已显著提高了作物产量和抗逆性,然而对其遗传、分子机制的研究却相对滞后。杂种优势是生物界的一种普遍现象,泛指遗传基础有差异的亲本杂交产生的杂交组合(尤其是杂种F1 代)在生长势和生物量表现出优于亲本的现象,已经广泛应用于作物(玉米、水稻、油菜等)农业生产上,为解决世界粮食问题、提高大众生活质量做出了重要贡献。近年来,关于作物品质育种的关注度大幅提升,利用杂种优势培育高产优质品种也取得了一定的进展,但缺乏机制基础研究,影响杂种优势利用范围的进一步提升和延伸。此外,高产优质育种过程中,由于品质性状的遗传复杂性,大量的后代品质性状鉴定的规模迅速增加,而且测定品质性状过程相对繁琐,需要结合生理生化实验方法,极大的限制优质品种培育的进度。因此,本文旨在综述近年来利用多组学技术解析作物杂种优势的分子遗传机制以及其对产量和其他农艺性状的杂种优势预测研究,为作物杂种优势遗传分子机制研究和优质品种培育提供借鉴。
杂种优势机制真正的探索始于上世纪九十年代。学术界解析杂种优势经典的遗传机理假说有3 种:第一种是“显性假说”(dominance hypothesis)[1],认为杂种优势在于亲本之一的显性等位基因(dominant alleles)抑制或互补了另一个亲本的不利或有害隐性等位基因(deleterious recessive alleles),杂种F1 中显性基因明显高于双亲,从而增强了杂合子代的生长势,随着后代的自交或近交,后代纯合体出现的几率不断增大,暴露出隐性基因所代表的有害性状,造成自交衰退。第二种假说是“超显性假说”(overdominance hypothesis),认为等位基因没有明显的优劣之分,而特定位点的杂合性在生理、生长以及生态适应性等方面表现明显优于纯合性类型[1];第三种假说是“上位假说”(epistasis hypothesis),认为来自父母本两个以上非等位基因之间的互作导致了杂种优势,某一个基因对性状的影响会受到其他一个或多个基因的影响,包括显性基因、加性基因、显性基因与加性基因之间的互作[2]。
从19 世纪开始,育种家利用遗传距离与杂种优势存在正相关的特点[3],从遗传背景差异大的种质中选择具有强杂种优势的亲本进行杂交组合,培育出了大批的优良品种,满足了市场需求。随着农作物种质资源的匮乏、优良种质资源和基因资源鉴定严重滞后,致使新杂交种选育时间加长,难以实现资源优势向经济优势的转化。尽管杂种中的全部基因资源来自双亲,但是杂种优势机制不是某一种假说可以解释的,杂种优势可能与物种、性状、亲本组合有关[4]。
1 杂种优势的遗传基础研究
分子数量遗传学采用分子标记技术定位控制表型变异的QTL,并对其遗传效应进行分析,使得从分子水平上解析杂种优势的遗传成为可能。杂种优势主要体现在有差异的亲本杂交产生的杂交组合后代的表型上。围绕杂交双亲具有一定的遗传差异,其亲本的遗传差异决定了杂种优势强弱特性,了解优异等位变异位点的遗传效应对指导作物杂种优势育种中选择亲本和有目的培育优良组合具有重要的实践意义。目前,利用分子数量遗传学方法进行农作物杂种优势研究的遗传群体设计主要有:通过NCIII 设计的F2 群体、三重测交设计群体、永久F2 群体以及基于NCIII 设计的回交群体等,以及多个群体组合探究杂种优势遗传效应(表1)。在不同作物的杂种优势遗传基础研究中,可能某个遗传效应对某个具体性状呈现主要作用,因而不同作物、不同遗传背景的材料和群体,得到的杂种优势遗传基础的解释并不唯一。此外,对水稻、棉花和玉米的杂种优势遗传学研究表明,自花授粉作物的杂种优势产生的原因较为复杂,可能由显性、超显性和上位性共同控制或其中的两种的互作效应作用,而异花授粉作物的杂种优势产生的重要原因可能是显性或超显性单个效应影响,其遗传基础相对较简单。
作物杂种优势农艺性状的QTL 定位结果表明,不同性状的杂种优势遗传基础不尽相同,经典的显性、超显性和上位性效应位点在不同程度上控制着农艺性状的杂种优势。在拟南芥中,Seymour 等[5]利用 30 个亲本以半双列杂交设计方案组配了435 个杂种,通过全基因组关联分析检测到的显著位点可以解释群体内20%的开花期和干重表型变异,并挖掘到了具有经典的显性和超显性效应位点,包括了与开花时间有关的MADS-box 转录 因 子 AGAMOUS-LIKE 50(AGL50)。 Yang等[6]利用200 个拟南芥杂交组合探索了生物量杂种优势的遗传基础,研究结果发现全基因组的超显性效应对生物量贡献不大,但显著的杂合位点遗传效应与生物量极显著相关,挖掘到的重要候选基因涵盖了细胞、代谢、发育及胁迫响应的生物功能途径,包括WUSCHEL、ARGOS 以及细胞周期相关基因。在水稻中,Chen 等[7]利用 4 个雄性不育系和96 个父本构建了384 个杂种F1 株系,对一般配合力(GCA)和特殊配合力(SCA)进行全基因组关联分析,研究结果共检测到34 个位点与农艺性状、GCA 和 SCA 显著关联,其中 Ghd8、GS3 和qSSR4 能解释30.03% 的籽粒产量GCA 表型变异。Huang 等[8]利用 17 个代表性水稻种质组配出了10 074 个杂种F2 株系,通过对产量性状进行QTL 定位及GWAS 分析,结果检测到控制杂种优势的主要基因位点 HD3a、TAC1、LAX1、Ghd8;进一步研究发现,在杂交配组中这些基因位点产生的单倍型组合,在杂交一代中高效地实现了对水稻花期、株型、产量各要素的理想搭配。在玉米中,Liu 等[9]对 3 个杂交 F2 群体的 5 360 个株系株高和产量等19 个农艺性状进行QTL 定位,研究结果共定位到628 遗传效应位点,其中多数位点以杂合基因型的完全与不完全显性、超显性效应贡献强杂种优势模式。在所定位到的QTL 位点中,Ky4q19 是一个影响玉米穗分枝数和每穗产量的杂种优势QTL 位点,该位点包含玉米主效基因ub3,与水稻株型产量基因OsSPL14(IPA1)的同源基因,将水稻基因OsSPL14 转入我国南方水稻后,其产量增加了10%左右。在油菜中,Jan 等[10]对950个杂交F2 株系开花和芥子油苷含量的GCA 进行全基因组关联分析,结果共检测到44 个SNP 与开花时间和芥子油苷含量GCA 显著关联,其中位于A02 染色体上含有开花基因FLOWERING LOCUS T(FT)和 FLOWERING LOCUS C(FLC)的2 个单倍型对开花时间具有明显的加性效应;而位于染色体A02 和C02 上的具有加性效应同源单倍型能显著降低芥子油苷含量。

表1 基于QTL定位分析作物杂种优势的遗传基础主要进展Table 1 The research progress on the genetic basis of crop heterosis based on the analysis of QTL mapping
综上所述,对不同作物、不同遗传背景的材料和群体的杂种优势遗传基础解释并不相同,不同性状杂种优势的遗传基础也不尽相同。因此,作物杂种优势形成的遗传机制十分复杂,其产生与遗传背景密切相关,受多个遗传位点控制,单一的模式并不能完全解释其遗传机制。随着植物性状自然遗传变异的深入研究和多组学技术的日益发展,有望能快速剖析目标性状杂种优势调控的分子遗传机制,为作物杂种优势的遗传研究和应用探索出一条新的思路。
2 基于多组学的杂种优势的分子机理研究
尽管作物杂种优势早在一百年前就开始应用于育种,且增产效果突出,但其机理仍然是一个悬而未决的复杂问题。进入21 世纪,随着“组学”技术的发展,对杂种优势的分子机制研究也相应地从转录组、表观组和蛋白质方面开展了探索,期望从基因的具体表现过程或结果更深入地认识杂种优势机制。
2.1 基于转录组解析植物杂种优势分子机理
目前,通过转录组学研究杂种优势分子遗传机制已取得了阶段性的进展。在水稻中,Wei 等[24]对超级稻LYP9 和亲本的叶片、穗部等7 个组织进行基因表达比较分析,发现F1 代差异基因数明显高于亲本,其中10.6%的差异表达基因在能量代谢和物质转运类别中有明显的富集,而且多数差异表达基因位于水稻中已知的产量相关QTL 中,如位于8 号染色体qRH8 对杂种优势的贡献源于杂种中功能性和非功能性的位点杂合性,非功能性位点RH8PA64S在编码区的有2 处插入和1 处缺失,缺失导致肽链提前终止并蛋白功能丧失 。在玉米上,Song 等[26]对正反杂交种 B73/Mo17 和Zheng58/Chang7-2 的穗位叶和茎高进行转录组分析,结果表明杂交种的促进衰老基因(ZmNYE1、ZmORE1、ZmWRKY53和ZmPIFs)表达量明显降低,而光合相关及赤霉素合成相关基因显著上调,从而促使了抽丝后穗位叶衰老延缓。此外,在玉米杂交种穗部发育的转录组分析结果也发现多数差异基因的表达具有明显的加性效应,杂交种和亲本之间的差异表达基因主要受顺式调控元件作用产生表达差异[27]。由此可见,在水稻和玉米等作物上利用转录组研究杂种优势的相关研究结果证明,杂种后代与亲本之间的基因差异表达有助于解析植物杂种优势形成的分子机理。
2.2 基于表观组解析植物杂种优势分子机理
杂种优势导致的基因表达差异也可能在于表观遗传变异。通过DNA 甲基化、组蛋白修饰和小分子RNA(sRNA)等层次调节植物基因表达,从而在植物体响应外界环境胁迫、自身生长发育和内在稳定基因组等方面发挥重要作用。植物DNA甲基化是目前表观遗传学研究的重要内容,在不改变DNA 序列的前提下,通过DNA 甲基化转移酶在基因组CpG 二核苷酸的胞嘧啶和腺嘌呤碱基上共价结合一个甲基基团,从而引起DNA 稳定性、DNA 构象及DNA 与蛋白质互作、染色质结构的改变,最终控制基因表达[28]。Wang 等[29]对杂种优势性状的稳定性研究发现,亲本基因组和表观基因组之间的相互作用对拟南芥杂种的活力起重要作用。在水稻中,Zhang 等[30]从“YY12”超级稻中鉴定出一个调控理想株型、增产并与降低DNA甲基化相关的QTL(qWS8/ipa1-2D),在最佳表达/剂量时可获得超级产量。Wang 等[31]对大豆4个亲本和12 个杂交种的12 个农艺性状与DNA 甲基化程度进行相关性分析,结果发现低度甲基化有助于增加节间数,过度甲基化有利于增加茎粗。此外,Shen 等[32]对拟南芥 Ler 和 C24 及其正反杂交F1 的全基因组DNA 甲基化水平研究结果表明,杂交种F1 甲基化水平比亲本显著增加,尤其在转座元件上发现了大量的甲基化;而且这些甲基化的位置主要发生在与亲本甲基化不同的区域和被sRNA 覆盖的区域。拟南芥和玉米上也同样发现杂种 F1 的 24-nt siRNA 比亲本的表达水平低[33,34]。总之,从表观学角度对杂种优势遗传基础进行探讨,虽然使得杂种优势机理的解析变得更为复杂,但是这也是阐释机理不可或缺的环节。
2.3 基于蛋白质组解析植物杂种优势分子机理
尽管在转录组水平上研究杂种优势促进了其机制的解析,但基因表达水平并不总能反映其蛋白质水平。因此,从蛋白质组水平上可以进一步探究杂种优势形成的机制。随着质谱鉴定技术、色谱分析与质谱联动技术及双向荧光差异凝胶电泳等相关技术的发展,蛋白质组学相关技术已经应用于杂种优势机制研究。Hu 等[27]利用质谱技术对玉米杂交种郑单909 及其亲本的幼穗进行蛋白组定量分析,结果发现在杂交种中的蛋白质表达模式主要是碳代谢和氮同化影响的显性表达模式。Xiang 等[35]在杂交超级稻 LYP9 种找到 11 个与亲本有显著差异的蛋白质,其中包括参与植物体中黄酮类化合物合成的苯丙氨酸解氨酶。由此可见,作物杂种优势现象的分子机制主要与生长和发育相关通路上的基因大量表达有关,从而提高了植物对环境的耐受性,并促使其他基因发挥最大遗传贡献。蛋白质技术分析蛋白差异表达在广度和精度上仍受技术因素的限制,因此高分辨率和高灵敏度蛋白质组学定量技术及其他相关技术的发展,将会有助于揭示杂种优势形成的分子机理。
目前,杂交种F1 代在转录组、表观组及蛋白组水平与亲本的差异研究均局限于某个器官或组织,尽管如此,基因组、转录组、蛋白质和表观组等各种组学技术及整合生物学的发展,将为全面解析杂种优势机制提供更加有力的技术支撑。
3 杂种优势相关重要基因与途径
挖掘杂种优势相关的重要基因一直是该领域研究热点和难点之一。目前对作物产量和其他农艺性状的杂种优势还主要集中在基于多组学挖掘相关优良位点和基因,但是被克隆到的重要基因少之又少。对优异等位基因的功能验证和分子调控网络构建是未来作物杂种优势研究的重点。
目前发现的杂种优势功能基因主要集中在与植物株型和开花时间相关性状上,如水稻Os-SPL14和玉米ub3,水稻Hd3a和 番茄SFT[8]。在籼稻和粳稻组合的杂交种中,OsSPL14或IPA1具有超显性效应,对杂优贡献较大。等位基因ipa1对穗粒数增加作用很大,而野生型等位基因IPA1却对结实率和穗数贡献大。杂交种的IPA1和ipa1基因序列位点变异在于其启动子区域,强超显性可能在于其杂合状态下的基因表达。此外,与水稻父本TAC1/TAC1相比杂合TAC1/tac1可显著促进穗粒数和千粒重,由于等位基因tac1 会减少分蘖数,导致杂合TAC1/tac1提高单株产量不显著,但是杂交种的分蘖数显著增加,促使了单位面积的产量显著提高[8]。在两系杂交水稻的杂种优势研究结果中发现LAX1和OsMADS51分别对粒重和抽穗期具有部分正显性效应[8];LAX1与水稻穗分支和末端小穗花芽分化有关[36],而Os-MADS51是 Ehd1、OsMADS14 和 Hd3a 上游促进短日照开花的调控因子[37]。类似于番茄中杂合SFT 对番茄产量具有超显性效应[38],具有杂优效应的水稻hd3a 等位基因有2 个碱基替换,与长日照延迟开花有关[8]。综上所述,杂种优势对作物产量和开花期的作用效应较为明显,可以探索利用这些等位基因作为设计育种标记,选择合适的亲本组配杂交种。
在转录组水平上分析杂种优势分子机制的结果中发现,基于防御系统相关的基因表达水平显著降低,植株的生长对环境产生“顿感现象”,促使大量有关植物生长和发育的基因过量表达并发挥其最大优势,最终导致杂交种的生长势和发育明显高于亲本。例如,最近发现某些杂交组合里因低水平水杨酸调控的基本防御相关基因表达降低可能与其杂种优势有关。杂交种的防御系统中水杨酸(SA)合成相关基因的表达也显著降低,而生长素(IAA)合成途径相关基因表达上调,这些上调的差异基因杂交种更多叶片细胞数量和较大叶片显著相关[39]。拟南芥C24 中过表达水杨酸水解酶(salicylate1 hydroxylase,NahG)可 以 导 致 C24 NahG 植株高大,类似杂种表型。拟南芥C24♀×Ler♂的F1 和C24 NahG 具有类似的防御和衰老相关的差异表达基因和代谢途径,包括水杨酸合成相关基因以及水杨酸响应基因表达水平的降低;生长发育与防御资源分配重要基因TL1 BINDING TRANSCRIPTION FACTOR1、衰 老相关转录因子WRKY53、NAC-CONTAINING PROTEIN29和ORESARA1表达水平在杂交种中都降低了 。杂种杨树与其亲本表现了极为显著的转录组差异,胁迫相关途径显著下调而代谢途径却多数上调[41]。此外,杂种优势相关的分子机制研究表明差异表达基因主要体现在光、激素及过氧化氢介导的信号通路、逆境响应及花发育等相关生物过程[42]。
4 多组学预测在植物杂种优势种应用
基因组预测是通过数量遗传统计模型计算标记位点的遗传效应值,预测特定性状的育种值,可以显著提高产量或农艺性状改良的育种效率。利用系谱和基因组信息预测杂优组合潜力的过程中,研发和演变了许多算法。在水稻中,Cui 等[43]利用NCIII 设计和水稻全基因组160 万个标记对1 495 个杂种的育种值进行杂交试验和预测准确性评估,研究结果发现使用最佳线性无偏预测(BLUP)对其表型的预测准确性要比基于多元回归的其他模型在标记数和样本量要求上更具优越性,其中对100 个杂交种的产量、粒数、千粒重、株高、粒长和粒宽预测准确性分别为0.54、0.62、0.54、0.58、0.92 和 0.87,此结果为水稻杂交种选育提供了更为有效的分子辅助育种方法。Kadam等[44]利用全基因组 2 296 个 SNP 和 BLUP 模型对玉米杂种F1 的产量和株高育种值预测,其预测准确性范围为0.28~0.91。用全基因组水平的SNP来估计个体育种值,不仅使得品种选育具有导向性,而且能够更好地将有利基因聚合在一起,比传统育种更加精确和高效。但基因组水平预测仍然只是通过连锁信息获取基因的状态特征,对复杂的基因互作(上位性)反映不足。
表观组、转录组和蛋白组是介于基因组和表型之间反映基因活性特征的重要环节,已有研究结果发现,多组学的结合能更好地提高杂种优势的准确性。Zenke-Philipp 等[45]基于转录组的岭回归(ridge regression)方法对98 个玉米杂交种籽粒产量、籽粒干物质含量的杂种表现和中亲优势进行预测,结果发现使用转录组预测杂种优势的准确性不仅与全基因组DNA 标记预测的趋势一致,而且利用转录组对玉米干物质产量性状方面的杂种优势预测的准确性也明显高于全基因组分子标记的基因组预测准确性[46]。
整合基因组与代谢组预测杂种优势也取得了较好的进展。例如,Riedelsheimer 等 利用全基因组56 110 个SNP 和130 个代谢组对玉米生物量的杂种优势预测准确性范围为0.72~0.81。在水稻上,遗传性较高的产量性状,代谢组预测的杂种优势准确性是基因组预测准确性的2 倍;基于杂交种的表型育种值,从21 945 组合中选择出10 个最佳杂交组合,其杂交种产量增产30%,快速有效地提高了育种效率[48]。此外,Schrag 等[49]比较了基因组、转录组(mRNA 和sRNA)、代谢组预测玉米籽粒产量和干物质含量的杂种优势准确性,研究结果发现用mRNA 预测产量杂种优势的准确性更高,基因组预测籽粒干物质杂种优势准确性比较高,而sRNA 对这2 个性状的杂优预测准确性较低。
5 作物杂种优势的利用及展望
杂种优势的优良遗传效应基因是在人工压力和自然选择下经过多次重组,种质资源间相互渗透形成了丰富的基因库,不同生态型种质遗传基础广泛,能适应较长期的育种目标。杂种优势机理首先在玉米上得到成功利用,并进一步扩展到更多的农作物上。目前,杂种优势已经广泛应用于玉米、水稻、油菜、高粱、大豆等粮食作物和多种蔬菜作物杂交种的推广上。我国杂粮的优势利用还存在一些挑战。例如,谷子是我国主要的杂粮作物之一,也是旱作生态农业不可替代的作物,具有抗旱、耐逆、杂种优势明显等特性。相比其他作物,谷子杂交种在生产上已有大规模的利用,但是由于不育系和常规种的遗传距离较近,缺少综合性状较好的不育系,谷子高产和优质相关性状的杂种优势还有大幅提升空间。理清谷子亲本遗传背景,拓宽遗传距离,不断改造和创新,丰富谷子种质资源,提升谷子杂种优势利用水平。此外,谷子杂交种制种效率较低,主要原因在于谷子花比较小,开放度小,开放时间较短,不育系柱头外露差,严重地限制了目标性状育种的进程。更重要的是,目前对于作物杂种优势的分子遗传机制研究主要集中于产量性状和相关的农艺性状方面,而随着而人们生活水平的提高,市场需求已经转向功能品质方面,品质已经成为杂粮消费者关注的热点。为此,充分发挥多组学技术优势,探索重要品质杂种优势形成机制,促进我国优异品质杂粮品种的培育是推动我国功能农业快速发展的重要契机之一。
杂种优势的形成是遗传、表观遗传和基因调控网络共同作用引起的一系列生物体内的复杂变化,未来应建立一个系统模型和研究平台对杂种优势遗传机制进行研究。目前,利用分子数量遗传学研究作物杂种优势大多仍停留在连锁分析和几个特定组合衍生的遗传分离群体上。目前的研究结果表明,利用连锁群体解析杂种优势的遗传机制效应具有一定的局限性。进行作物杂种优势的QTL 定位时,除了构建不同的遗传分离群体用于连锁分析外,还需用自然群体配制测交关联群体进行杂种优势位点的关联分析。连锁分析与关联分析相结合将会加快挖掘和验证杂种优势的QTL。此外,杂种优势的形成是一个非常复杂的生物现象,由基因、RNA 转录丰度和代谢环境相互作用使得作物杂种后代在生长、发育和分化等方面具有强于亲本,仅从基因组水平上研究目标性状的杂种优势不免带有一定的片面性。今后还需结合基因组、转录组和代谢组等多组学的杂种优势形成机理研究,从多个层次上综合分析,才有可能揭示作物杂种优势的分子遗传机理。
目前我国很多育种企业缺乏良好的杂种优势模式和清晰的种质资源体系,尤其缺乏完善的表型数据与基因型数据体系,这些不利因素限制了我国作物杂种优势利用。对作物目标性状的杂种优势分子遗传机制研究有助于进一步扩大杂种优势的应用范围,例如快速培育高产优势新品、基于功能基因改造的新种质创制等。
基于前期多组学解析杂种优势的分子遗传机制和基因组预测的应用,该研究提出了利用多组学拓展了作物杂种优势的应用范围。如图1 所示,首先基于大数据技术的“基因型到表型”全基因组预测策略;通过统计模型对基因组与转录组和代谢组进行全基因组预测,计算不同基因变异位点的育种值,构建育种位点效应值数据库。其次,通过育种值预测表型辅助育种家选系、配组合;选择遗传背景差异较大且优质等位变异聚合度较高优势父本和亲本进行分子设计育种,对杂交后代进行高产优质相关表型性状的鉴定,筛选高产优质目标性状的强优势亲本和组合,培育高产优质新品种。与此同时,结合数量遗传学和转录组学挖掘控制杂种优势效应的候选基因。最后,采用新型基因组技术、优势品种设计育种推动我国育种产业的发展,分子生物学的基因编辑技术探究控制杂种优势重要基因的分子生物学功能,创制符合育种新目标的种质,并构建优势等位基因数据库,推动我国作物的实现工程化育种进程。该育种策略期望在作物杂种优势分子遗传研究方面有效利用全基因组选择育种、分子设计育种,快速精准地筛选目标性状强优势候选组合和材料,大幅度降低田间测试的成本,降低田间的工作量,结合优异等位基因的功能验证研究,势必推动杂种优势的育种分子理论突破,尤其是杂种优势育种的应用范围。
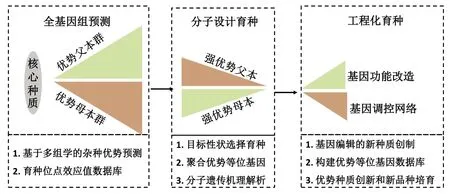
图1 基于多组学的作物杂种优势育种策略Fig.1 Breeding strategies for crop heterosis based on multi-omics approaches
猜你喜欢
中国糖料(2023年4期)2023-11-01 09:34:46
蔬菜(2021年7期)2021-11-27 03:45:25
今日农业(2021年14期)2021-11-25 23:57:29
四川蚕业(2020年2期)2020-07-10 03:14:48
中国果业信息(2019年11期)2019-01-05 20:47:24
科学导报(2018年47期)2018-05-14 12:06:10
西南农业学报(2016年5期)2016-05-17 05:42:23
山西农经(2016年5期)2016-02-28 14:24:37
现代农业(2015年5期)2015-02-28 18:40:52
新疆农垦科技(2014年10期)2014-02-28 19:21:08
