
低钙高镁人工脑脊液对大鼠脑片臂旁外侧核神经元温度敏感性的影响#
2020-06-30 08:10邓杰文胡红利杨培培黄成赵曦全俊如许莉张录顺张洁
四川生理科学杂志 2020年2期
邓杰文 胡红利 杨培培 黄成 赵曦 全俊如 许莉 张录顺 张洁△
(1.成都医学院基础医学院,四川 成都 610500;2.成都医学院临床医学院,四川 成都 610500;3.成都医学院检验医学院,四川 成都 610500)
根据温度敏感性是否依赖于突触传递,体温中枢视前区(Preoptic area,POA)的温敏神经元分为固有温敏神经元和条件温敏神经元两类。灌流低钙高镁人工脑脊液(Artificial cerebrospinal fluid,ACSF)阻断POA神经元突触传递后,大部分POA热敏神经元保留了温敏性,其温敏机制源于内在离子电导的改变,与突触传递没有关系,属于固有温敏神经元;与之相反,阻断突触传递后,所有POA冷敏神经元失去了冷敏性,其温度敏感性是突触传递形成,属于条件温敏神经元[1-2]。
臂旁外侧核(Lateral parabrachial nucleus,LPBN)能够将经脊髓传来的皮肤温度信号直接上传至POA区,参与了自主性和行为性体温调节[3-5]。前期研究发现,大鼠LPBN参与前馈体温调节的三个亚核(背亚核、中央亚核及外亚核)中均存在对局部脑温变化敏感的热敏神经元和冷敏神经元[6-8],但其温度敏感性是否依赖于突触传递尚不清楚。因此,本研究通过应用脑片膜片钳技术,灌流低钙高镁ACSF阻断突触传递,观察LPBN神经元胞外自发放电活动及温度敏感系数(m)值的变化,探讨LPBN神经元温度敏感性是否是内在固有特性,为后续进一步研究LPBN温敏神经元的温敏形成机制及其在温度信息整合中的作用打下基础。
1 材料与方法
1.1 材料与试剂
健康雄性SD大鼠,体重80~150 g,购自成都达硕生物医学科技有限公司。玻璃电极购自Sutter公司。HEPES、Na2GTP、葡萄糖酸钾、EGTA、MgATP购自Sigma公司。ACSF成分(mmol•L-1):NaCl 124,MgSO41.3,CaCl22.4,KCl 5,KH2PO41.24,NaHCO326,葡萄糖 10(渗透压 300 mOsm•kgH2O-1,pH 7.4)。低钙高镁ACSF成分(mmol•L-1):NaCl 119,MgSO411.4,CaCl20.2,KCl 5,KH2PO41.24,NaHCO326,葡萄糖 10(渗透压 300 mOsm•kgH2O-1,pH 7.4)。电极内液成分(mmol•L-1):葡萄糖酸钾 130,Na2GTP 2,MgATP 2,HEPES 10,CaCl21,EGTA 10(渗透压 290 mOsm•kgH2O-1,pH 7.2)。
1.2 LPBN大鼠脑薄片的制备
雄性SD大鼠以4%戊巴比妥钠腹腔注射麻醉后,迅速断头取脑,制备含LPBN的脑干块[6-8]。用502胶将脑干块粘贴于振动切片机标本盘内进行振动切片。当一月牙形透明区出现在脑桥头端背外侧缘,表明已达LPBN吻端,开始切取脑片。通常可切取2至3片含LPBN的脑薄片(300 μm厚)。
1.3 脑片膜片钳实验
按参照文献方法孵育脑片,并选取状态良好的LPBN神经元进行实验[6-8]。电极阻抗为4~7 MΩ,调整三维微推进器(MPC 200 Sutter Instruments,USA),使电极尖端与细胞膜表面接触,适当给予负压抽吸,形成细胞贴附式构型,然后转换至电流钳以记录LPBN神经元的胞外自发放电,放电信号经放大器(EPC-10,HEKA,Germany)放大后,通过Patchmaster软件进行信号采集,低通滤波为2 KHz,采样频率为10 KHz。
记录36℃~37℃范围内LPBN神经元基础放电活动后,在32℃~40℃范围内快速改变浴液温度,检测LPBN神经元自发放电频率的变化[6-8]。将放电频率和局部脑温之间的线性回归系数定义为温度敏感系数m值(Impulses),根据m值来确定LPBN神经元温度敏感性。m值≥0.8 imp·s-1·℃-1定义为热敏神经元,m值绝对值≤0.2 imp·s-1·℃-1定义为低等斜率的温度不敏感神经元,温度敏感系数m值>0.2 imp•s-1·℃-1和m值<0.8 imp•s-1·℃-1定义为中等斜率的温度不敏感神经元[6-8]。鉴定神经元温敏感性后,灌流低钙高镁ACSF10~15 min阻断突触传递,记录LPBN神经元自发放电活动和m值的变化,最后灌流常规ACSF洗脱10~15 min。
1.4 统计学方法
Patchmaster记录的原始数据经ABF Utility转换后,用Clampfit分析,计算突触阻断后各指标相对于突触阻断前的变化率。数据以均数±标准差(±SD)表示,采用SPSS21.0统计软件对数据进行配对样本t检验,P<0.05表示有显著性差异。
2 结果
2.1 低钙高镁ACSF对LPBN神经元自发放电活动的影响
表1显示35个LPBN自发放电神经元突触阻断后放电活动和温度敏感性的变化。灌流低钙高镁ACSF阻断突触传递后,热敏神经元(n=13)和温度不敏感神经元(n=22)的自发放电频率变化均无统计学意义(P>0.05),但中等斜率和低等斜率温度不敏感神经元放电频率都呈现出多样性改变,如中等斜率温度不敏感神经元(n=11)的放电频率7个加快,3个减慢,1个无变化;低等斜率温度不敏感神经元(n=11)的放电频率6个加快,3个减慢,2个无变化,见图1A。灌流低钙高镁ACSF阻断突触传递后,对三类神经元的放电幅度均无显著作用(P>0.05),见表1,图1B。
表1 突触阻断后不同类型LPBN神经元放电活动和温度敏感性的变化(±SD)
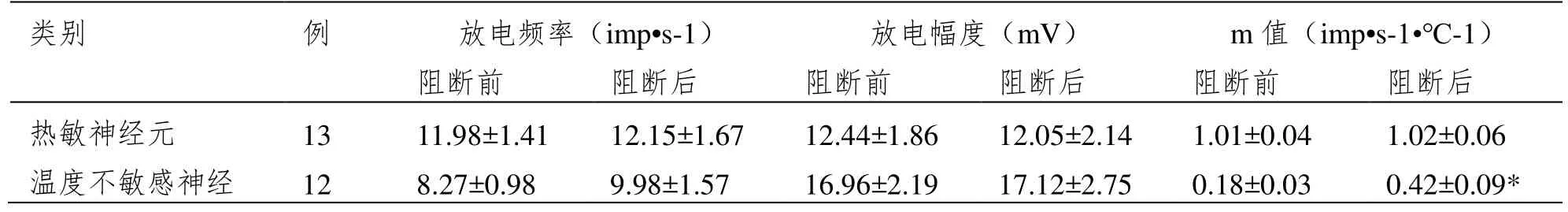
表1 突触阻断后不同类型LPBN神经元放电活动和温度敏感性的变化(±SD)
注:与突触阻断前比较,*P<0.05。
类别 例 放电频率(imp•s-1) 放电幅度(mV) m值(imp•s-1·℃-1)阻断前 阻断后 阻断前 阻断后 阻断前 阻断后热敏神经元 13 11.98±1.41 12.15±1.67 12.44±1.86 12.05±2.14 1.01±0.04 1.02±0.06温度不敏感神经 12 8.27±0.98 9.98±1.57 16.96±2.19 17.12±2.75 0.18±0.03 0.42±0.09*
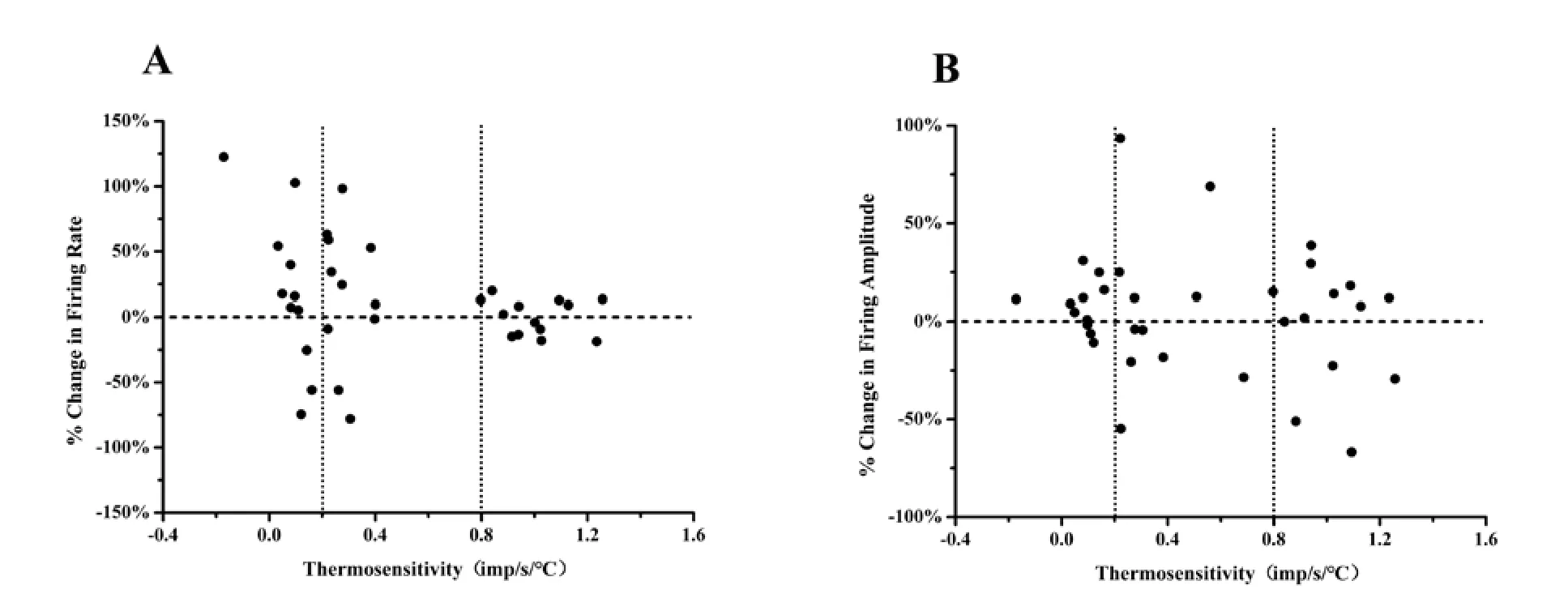
图1 每个LPBN低等斜率、中等斜率温度不敏感神经元和热敏神经元突触阻断前后放电频率(A)和放电幅度(B)的变化百分率
2.2 低钙高镁ACSF对LPBN神经元温度敏感性的影响
灌流低钙高镁ACSF阻断突触传递后,热敏神经元m值无明显变化(P>0.05),见表1、图2。图3所示是1例热敏神经元,突触阻断前m值为 1.09 imp•s-1·℃-1,突触阻断后的m值为0.87 imp•s-1·℃-1,洗脱后的m值为0.94 imp•s-1·℃-1,始终保持热敏性。突触阻断后温度不敏感神经元的m值变化显著(P<0.05),见表1。其中,64%(7/11)的中等斜率温度不敏感神经元m值变化率在70%以上;82%(9/11)低等斜率温度不敏感神经元的m值变化率在140%以上,见图2。图4所示低等斜率温度不敏感神经元,突触阻断前的m值为0.05 imp•s-1·℃-1,突触阻断后的m值增加到1.14 imp•s-1·℃-1,变成热敏神经元,洗脱后,其m值又恢复为0.05 imp•s-1·℃-1,失去热敏性。在22例温度不敏感神经元中,有4例出现相似的变化,突触阻断后,温度敏感系数增加非常明显,变成了热敏神经元,洗脱后又恢复为温度不敏感神经元,这4例温度不敏感神经元的放电频率明显增加(P<0.01),突触阻断前后的放电频率分别是11.85±5.72 imp•s-1和17.00±5.35 imp•s-1(n=4)。
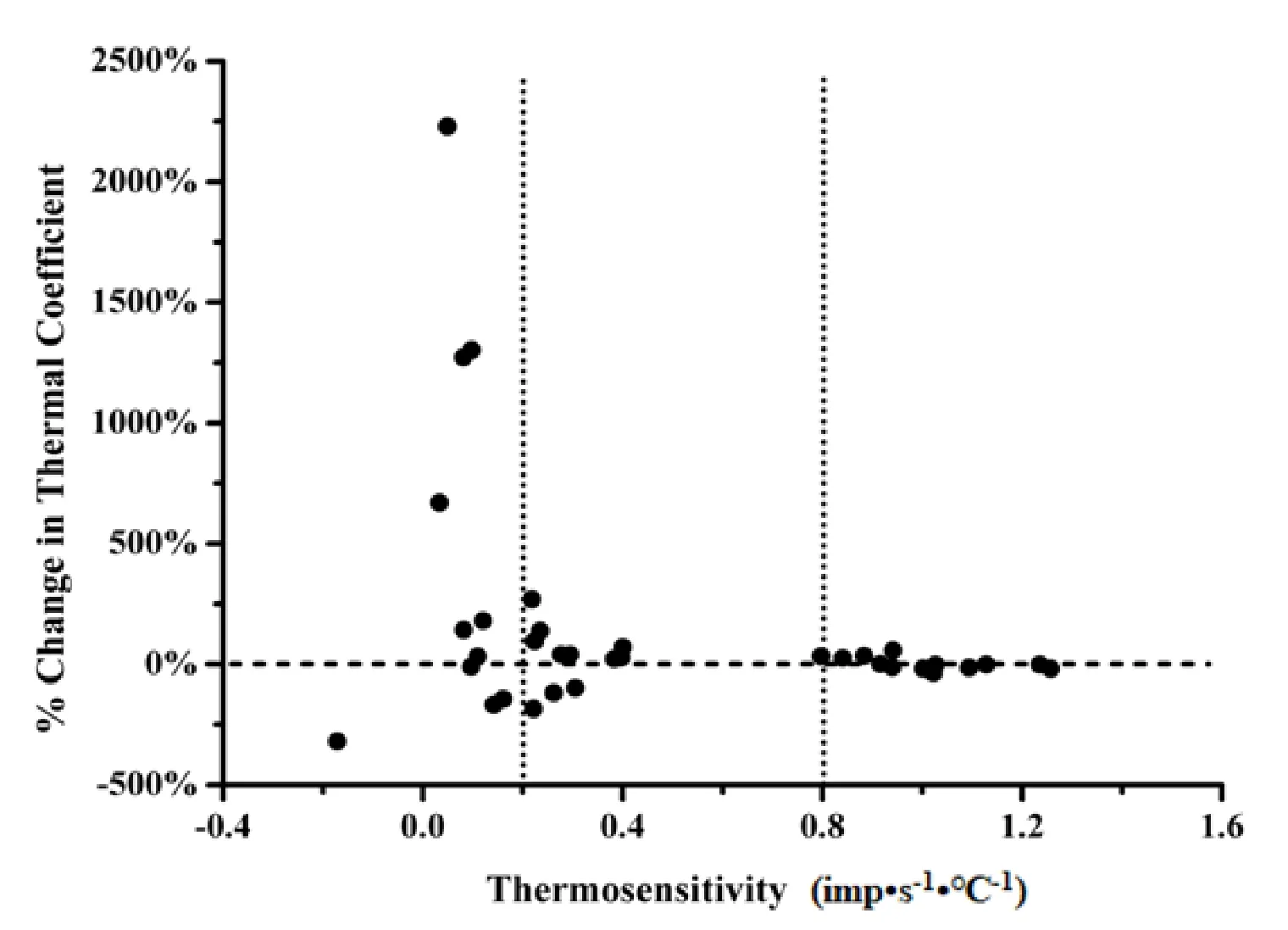
图2 每个LPBN低等斜率、中等斜率温度不敏感神经元和热敏神经元突触阻断前后m值的变化百分率
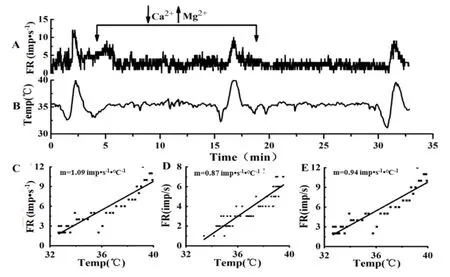
图3 突触阻断前、后和洗脱后LPBN热敏神经元自发放电频率和温度敏感性的变化
3 讨论
LPBN能够将经脊髓传来的皮肤温度信号直接上传至POA区,引起前馈体温调节反应,以维持体温稳定[3-5]。采用脑片膜片钳技术,我们在LPBN参与前馈体温调节的三个亚核中都发现对局部脑温变化敏感的热敏和冷敏神经元[6-8]。在极端温度下,当前馈体温调节已不能维持体温稳定,或在运动、摄入冷流体食物、出血等导致体温的内稳态被破坏时,分布在POA、LPBN等核团的温度敏感神经元可监测局部脑温变化,以负反馈体温调节的方式来维持体温稳定[6]。
LPBN除了接受皮肤前馈温度信号外,还能感受局部脑温的变化。,但温敏形成机制尚有待研究。在体温中枢POA区,灌流低钙高镁ACSF阻断突触传递后,几乎所有热敏神经元保留了热敏性;与之相反,突触阻断后,所有冷敏神经元失去了冷敏性,表明POA神经元热敏性是其内在特性,冷敏性则是突触传递形成[1-2]。本实验采用相似的方法技术,分析LPBN神经元的温度敏感性是否依赖于局部突触网络,为后续进一步探讨其温敏形成机制及其在温度信息整合中的作用打下基础。
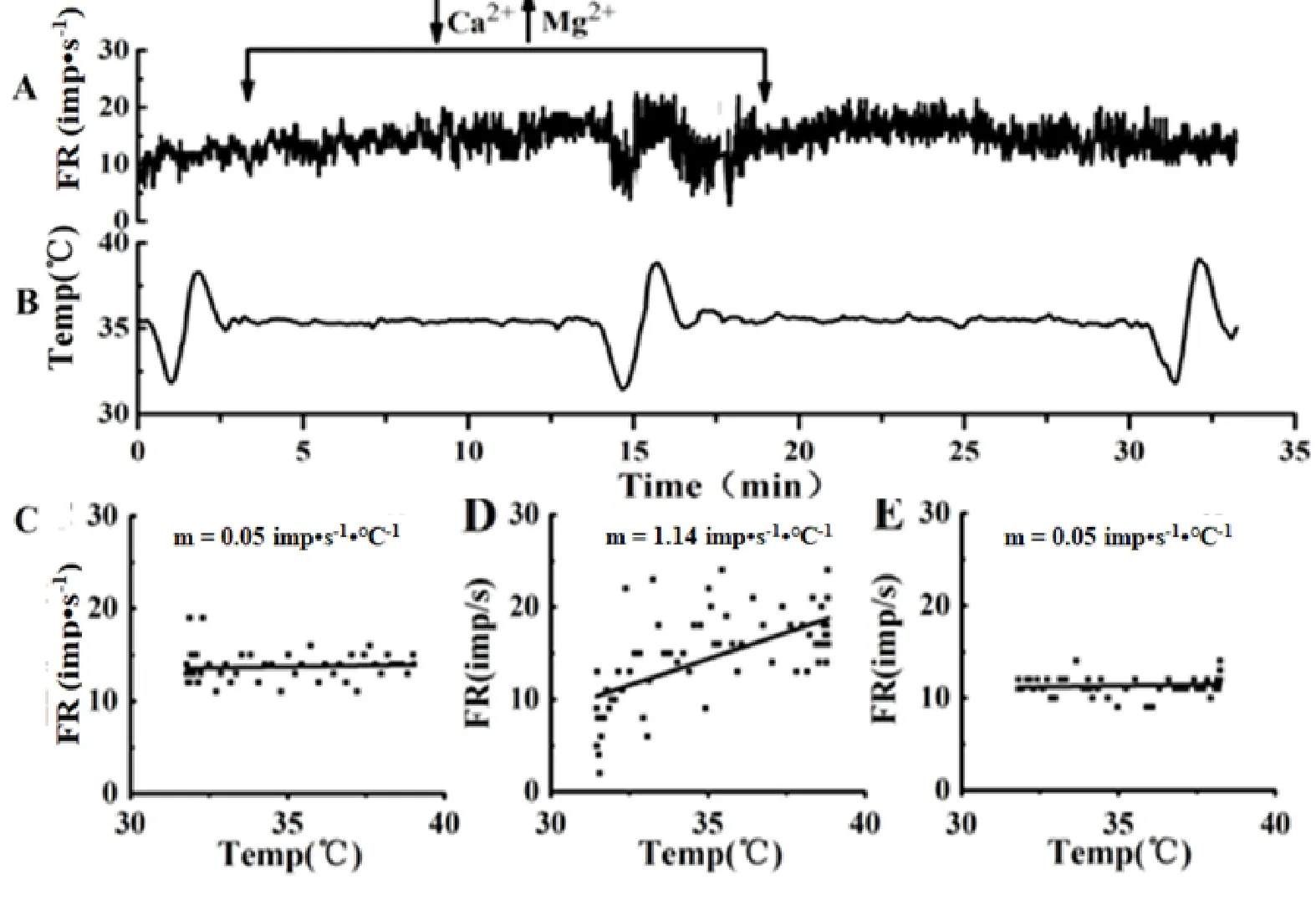
图4 突触阻断前、后和洗脱后LPBN温度不敏感神经元自发放电频率和温度敏感性的变化
我们在LPBN离体脑片灌流低钙高镁ACSF 10~15 min阻断突触活动后发现, LPBN热敏神经元在灌流低钙高镁ACSF后,放电频率和幅度并无显著改变,其温度敏感性在突触阻断前、后和洗脱后,也无明显的变化。说明和POA区相似,LPBN热敏神经元的温敏特性是其内在特性,而不是突触传递形成,属于固有温敏神经元,其热敏形成可能与其内在离子电导的温度依赖性变化有关[1-2,9]。突触阻断后,热敏神经元的放电频率和温度敏感性几乎都没有发生变化。提示,LPBN局部的许多突触联系并未投射到热敏神经元,热敏神经元很有可能更多接受的是LPBN以外的突触联系并在此发生信息整合,如脊髓传来的皮肤温度信息,孤束核传来的内脏温度信息,都有可能会聚到这些热敏神经元并与局部温度变化信息发生整合[3]。因此,LPBN极有可能与POA和脊髓相似,既能接受外周传来的温度信息,也能感受局部温度变化,并将传入的和局部的温度信息整合后上传到体温中枢POA区,进一步引起体温调节反应。
灌流低钙高镁ACSF后,LPBN温度不敏感神经元的放电频率呈多样性改变,幅度无明显变化,m值在突触阻断后大多增加。有4例温度不敏感神经元在突触阻断后变成了热敏神经元,洗脱后又恢复为温度不敏感神经元。说明这类温度不敏感神经元本身可能具有热敏性,但其热敏性被相邻温敏神经元的突触活动所抑制。例如,与相邻热敏神经元形成抑制性突触或与冷敏神经元形成兴奋性突触都可能导致其热敏性被抑制。根据这些神经元突触阻断后放电频率增加的现象,我们推测这些温度不敏感神经元极有可能接受了相邻热敏神经元的抑制性突触输入,导致其热敏性被抑制,呈现出温度不敏感性。但这尚需通过观察温度变化对温度不敏感神经元抑制性突触活动的影响来进一步验证。
LPBN功能复杂,除了参与体温调节,还与代谢[10]、心血管[11]等活动的调节和痛、痒觉等信息的传递有关[12]。突触阻断后,LPBN温度不敏感神经元的放电频率呈多样性改变。说明许多LPBN局部突触联系投射到温度不敏感神经元,并形成了复杂的局部突触网络,这些温度不敏感神经元虽然不能感受局部脑温的变化,但可能参与了其他与体温调节有关内稳态的调控,如代谢、心血管活动的调节,突触阻断后的多样性反应,也可能与这些神经元参与多种功能调节有关。下一步需进一步研究其他内稳态系统反馈信号如葡萄糖、血压对温度敏感及不敏感的神经元活动的影响。
综上所述,LPBN热敏神经元属于固有温敏神经元,其温敏特性不是突触传递形成,而是其内在特性,可能与其内在离子电导的温度依赖性变化有关。
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
电气技术(2022年6期)2022-06-27
保健医苑(2021年4期)2021-12-01
载人航天(2021年5期)2021-11-20
现代仪器与医疗(2021年1期)2021-06-09
现代临床医学(2021年1期)2021-01-26
甘肃教育(2020年4期)2020-09-11
语数外学习·高中版上旬(2020年8期)2020-09-10
福建中学数学(2016年7期)2016-12-03
医学信息(2015年5期)2015-03-31
