
拟穴青蟹Na+/H+-exchanger基因克隆及其在盐度胁迫下的表达分析
2020-06-10 01:30张凤英马春艳马凌波乔振国蒋科技
海洋渔业 2020年2期
刁 乐,张凤英,宋 炜,马春艳,王 伟,谌 微,马凌波,赵 明,乔振国,蒋科技
(1. 中国水产科学研究院东海水产研究所,农业农村部远洋与极地渔业创新重点实验室,上海 200090; 2. 上海海洋大学水产与生命学院,上海 201306)
盐度对水生甲壳动物的生长发育和新陈代谢等具有极其重要的影响[1-2],水生甲壳动物Na+/H+-exchanger向膜内和膜外分别转运Na+和H+[3-5],维持胞内Na+稳态,对广盐性蟹类在低盐环境下的Na+摄入具有重要影响[6-11]。拟穴青蟹(Scyllaparamamosain)隶属于节肢动物门(Arthropoda) ,甲壳纲(Crustacea) ,十足目(Decapoda),短尾亚目(Brachyura) ,梭子蟹科(Portunidae),青蟹属,广泛分布于我国东南沿海地区[12-13],是我国重要的海水捕捞和养殖品种。拟穴青蟹幼体在27~35盐度条件下都能正常发育成仔蟹,但在盐度27时的成活率和生长情况最好[14-16]。随着个体的生长发育,拟穴青蟹具有逐步趋向低盐环境生活的习性[17-18]。拟穴青蟹幼体相比仔蟹更易遭受盐度改变带来的影响[17],幼体培育阶段的盐度环境往往直接影响育苗的成活率,其中大眼幼体变态期是幼体培育过程中死亡率最高的阶段[17-18]。当前海水经济蟹类中,拟穴青蟹苗种规模化繁育技术还未完全成熟,其中一个关键因素就是目前拟穴青蟹幼体阶段盐度胁迫的研究很少,缺乏相应的调控思路与方法。
水生甲壳动物中,三疣梭子蟹(Portunustrituberculatus)[5]、普通滨蟹(Carcinusmaenas)[19]、中国明对虾(Penaeuschinensis),麦龙螯虾(Cheraxtenuimanu)[20]、天空蓝魔(Cheraxdestructor)[20]和红螯螯虾(Cheraxquadricarinatus)[21]等已见Na+/H+-exchanger基因的相关报道。马金武等[5]研究发现,Na+/H+-exchanger基因主要在三疣梭子蟹的鳃组织中特异性表达,在低盐中起作用,在高盐中的作用不明显。XU等[22]报道了拟穴青蟹Na+/H+-exchanger的部分序列,但未报道Na+/H+-exchanger的表达情况。本研究克隆获得拟穴青蟹Na+/H+-exchanger的cDNA全长,并对该基因及其编码的氨基酸序列进行生物信息学分析,分析该基因在不同组织和不同幼体时期的表达变化,同时研究拟穴青蟹大眼幼体时期盐度胁迫下Na+/H+-exchanger的表达情况。以期为广盐性甲壳动物Na+/H+-exchanger基因渗透调节机理的研究、拟穴青蟹耐低盐新品系的选育等提供参考。
1 材料与方法
1.1 实验材料
拟穴青蟹幼体取自海南琼海东海水产研究所养殖基地,不同幼体分期为受精卵(F.egg)、溞状幼体I~V(Z1~Z5)、大眼幼体(M)、一期仔蟹(C1)和二期仔蟹(C2),加无液氮型样品RNA保存液(生工生物工程有限公司)冻至-80℃保存备用。成体购买于中国海南省文昌市环球码头,实验前在实验室水循环系统提前养殖一周。
1.2 拟穴青蟹Na+/H+-exchanger基因cDNA全长克隆
前期通过转录组测序获得了拟穴青蟹Na+/H+-exchanger的核心序列,提取成体雌蟹的神经节组织构建5′-RACE和3′-RACE cDNA文库,在预测的ORF区设计RACE特异性引物,进行5′-RACE和3′-RACE PCR扩增反应得到该基因全长cDNA序列。RACE使用Clontech公司(TAKARA,中国大连)的SmartTM Race cDNA Amplification kit试剂盒,按照使用说明书进行操作。5′-RACE 的特异性引物为Sp-NHE-F(AGACCACCAGATACCACCGA),3′-RACE的特异性引物为Sp-NHE-R(ACCTCCAATCCCCAAAC
CTTAC),另一方向引物为试剂盒自带通用引物。
RACE产物通过琼脂糖凝胶电泳进行检测,将目的条带进行割胶回收,克隆到pMD19-T载体上转化到大肠杆菌DH5α感受态细胞中,挑取阳性克隆送至上海杰李生物公司进行测序。测序结果利用DNAMAN软件进行拼接获得Na+/H+-exchanger的cDNA全长。除试剂盒提供的引物外,其他引物委托上海杰李生物技术有限公司合成。
1.3 生物信息学分析
使用ORF Finder(http ://www.ncbi.nlm.nih.gov/gorf/gorf.html)查找序列的开放阅读框并翻译,在Nr 数据库进行blast比对分析从而对基因进行初步注释。利用ExPASy(http://au.expasy.org/drug_design)分析蛋白质的分子量和等电点。
在UniPortKB数据库中下载多个物种的Na+/H+-exchanger序列,所使用的序列包括:三疣梭子蟹A0A286QYA6、普通滨蟹Q23706、中国明对虾A0A286QYA6、麦龙螯虾A0A0C5DFF2、天空蓝魔A0A0C5DNU5、红螯螯虾A0A0C5DNU5、黑腹果蝇(Drosophilamelanogaster)Q8MQP4、埃及伊蚊 (Aedesaegypti)Q7Z0L7、中华按蚊(Anophelessinensis)A0A084VYE0、冈比亚按蚊(Anophelesgambiae)Q5TNB1、尼罗罗非鱼(Oreochromisniloticus)I3J5H3、斑马鱼(Daniorerio)F1R610、雀鳝(Lepisosteusoculatus)W5MNT3、智人(Homosapiens)Q14940、小白鼠(Musmusculus) B2RXE2。
使用 DNAMAN 软件对部分物种的Na+/H+-exchanger氨基酸序列进行多重比对分析,使用MEGA7的clustalW对16条Na+/H+-exchanger序列进行比对,邻接法构建发育树,Bootstrap设置为1 000。采用ORF Finder进行基因开放阅读框(ORF)预测,采用Blast程序分析目的基因与其他物种的相似度,使用DNAMAN软件进行基因编码氨基酸序列的多重序列比对。使用ProtParam tool、SMART、TMHMM Server v. 2.0和ProtScale等在线生物信息分析工具对基因编码蛋白的基本物理性质、结构域、信号肽、跨膜结构和亲、疏水性进行预测分析。
1.4 Na+/H+-exchanger基因的组织表达分析和时空分布分析
提取成体蟹的脑神经节、鳃、心脏、中肠、卵巢(卵巢发育I期)、纳精囊、精巢、胸神经节组织,以及从受精卵到仔蟹Ⅱ期不同幼体发育时期样本的总RNA,使用DNAase I处理后反转录用于Na+/H+-exchanger的表达分析。RNA提取使用北京艾德莱生物技术有限公司的总RNA提取试剂盒,反转录使用TOYOBO的荧光定量反转录试剂盒参考说明书进行。荧光定量使用试剂为TAKARA公司的SYBR green PCR master mix(TAKARA,大连)。反应体系和程序参考本实验室常规方法进行[23],反应体系为:cDNA模板1 μL,SYBR Primix ExTaq5 μL,上下游引物各0.5 μL,ddH2O 3 μL,PCR反应程序为:95 ℃ 30 s; 95 ℃ 5 s, 60 ℃ 34 s, 40 个循环; 95 ℃ 15 s, 60 ℃ 1 min, 95 ℃ 15 s。
由于18S rRNA在拟穴青蟹的多种不同样本中能稳定表达,适合作为拟穴青蟹基因表达分析的内参基因。本文使用18S作为内参基因[24],Na+/H+-exchanger的特异性引物为Sp-NHE-RTF(TGTGATGTGTTACGGAGGGC)和Sp-NHE-RTR(AAACCATGGCGATAGTGGCA)。采用标准曲线法进行分析计算检测Na+/H+-exchanger基因在各个样本中的相对表达量。
1.5 拟穴青蟹大眼幼体在不同盐度胁迫下对Na+/H+-exchanger的表达调控分析
实验所用为拟穴青蟹大眼幼体3日龄,实验设置盐度7、17和37试验组和自然海水对照组(盐度27),保持温度与养殖池温度一致,每组约40只幼体,盐度37试验组水体由自然海水与卤水混匀调制,盐度7和17试验组水体由自然海水与灭菌处理后的自来水(灭菌处理程序与养殖池海水处理程序一致)混匀调制,实验前水体充分曝气。各盐度组分别在实验20 min、1 h、2 h和6 h时取样,放于无RNA酶离心管中,标记编号后置于RNA保存液中冷冻保存。
2 结果与分析
2.1 Na+/H+-exchanger基因cDNA全长克隆及序列分析
本研究采用RACE技术克隆获得了拟穴青蟹Na+/H+-exchanger的全长(GenBank 接收号:MK408672),cDNA全长3 624 bp(图1),开放阅读框(ORF)长2 886 bp。ORF编码氨基酸961个,预测蛋白分子量为107.1 kDa,理论等电点为6.90。
通过氨基酸序列多重比对分析表明,Na+/H+-exchanger氨基酸序列在60~660氨基酸区段内保守性较高。SMART分析预测表明该基因编码蛋白结构域包括: Na+/H+-exchanger蛋白结构域(82~484)、信号肽 (0~21)和4个低复杂结构(23~42、 675~684、855~868和889~902)。TMHMM和ProtScale在线工具分析表明,Na+/H+-exchanger基因编码蛋白质结构中含有12个跨膜结构域,主要分布于0~487氨基酸区段,该区段内亲、疏水性氨基酸均有分布,488~951氨基酸编码蛋白无跨膜区段,位于膜内,呈亲水性。
2.2 Na+/H+-exchanger基因的多序列比对和系统发育分析
多序列比对分析发现,拟穴青蟹Na+/H+-exchanger基因编码氨基酸序列与三疣梭子蟹、中国明对虾、普通滨蟹、麦龙螯虾、天空蓝魔和红螯螯虾的一致度分别为84.9%、62.7%、56.5%、53.7%、53.5%和52.1%(图2)。利用MEGA 4.0软件构建系统发育进化树分析表明,脊椎动物和无脊椎动物Na+/H+-exchanger遗传进化距离较远,埃及伊蚊等昆虫类聚为一支,拟穴青蟹同三疣梭子蟹亲缘关系最近,并同中国明对虾等聚为一支(图3)。
2.3 Na+/H+-exchanger基因时空表达分析
利用qRT-PCR检测Na+/H+-exchanger基因在拟穴青蟹成蟹不同组织中的表达情况。组织分布分析表明,拟穴青蟹Na+/H+-exchanger在卵巢中表达量最高,其次是精巢、鳃、中肠、胸神经节、心脏、纳精囊,脑神经节中表达量最低(图4)。除性腺组织外,其他组织表达情况与三疣梭子蟹和普通滨蟹在Na+/H+-exchanger基因上的研究结果基本一致。在幼体发育过程中,Na+/H+-exchanger基因在受精卵时期表达量最高,大眼幼体期次之。溞状幼体Ⅱ期(Z2)最低,从受精卵期到溞状幼体Ⅱ期(Z2)逐渐降到最低。然后随着发育逐渐升高,到大眼幼体期(M)再次达到较高值,之后逐渐下降(图5)。


图1 拟穴青蟹 Na+/H+-exchanger的cDNA全长及其翻译的氨基酸序列Fig.1 cDNA and deduced amino acid sequence of Na+/H+-exchanger from Scylla paramamosain注:方框内为起始密码子(ATG)和终止密码子(TAG);* 代表终止密码子;黄色阴影所示为荧光定量引物结合位点;阴影部分分别是信号肽(0~21)和Na+/H+-exchanger蛋白结构域(82~484)Note: The starting codon(ATG) and the stop codon (TAG) are in the box; * stands for termination codon; the yellow shadow regions are the binding sites of fluorescence quantitative primers; the shadow regions are signal peptide(0-21) and Na+/H+-exchanger domain(82-484)

图2 Na+/H+-exchanger氨基酸序列与其他物种Na+/H+-exchanger的序列比对Fig 2 Na+/H+-exchanger amino acid sequence compared with Na+/H+-exchanger of other species注:黑色下划线标注为预测的Na+/H+-exchanger蛋白结构域Note: Na+/H+-exchanger domain is underlined

图3 基于Na+/H+-exchanger氨基酸序列的物种NJ系统发育树Fig 3 Phylogram based on neighbor-joining tree of species of Na+/H+-exchanger amino acid sequences

图4 Na+/H+-exchanger基因在不同组织中的表达Fig 4 Tissue distribution analysis of Na+/H+-exchanger注: Cg:脑神经节;Gi:鳃;H:心脏;Mi:中肠;Ov:卵巢;Sr:纳精囊;Te:精巢;Tg:胸神经节Note: Cg: cephalic ganglion;Gi: gill;H: heart; Mi: midgut;Ov: ovary;Sr: seminal receptacle;Te: testes;Tg: thoracic ganglion
2.4 Na+/H+-exchanger基因在不同盐度胁迫下的表达分析
盐度7、17和37试验组和养殖塘自然海水对照组(盐度27)的测定结果表明,盐度从27骤降至7和17时,7和17试验组Na+/H+-exchanger基因的表达水平明显高于对照组。在1 h和2 h期间,盐度7和17试验组Na+/H+-exchanger基因的表达水平明显下降,此时已有足够的蛋白产物,推测这是此时其表达水平降低的原因;而6 h时,盐度7和17试验组Na+/H+-exchanger基因的表达水平基本恢复正常,说明幼体已适应这种盐度环境。盐度37试验组在20 min时Na+/H+-exchanger基因先下降,在1~2 h期间逐渐恢复至正常表达水平,但到6 h时其表达水平再次下降(图6)。
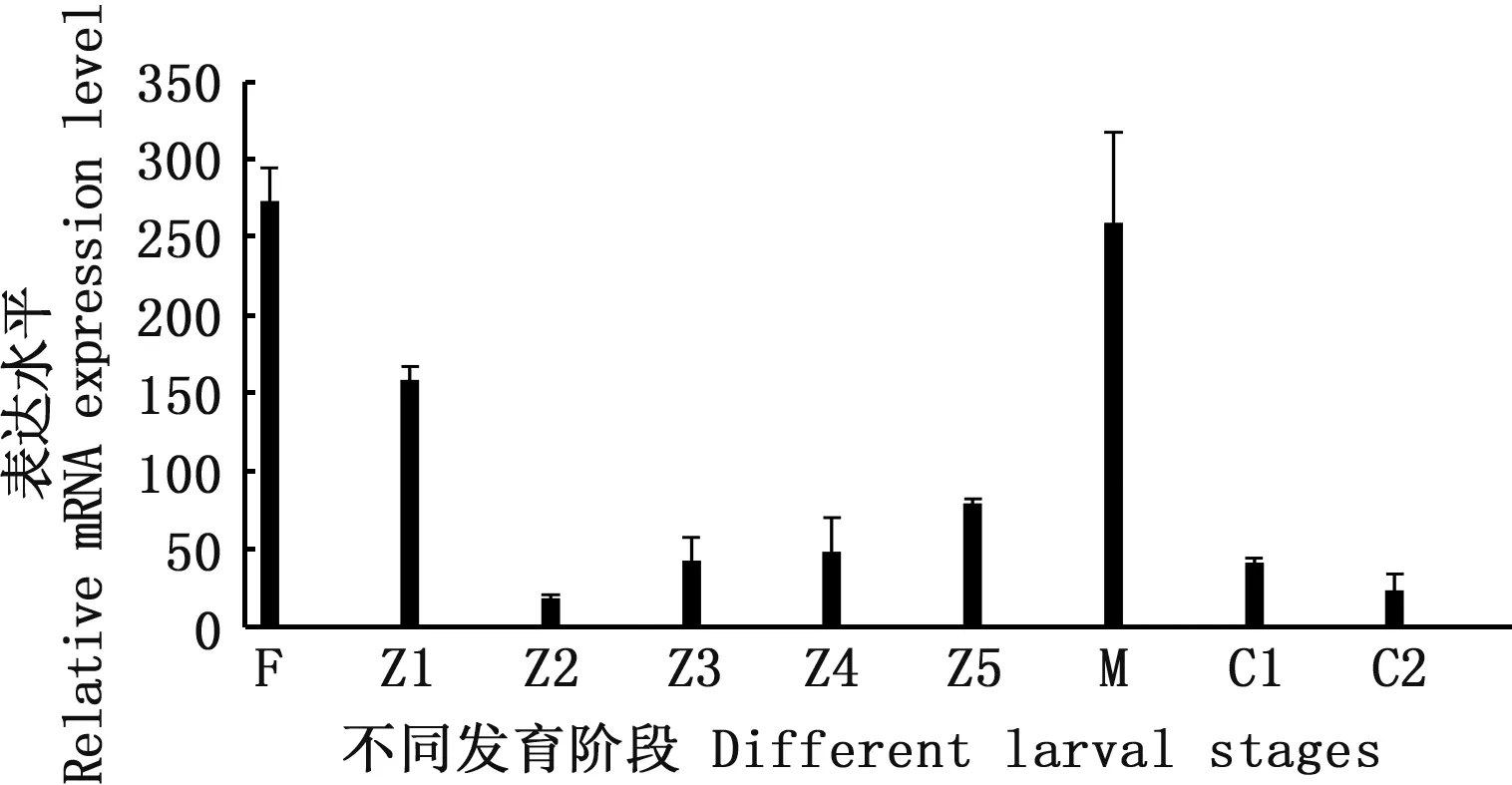
图5 Na+/H+-exchanger基因在不同发育阶段中的表达Fig 5 Expression pattern of Na+/H+-exchanger during different larval development stages注: F. egg: 受精卵;Z1~Z5:溞状幼体I-V;M: 大眼幼体;C1:一期仔蟹;C2:二期仔蟹Note: F. egg: fertilized eggs;Z1-Z5: five developmental phases of the zoea stage;M: megalopa stage;C1: the early juvenile crab;C2: the second juvenile crab
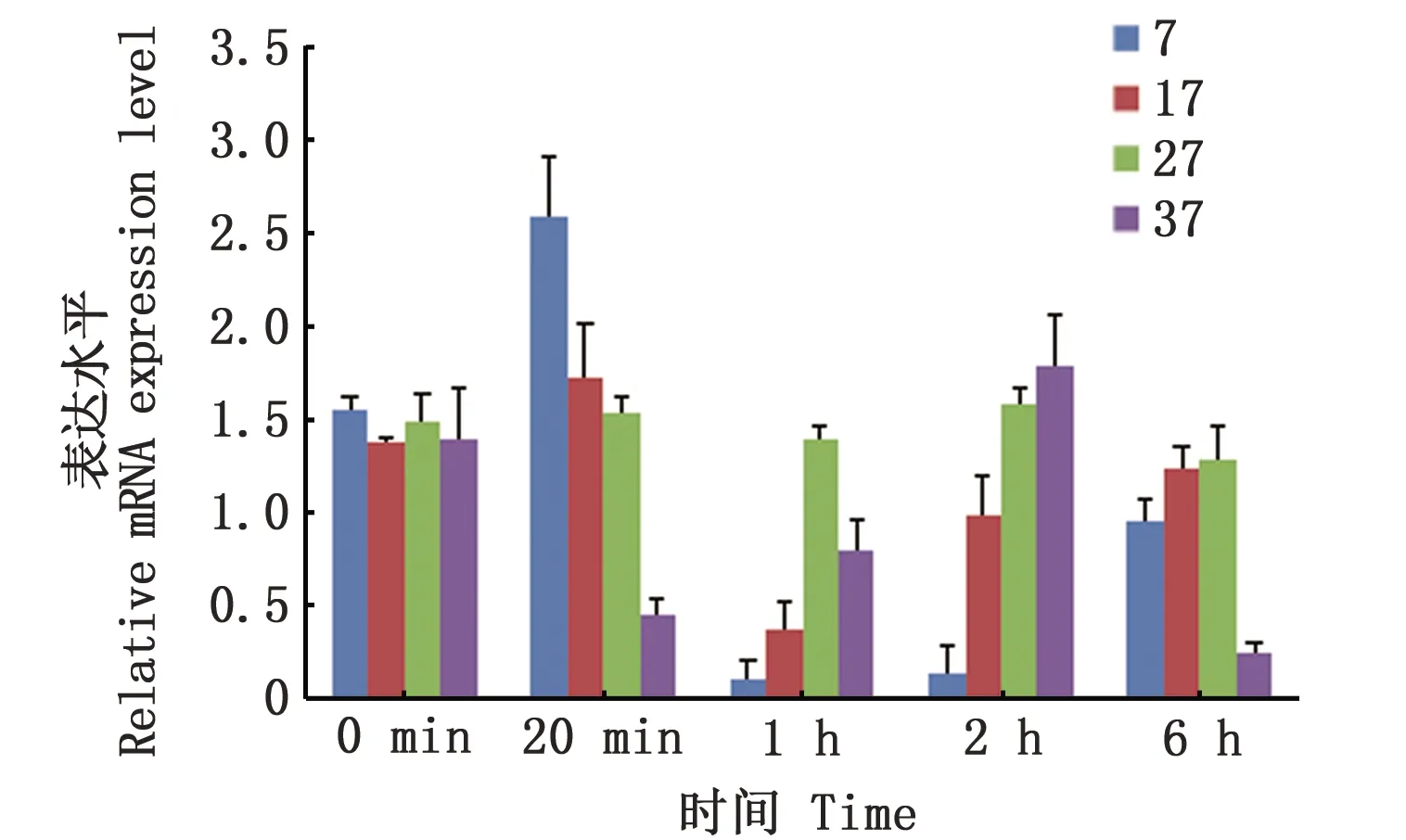
图6 盐度胁迫下Na+/H+-exchanger基因在拟穴青蟹大眼幼体期的表达情况Fig 6 Expression of Na+/H+-exchanger gene in the megalopae of Scylla paramamosainunder salinity stress
2.5 Na+/H+-exchanger蛋白结构域的比较分析
通过SWISS-MODEL在线程序对拟穴青蟹Na+/H+-exchanger蛋白结构域(82~484/961)进行三级结构的预测分析,以智人Na+/H+antiporter的三级结构模型(PDB: 4cza.1.A)为模板预测拟穴青蟹Na+/H+-exchanger蛋白结构域的三级结构,并与三疣梭子蟹(A0A286QYA6) Na+/H+-exchanger蛋白结构域(84~486/986)、斑马鱼(F1R610)Na+/H+-exchanger蛋白结构域(100~505/970)和黑腹果蝇(Q8MQP4) Na+/H+-exchanger蛋白结构域(199~603/999) 的三级结构进行比较分析(图7)。
结果表明,拟穴青蟹、三疣梭子蟹、斑马鱼Na+/H+-exchanger蛋白结构域与黑腹果蝇Na+/H+-exchanger蛋白结构域相比较更为保守,说明Na+/H+-exchanger蛋白结构域的保守性基本符合生物进化规律,但也受物种生物特性及生活环境等影响,总体而言,Na+/H+-exchanger蛋白结构域在这些海洋动物间是高度保守的。

图7 拟穴青蟹Na+/H+-exchanger domain (A)、三疣梭子蟹Na+/H+-exchanger domain (B)、斑马鱼Na+/H+-exchanger domain (C)和黑腹果蝇Na+/H+-exchanger domain (D)蛋白三维结构模型Fig 7 Three-dimensional structure models of Na+/H+-exchanger domain from Scylla paramamosain (A), Portunus trituberculatus (B),Danio rerio (C) and Drosophila melanogaster (D)
3 讨论
本研究首次克隆得到拟穴青蟹Na+/H+-exchanger基因cDNA全长,可编码961个氨基酸,其氨基酸序列与三疣梭子蟹一致性最高,达到84.9%,这一结论印证了拟穴青蟹与三疣梭子蟹同属梭子蟹科且具有相似生存环境和生活习性的特点,与PÉQUEUX[2]提出的地域分布对生物渗透调节能力具有重要影响的结论一致。SMART在线结构域预测分析发现该基因具有典型的Na+/H+-exchanger蛋白结构域(82~484),充分证明该基因为拟穴青蟹Na+/H+-exchanger基因。通过SWISS-MODEL在线程序对拟穴青蟹等物种的Na+/H+-exchanger蛋白结构域进行三级结构的预测分析,结果也表明Na+/H+-exchanger蛋白结构域在这些海洋动物间是高度保守的。TMHMM和ProtScale在线工具预测表明0~487氨基酸区段具有12个跨膜α螺旋,与拟穴青蟹Na+/H+-exchanger蛋白结构域重叠,且该区段内亲疏水性氨基酸均有分布,推测该区段可能与离子的跨膜转运功能相关[21]。此外,系统进化分析表明,拟穴青蟹Na+/H+-exchanger符合遗传进化规律。
利用RT-qPCR分析表明拟穴青蟹Na+/H+-exchanger基因的表达具有组织特异性,除性腺组织外与三疣梭子蟹和普通滨蟹Na+/H+-exchanger基因的研究结果基本一致[5,19],但三疣梭子蟹与普通滨蟹目前没有Na+/H+-exchanger基因在性腺组织表达的分析研究报道。本文首次在梭子蟹科探究该基因在性腺组织的表达,并发现该基因在拟穴青蟹性腺相关组织(卵巢、精巢和纳精囊)表达量较高,其中卵巢和精巢表达显著高于鳃。盐度的变化会直接影响拟穴青蟹变态和蜕壳周期与成功率,水环境的稳态是拟穴青蟹产卵和发育的必要条件,这个过程中Na+/H+-exchanger基因可能扮演着重要的作用。
盐度7试验组拟穴青蟹大眼幼体在20 min表达量显著高于盐度17和对照组(盐度27),表明Na+/H+-exchanger基因在较低盐度环境下的作用更加明显。盐度7和17试验组在1 h表达水平显著低于对照组(盐度27),明显受到抑制,推测是前期Na+/H+-exchanger基因的高表达已满足渗透调节作用,此时已有足够的蛋白产物。低盐组(盐度7和17)在2~6 h,Na+/H+-exchanger基因表达水平基本恢复正常,体现了拟穴青蟹大眼幼体有一定低盐环境适应能力。结果表明,拟穴青蟹Na+/H+-exchanger基因在盐度适应过程中主要在低盐胁迫下起作用,且其在低盐环境下是一个快速应答的基因,在高盐环境中作用不明显。高盐组(盐度37)该基因的表达在20 min内表达量低于对照组,推测是因为高盐度对基因表达产生抑制。
本研究探讨了盐度胁迫作用下拟穴青蟹Na+/H+-exchanger基因的表达情况,通过对拟穴青蟹Na+/H+-exchanger基因的克隆、鉴定和表达分析,初步论述了该基因的序列特征、Na+/H+-exchanger蛋白结构域及其在拟穴青蟹大眼幼体盐度适应过程中的生理作用,为水生甲壳动物渗透压调控分子机制提供了理论支撑。
猜你喜欢
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
水产养殖(2020年9期)2020-09-24
故事大王(2020年8期)2020-08-04
海峡姐妹(2018年11期)2018-12-19
北京广播电视报(2018年39期)2018-11-23
河北渔业(2018年4期)2018-05-08
现代营销·经营版(2017年3期)2017-05-17
学苑创造·A版(2016年7期)2016-07-06
作文周刊·小学五年级版(2016年1期)2016-06-29
河北渔业(2015年1期)2015-01-19
