
火炬树幼苗对镉胁迫的生理响应及积累特性
2020-06-04 02:27杨塍希国伟强和文懿王明雪曲同宝
福建农林大学学报(自然科学版) 2020年3期
杨塍希, 国伟强, 和文懿, 王明雪, 曲同宝
(吉林农业大学园艺学院,吉林 长春 130118)
随着经济的发展和城市化进程的不断加快,土壤重金属污染越来越严重.在诸多导致土壤环境污染的重金属元素中,镉污染最为广泛,我国农地的镉点位超标率高达7%[1].土壤镉超标破坏了植物细胞膜透性、生理代谢、光合代谢等系统的平衡,导致植物生长放缓、叶片褪绿、生物量下降等,同时还能通过食物链的积累威胁人类的健康与生存[2-3].因此,治理土壤镉污染已是刻不容缓.
植物修复技术是治理土壤重金属污染的重要手段,利用木本植物进行重金属污染修复是当前研究的热点,原因在于木本植物,如构树(Broussonetiapapyrifera)[4]、竹柳(Salixsp.)[5]、盐肤木(Rhuschinensis)[6]等生物量大、根系发达,吸收积累的重金属在短期内不会重新释放到环境中,同时又产生大量的木材或生物质能源.因此在植物修复方面,木本植物比超积累型草本植物具有更好生态效应和景观效应[7-9].
火炬树(RhustyphinaL.)为漆树科(Anacardiaceae)盐肤木属(Rhus)落叶灌木或亚乔木,有一定入侵性.但其易繁殖、生物量大、生长速度快、抗旱保水、耐盐碱、适应性极强,现广泛应用于荒山造林与城市园林建设[10-11].同时研究发现,火炬树叶片可积累镉[12],其根蘖苗也可用于尾矿地水土的保持[13-14],对镉具有一定的抗性与富集能力[15-17].但目前火炬幼苗在镉胁迫环境下生长、光合生理变化尚未清楚.本试验利用盆栽法,探究火炬树播种苗在受镉胁迫时的生长生理活性、光合特性及镉的积累特性,旨在探明火炬树对镉胁迫的耐受程度及适应能力,为工矿业废弃地、垃圾填埋场等的土壤镉修复提供树种选择依据.
1 材料与方法
1.1 供试材料
火炬树种子于2018年4月采收于吉林农业大学校园内,净种后千粒重为10.46 g.供试土壤采自吉林农业大学园艺学院苗圃露地表层(0~20 cm)土壤,过20目孔径筛,风干备用.土样的基本理化性质和重金属Cd背景值如下:有机质56.39 g·kg-1,全氮4.50 g·kg-1,碱解氮151 mg·kg-1,速效磷13.78 mg·kg-1,速效钾204 mg·kg-1,pH 值6.7,土壤Cd2+背景值0.25 mg·kg-1.
1.2 试验方法
于2018年5月1日—9月10日在吉林农业大学园艺学院苗圃开展试验.参考垃圾填埋场、工业区重金属Cd2+污染程度设定本试验所用的重金属浓度[18-19].以CdCl2(分析纯试剂)为供源,Cd2+浓度设定为0(以去离子水为对照)、25、50、75、100 mg·kg-1,共5个处理.采用盆栽控制试验法,基质为园土∶河沙∶草炭=2∶2∶1.称取配好的供试基质土样2 kg于花盆(20 cm×18 cm)中,盆下垫塑料托盘以防止重金属添加物渗漏,将上述设定重金属处理以水溶液形式添加至盆栽培养基质,翻拌均匀.重金属土壤钝化平衡一个月后,放入已催芽浸种的火炬树种子,每盆10粒,上覆1 mm沙土.幼苗出土10 d后定苗,每盆保留3株火炬树幼苗.自放入种子起,采用称重法每隔2 d根据每盆土壤水分状况及时补充土壤水分,土壤持水量保持在70%.苗圃内白天气温25~35 ℃,夜间气温15~20 ℃,相对湿度50%~60%,自然光照射.每个处理3个重复,共15个盆栽.于9月10日前后测定火炬树的光合特性和生长生理指标.
1.3 测定方法
1.3.1 生长指标测定 株高:用直尺垂直测量地上部分至长出完整叶片的最高点.主根长:用直尺垂直测量地下部分至主根根尖.根冠比:收获时将火炬树幼苗分为地上部(茎、叶)与地下部(根)2个部分取样,植物样品用离子水洗净,经105 ℃杀青20 min,80 ℃下烘干至恒重后,统计生物量.每个处理重复3次,取平均值.
1.3.2 叶绿素、丙二醛含量测定 采取乙醇—丙酮(1∶1)混合液浸泡法测定叶绿体色素,采用硫代巴比妥酸法测定丙二醛(MDA)含量,采用考马斯亮蓝G-250染色法测可溶性蛋白含量[20].
1.3.3 叶片气体交换参数测定 在晴朗无风的天气条件下,上午9:00—11:00,取成熟并完全展开的叶片,使用CIRAS-2便携式光合测定仪(PPSystems, America)测定叶片气体交换参数.每个处理测定3株.测定指标包括净光合速率(Pn) 、气孔导度(Gs) 、蒸腾速率(Tr)与胞间CO2浓度(Ci) ,并计算水分利用效率 (WUE=Pn/Tr) .
1.3.4 叶片叶绿素荧光参数测定 使用FMS-2脉冲式荧光仪(Hansatch, UK)测定火炬树叶绿素荧光参数,测定部位为成熟并完全展开的叶片.暗适应30 min后测定初始荧光(Fo)、最大荧光(Fm)和最大光化学效率(Fv/Fm).暗反应下荧光参数测定完后,测定光适应下的最大荧光(Fm′)、光适应下的最小荧光(Fo′)及稳态荧光(Fs)和实际光能转化率(ΦPSⅡ),通过以上数据计算出光化学淬灭系数[qP=(Fm′-Fs)/(Fm′-Fo′)],非光化学淬灭系数[qNP=(Fm-Fm′)/(Fm-Fo′)]和表观电子传递速率[ETR=ΦPSⅡ×α(0.84)×PFD,该试验矫正后PFD=1 460 μmol·m-2·s-1×0.5].
1.3.5 Cd2+含量测定 将测定完干重比的植物样品粉碎,称取0.3 g,用10 mL 65%HNO3消解,植物Cd2+含量用电感耦合等离子原子发射光谱仪ICP-OES(ISIR Intrepid Ⅱ XSO, America)测定.
转移系数(TF)=地上部分平均Cd2+含量/根部平均Cd2+含量
1.4 数据处理
用Microsoft Excel 2010进行数据统计,采用SPSS 21.0对样本数据进行单因素方差分析(one-way ANOVA),采用LSD法和Duncan检验分析样本间的差异显著性.
2 结果与分析
2.1 Cd2+胁迫对火炬树幼苗生长的影响
如表1所示,低浓度Cd2+处理下,株高、主根长与对照组相比差异不显著,随着Cd2+浓度的升高,Cd2+对火炬树幼苗的株高、主根长、地上和地下部分生物量产生显著的抑制作用(P<0.05).当Cd2+浓度为100 mg·kg-1时,抑制作用最强,株高、根长、地上和地下部分生物量分别比对照降低了46.55%、23.52%、68.67%、55.63%.根冠比在不同浓度Cd2+处理下差异不显著(P>0.05).
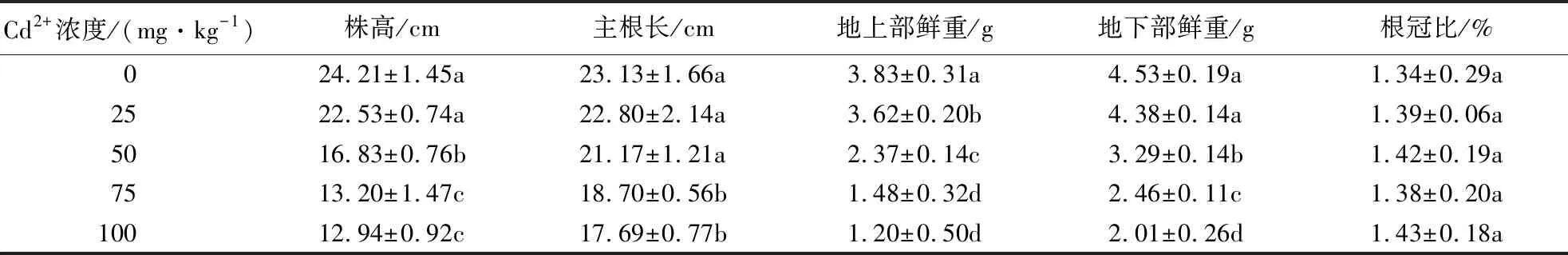
表1 Cd2+对火炬树幼苗生长的影响1)Table 1 Effects of Cd2+ on the growth and biomass of R.typhina
1)表中数据为均值±标准差,同列不同字母表示不同处理在0.05水平上的差异显著性.
2.2 Cd2+对火炬树幼苗叶片MDA含量、可溶性蛋白含量的影响

表2 Cd2+对火炬树幼苗MDA含量、可溶性蛋白含量的影响1)Table 2 Effects of Cd2+ on the MDA and soluble protein levels of R.typhina
1)表中数据为均值±标准差,同列不同字母表示不同处理在0.05水平上的差异显著性.
由表2可得,随着Cd2+浓度的升高,MDA含量逐渐上升,可溶性蛋白含量呈先升高后降低的趋势.中低Cd2+浓度(25、50 mg·kg-1)下,MDA的变化不显著;当Cd2+浓度为100 mg·kg-1时,MDA含量比对照增加了27.71%.低Cd2+浓度下,可溶性蛋白含量基本不变,随着Cd2+浓度的升高,可溶性蛋白含量显著升高(P<0.05),但当Cd2+浓度为100 mg·kg-1时,可溶性蛋白含量反而略低于对照组(P>0.05).
2.3 Cd2+对火炬树幼苗叶片叶绿素含量的影响
随着Cd2+浓度的升高,火炬树幼苗叶片叶绿素a、叶绿素b、叶绿素总含量、类胡萝卜素含量大致呈下降趋势,只有叶绿素a/叶绿素b先升高后降低.当Cd2+浓度为25 mg·kg-1时,各指标与对照相比差异不显著(P>0.05);当Cd2+浓度为100 mg·kg-1时,叶绿素a、叶绿素总含量、类胡萝卜素的含量均显著降低(P<0.05),分别比对照降低了12.08%、6.98%、10.83%、16.99%(表3).叶绿素a/叶绿素b比值的下降表明,Cd2+对叶绿素a抑制作用大于对叶绿素b的抑制作用.
2.4 Cd2+对火炬树幼苗叶片气体交换参数的影响
Cd2+胁迫对火炬树幼苗叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)的影响各不相同.当Cd2+浓度为100 mg·kg-1时,火炬树幼苗Pn、Gs、Tr、Ci含量分别比对照组降低了44.81%、47.25%、28.68%和20.85%(P<0.05).火炬树幼苗的水分利用效率(WUE)均随Cd2+浓度的升高呈下降趋势,高浓度Cd2+(100 mg·kg-1)处理下,WUE比对照组降低了26.66%(P<0.05)(表4).

表3 Cd2+对火炬树幼苗叶绿素含量的影响1)Table 3 Effects of Cd2+ on the chlorophyll contents of R.typhina
1)表中数据为均值±标准差,同列不同字母表示不同处理在0.05水平上的差异显著性.

表4 Cd2+对火炬树幼苗光合气体参数的影响1)Table 4 Effects of Cd2+ on photosynthetic parameters of R.typhina
1)表中数据为均值±标准差,同列不同字母表示不同处理在0.05水平上的差异显著性.
2.5 Cd2+对火炬树幼苗叶片叶绿素荧光参数的影响
火炬树幼苗叶片最大量子产量(Fv/Fm)、实际光能转化率(ΦPSⅡ)、光化学淬灭系数(qP)、非光化学淬灭系数(qNP)和表观电子传递速率(ETR)均随Cd2+浓度的升高持续下降,初始荧光(Fo)不断增大,最大荧光(Fm)呈先上升后下降趋势.当Cd2+浓度为100 mg·kg-1时,Fo比对照增大了18.32%,Fm、Fv/Fm、qP、qNP、ETR分别比对照减小了24.42%、10.71%、44.96%、34.87%、39.72%,差异显著 (P<0.05),ΦPSⅡ差异不显著(P>0.05)(表5).

表5 Cd2+对火炬树幼苗叶绿素荧光参数的影响1)Table 5 Effects of Cd2+ on chlorophyll fluorescence parameters of R.typhina
1)表中数据为均值±标准差,同列不同字母表示不同处理在0.05水平上差异显著性.
2.6 火炬树幼苗对Cd2+的积累特性
由表6可得,随着Cd2+浓度的升高,叶片和茎的Cd2+积累量呈先增加后降低的趋势,根系的Cd2+积累量则不断上升(P<0.05).当Cd2+浓度为100 mg·kg-1时,根系的Cd2+积累量高达27.16 mg·kg-1,显著高于对照组和其他试验组.当Cd2+浓度为25 mg·kg-1时,火炬树幼苗转移系数为0.29,略高于其他处理组,但显著低于对照组.

表6 火炬树幼苗Cd2+积累特性1)Table 6 Accumulation characteristics of Cd2+ in R.typhina
1)表中数据为均值±标准差,同列不同字母表示不同处理在0.05水平上的差异显著性.
3 讨论与结论
植物生长与生物量的积累是植物修复技术的重要指标,能直接反映植物修复的效率[21].本试验中,低浓度Cd2+胁迫下,火炬树幼苗株高、主根长和生物量与对照相比均无显著差异,随着浓度的升高,此3项指标均呈显著下降趋势,且Cd2+对地上部分的抑制作用大于根部,这与燕江伟等[22]对互叶醉鱼草(Buddlejaalternifolia)的研究结果相似.这可能是因为生长在污染区的木本植物,根系通常采取避逆性机制,向污染较少的区域生长来降低重金属胁迫对根系的损伤[23],因此地下部分受重金属影响的程度低于地上部分.本试验中处理组的火炬树幼苗根系多局限在容器1/3处,主根盘曲,大量须根横向分布于距表层3~5 cm的土壤中,与对照中火炬树幼苗根系分布于整个盆栽土壤存在明显区别,这与施翔等[6]对盐肤木的研究结果相似.这表明火炬树幼苗通过减少根系与重金属的接触面积,来降低重金属对根系的伤害,表现出良好的耐性.
Cd2+对植物生理的影响多体现在MDA与可溶性蛋白的变化量上[24-25].MDA是植物在逆境条件下发生膜脂过氧化作用的产物,其含量的多少体现了植物对重金属环境适应能力的强弱[26].本试验中,随着Cd2+浓度的升高,火炬树幼苗叶片中MDA含量呈逐渐上升趋势,这与红瑞木(Swidaalba)[8]在Cd2+胁迫下的反应一致.可溶性蛋白含量的变化是衡量植物对重金属抗性的一个关键生理参数[27],火炬树幼苗叶片的可溶性蛋白含量随Cd2+浓度的升高呈先升高后降低的趋势,这与苦楝(Meliaazedarach)受Cd2+胁迫的结果相似[24].在低浓度Cd2+胁迫下,Cd2+可诱导植物合成重金属螯合肽(Cd-PC)以减少Cd2+对细胞的毒害,这时表现为可溶性蛋白含量增加;但当重金属胁迫进一步增强时,Cd-PC表达受到抑制,无法结合更多的Cd2+,此时可溶性蛋白含量降低[28].
重金属对植物生长和代谢的影响是多方面的,其中对光合作用的抑制尤为显著,叶绿素含量的高低直接影响植物光合作用水平[29].本试验中,火炬树幼苗叶片的光合色素含量均随着Cd2+浓度的升高而下降,与生物量变化规律相同,且叶绿素a比叶绿素b对Cd2+更加敏感,这与紫花苜蓿(Medicagosativa)受Cd2+胁迫时光合特性的变化规律一致[30].这是因为Cd2+会使类胡萝卜素等捕光色素复合体受损,破坏叶绿素酶系统,阻碍叶绿素a的合成,导致光合系统Ⅱ比光合系统Ⅰ更敏感,进而表现出叶绿素a比叶绿素b减少得更为显著[31].
光合作用气体交换参数是植物在环境变化中最为敏感的适应性特征之一[32].本试验发现,火炬树幼苗叶片Pn、Gs、Tr和Ci、WUE均受到Cd2+的抑制.但当Cd2+浓度为25 mg·kg-1时,Pn的下降幅度较小,说明在低浓度Cd2+胁迫下火炬树幼苗具有较强的光合适应性,可进行正常的光合生长,与张嘉桐等[33]对桑树(Morusalba)的研究结果一致.本试验中Pn下降的同时Ci值也明显下降,说明气孔限制是导致火炬树幼苗在Cd2+胁迫下Pn下降的主要原因.而气孔受限制主要是由于Cd2+在植物体内的积累使得气孔细胞的细胞壁弹性下降,并扰乱气孔保卫细胞中脱落酸等植物激素的合成,最终导致气孔长期处于关闭状态[34].
相比于“表观性”的气体交换参数,叶绿素荧光动力学参数更能反映植物“内在性”的特点.Fv/Fm不仅是光抑制和PSⅡ复合体受损的指标,也反映了植物潜在的最大光合能力.本试验中,低浓度处理下火炬树幼苗叶片的Fo、Fm较对照组均有不同程度的升高(P<0.05),但这程度的上升未对PSⅡ反应中心造成破坏;随着Cd2+浓度的升高,Fo持续上升,Fm却出现下降趋势,且Fv/Fm、ΦPSⅡ显著降低.此时Fo值的持续上升说明PSⅡ反应中心受损或不可逆失活,这与简敏菲等[35]对丁香蓼(Ludwigiaprostrata)的研究结果一致.同时在本试验中,火炬树幼苗叶片的qP、qNP和ETR也随Cd2+浓度的升高而显著降低,这与Cd2+胁迫下杨树(Populusxeuramericana)的叶绿素荧光特性变化一致[38].这些指标的变化均说明火炬树幼苗叶片中放氧复合体或捕光色素复合体受到了一定程度损伤.研究表明,高浓度Cd2+离子会干扰光合电子传递过程,导致PSⅡ反应中心部分失活的同时也抑制放氧复合体的电子传递,使得PSⅡ原初电子受体QA接受电子能力减弱,光能转化效率及捕获激发能的效率降低[36-37].
重金属积累能力与转移能力也是评价植物能否作为重金属污染土壤修复树种的重要指标.本试验中,火炬树幼苗单株Cd2+积累量最高为32.76 mg·kg-1,植株不同部位Cd2+积累能力表现为根>茎>叶.这是因为与根茎相比,火炬树幼苗叶片生理功能较弱,通过利用根部区域化固定、金属硫蛋白合成、Cd-PC形成等方式,将更多Cd2+贮蓄在根茎中,减少Cd2+向地上部分转移,这一分配策略有利于降低Cd2+对地上部分的生物毒性,体现了植物对Cd2+的耐性[38].
综上所述,Cd2+胁迫下,火炬树幼苗根系不可避免地吸收土壤中的Cd2+,导致根系生长受到抑制,植物营养元素和水吸收平衡变得紊乱,火炬树的生物量积累受到影响;根部吸收积累的Cd2+转运迁移至火炬树地上部分,使得植物叶片细胞发生膜脂过氧化,叶绿体结构被破坏,叶绿素合成受抑制,PSⅡ复合体受损害,表现为光合色素含量降低,光合速率与光合效率下降,影响光合产物的积累,最终表现为植物生长受到限制,总生物量降低.火炬树幼苗在中轻度Cd2+污染(25~50 mg·kg-1)的土壤中,生长指标、生化生理特性、光合作用受到的影响较小,具有较强的抗性与重金属积累能力,说明火炬树在轻度Cd2+污染的土壤环境中能较好地存活和生长,在植物修复领域,特别是受镉轻度污染的场地,具有广泛的应用前景.
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
音乐天地(音乐创作版)(2021年7期)2021-10-13
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
阅读(科学探秘)(2020年8期)2020-11-06
安徽农学通报(2020年7期)2020-05-26
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
中国火炬(2015年3期)2015-07-31
中国火炬(2015年2期)2015-07-25
