
外源茉莉酸甲酯对干旱胁迫下狭叶黄芩光合和生理特性的影响
2020-06-03 05:18吴建慧范卫芳高鹏飞
植物研究 2020年3期
吴建慧 范卫芳 牛 喆 张 静 高鹏飞 李 文
(东北林业大学园林学院,哈尔滨 150040)
茉莉酸甲酯(MeJA)是一类在植物体内合成的脂类衍生化合物,在植物正常的生长发育过程以及受到外界胁迫条件时,其可以同时作为信号分子和植物逆境相关的生长调节物质在此过程中发挥巨大的作用[1]。外源喷施时,MeJA能够通过气孔进入植物体内,进行长距离的信号转导以及植物之间的信息交流,从而诱导植物产生防御响应[2~3]。茉莉酸类化合物对植物耐旱性的影响在小麦[4]、大豆[5]等植物中已有报道。其主要通过影响植物的光合作用、渗透调节物质以及抗氧化酶系统等发挥作用。韩瑞宏等[6]对紫花苜蓿研究表明,干旱胁迫下MeJA在调节气孔运动、维持叶绿素含量方面起到了关键作用。外源MeJA减轻了干旱胁迫下烤烟叶片叶绿素的分解[7]。MeJA有效促进干旱胁迫下小麦叶片中β-胡萝卜素的积累,保护其他质体色素免受氧化降解,并通过调节气孔导度以避免植株水分的散失,从而缓解了干旱胁迫对光合同化能力的不良影响,增强小麦的抗旱性[8]。董桃杏等[2,9]的研究表明外源MeJA可以显著改善水分亏缺所导致的叶绿素降解、蒸腾速率降低以及光合作用的减弱。脯氨酸和可溶性糖作为重要的渗透调节物质,在逆境条件下可以通过降低细胞渗透势的方法来达到提高细胞吸水性的目的,使植物减轻不良环境带来的影响。研究发现[10],低浓度(0.25 μmoL·L-1)外源MeJA浸种能够促进干旱胁迫下玉米种子的萌发以及幼苗正常生长,促进植物幼苗的脯氨酸含量升高,高浓度(≥25 μmoL·L-1)MeJA浸种则对种子萌发和幼苗生长产生明显的不良影响。可溶性蛋白的积累也可以帮助减缓干旱胁迫对植物的伤害[11]。而且,在缺水状况下植物会产生大量活性氧,细胞发生膜脂过氧化,膜系统受到氧化伤害,导致植物组织损伤[12~13]。SOD、POD和CAT是与植物抗旱性密切相关的抗氧化保护酶[14]。邹清成等[15]对蝴蝶兰的研究发现施加MeJA使超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性升高,丙二醛(MDA)含量降低,表明一定浓度的MeJA处理可通过上调蝴蝶兰幼苗的抗氧化酶活性以增强抗逆性。
自然分布在东北地区的狭叶黄芩(ScutellariaregelianaNakai)是一种优良的野生植物,具有很高的观赏价值,可作为地被植物在园林中应用。目前JA类的作用已经在部分观赏植物以及农作物上得到很好的验证,但尚未对狭叶黄芩进行研究,因此本试验以狭叶黄芩为研究对象,探讨外援施加MeJA对狭叶黄芩的影响,以期通过本试验能够为狭叶黄芩在干旱地区的园林应用提供一定的理论参考。
1 材料与方法
1.1 材料
试验材料取自黑龙江省森林植物园的野生狭叶黄芩,移栽到花盆经过一段时间的缓苗后,于2018年7月选取生长一致、发育正常的狭叶黄芩进行试验。
1.2 试验方法
1.2.1 试验材料处理
将狭叶黄芩用15%的PEG-6000溶液进行干旱胁迫1周,用无水乙醇和蒸馏水将MeJA配制成浓度梯度为0.1、0.5、1.0、2.0 mmol·L-1的溶液(分别记为MJ1、MJ2、MJ3、MJ4),以无水乙醇加蒸馏水为对照(记为CK),采用喷雾法对狭叶黄芩地上部分叶片进行正反面均匀喷施。直至叶片上溶液欲滴为止,待叶片表面溶液自然蒸干后,将不同浓度处理的植株分开放置,在喷施外源物质后的第1、3、6、9和12 d选择健康的叶片进行取材用于生理指标的测定,每次取样重复3次。
1.2.2 表型特征观察
在喷施外源物质后的第1、3、6、9和12 d定时观察干旱胁迫下狭叶黄芩受害后的症状特征,包括老叶、新叶、嫩茎及茎干的颜色和形态。根据受害程度,划分狭叶黄芩受害的等级。0级:植株正常。Ⅰ级:顶芽萎蔫。Ⅱ级:单株小叶黄叶占1/3。Ⅲ级:单株小叶黄叶占1/2。Ⅳ级:单株小叶黄叶占2/3。Ⅴ级:整株小叶变褐[16]。
1.2.3 光合作用参数测定
采用美国Li-Cor公司的LI-6400便携式光合仪来测定光合作用参数。测量均在上午9:00~11:00进行,测量时选取生长良好、大小一致的叶片,每个处理测量3个叶片,每个叶片重复记录3次数据。测定指标包括叶片净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1)。
1.2.4 生理生化指标测定
脯氨酸含量的测定:采用李合生生理试验方法[17];蛋白质含量的测定:采用樊金娟的考马斯亮蓝G-250染色法测定蛋白质含量[18];超氧物歧化酶(SOD)含量的测定:采用氮蓝四唑(NBT)光还原法测定SOD活性[17];过氧化物酶(POD)活性的测定:采用刘萍的愈创木酚法测定POD活性[19];过氧化氢酶(CAT)活性的测定:采用紫外吸收法测定CAT活性[19]。
1.3 数据统计与分析方法
采用Excel 2007对测定的数据进行计算,采用SPSS22.0统计软件进行方差分析和显著性分析,采用Origin8.1进行绘图和制表,数据均为3次重复试验的平均值。
2 结果与分析
2.1 茉莉酸甲酯对干旱胁迫下狭叶黄芩表型变化的影响
由表1可知,随着干旱胁迫时间的增加,未处理组的狭叶黄芩受害程度由0级增加到Ⅳ级,而在不同浓度MeJA处理下,除了MJ1处理组,其他处理组在干旱胁迫下的受伤程度明显降低。由此可见,外源施加的MeJA处理能够有效减缓干旱胁迫对狭叶黄芩造成的影响。
表1 狭叶黄芩不同处理组的受害等级
Table 1 The damage level of different treatment groups ofS.regeliana
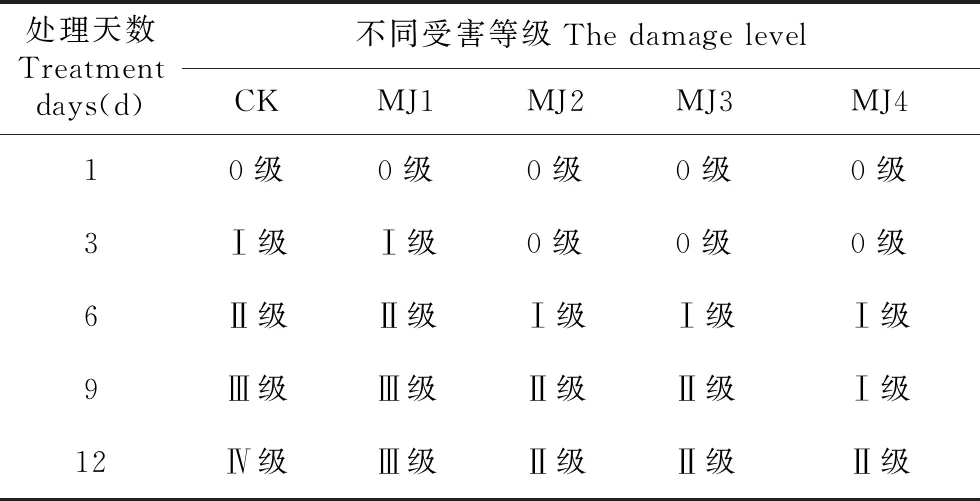
处理天数Treatment days(d)不同受害等级The damage levelCKMJ1MJ2MJ3MJ410级0级0级0级0级3Ⅰ级Ⅰ级0级0级0级6Ⅱ级Ⅱ级Ⅰ级Ⅰ级Ⅰ级9Ⅲ级Ⅲ级Ⅱ级Ⅱ级Ⅰ级12Ⅳ级Ⅲ级Ⅱ级Ⅱ级Ⅱ级
2.2 茉莉酸甲酯对干旱胁迫下狭叶黄芩光合特性的影响
由图1可知,干旱胁迫下,未处理的狭叶黄芩的净光合速率(Pn)呈下降趋势。各处理组的净光合速率随着时间的延长先上升后下降。特别是经MeJA处理后6和9 d时,所有处理组均显著高于CK,并且在处理9 d的MJ4处理组净光合速率达到最高,为20.59 μmol·m-2·s-1,相比CK增加了552.66%。12 d时所有处理组的净光合速率均下降。
不同浓度MeJA处理下,狭叶黄芩的胞间二氧化碳的变化如图2所示,在干旱胁迫下,用MeJA处理后1 d时,MJ1显著高于CK,其他处理组则显著低于CK。处理后3 d时,所有处理组则分别显著高于CK。处理后6、9 d时,所有处理组均低于CK。处理12 d时,MJ1相比CK有所上升,MJ2、MJ3、MJ4处理组均下降。
由图3可以看出,随着时间的延长,干旱胁迫下未处理组的狭叶黄芩的气孔导度均下降。MeJA处理后3、6、9 d时,所有处理组的气孔导度均显著高于CK,其中在用2 mmol·L-1MeJA处理9 d后,MJ4处理组气孔导度达到最高,为0.35 mol·m-2·s-1,相比CK增加了491.71%。处理后12 d时MJ2相对于其他处理组显著高于CK,比CK增加了163%。
由图4可知,干旱胁迫下未处理组的狭叶黄芩的蒸腾速率下降,处理组的蒸腾速率则先上升后下降。在MeJA处理后3、6、9 d时,所有处理组的蒸腾速率都高于CK,其中用2 mmol·L-1MeJA处理9 d时狭叶黄芩的蒸腾速率达到最高,为9.25 mmol·m-2·s-1,相比CK增加了574.25%。而处理12 d后除MJ4蒸腾速率的降低外,其他处理组均高于CK。
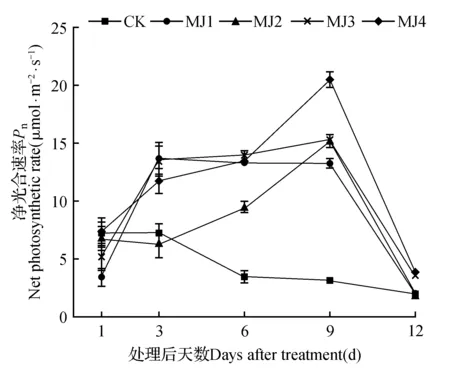
图1 干旱胁迫下茉莉酸甲酯对狭叶黄芩净光合速率的影响Fig.1 Effects of MeJA on net photosynthetic rate of S.regeliana Nakai under drought stress
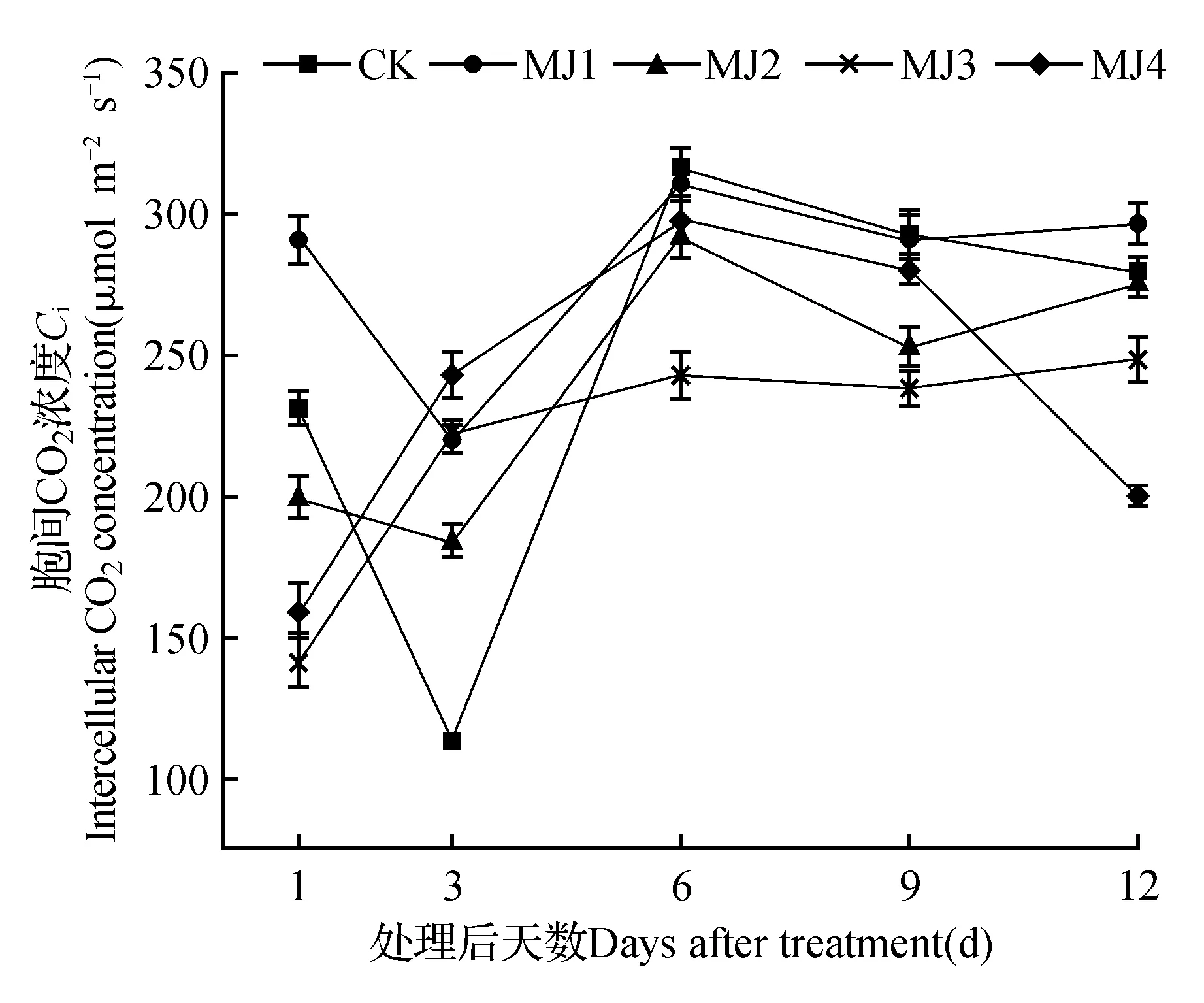
图2 干旱胁迫下茉莉酸甲酯对狭叶黄芩胞间CO2浓度的影响Fig.2 Effects of MeJA on intercellular CO2 concentration of S.regeliana Nakai under drought stress

图3 干旱胁迫下茉莉酸甲酯对狭叶黄芩气孔导度的影响Fig.3 Effects of MeJA on stomatal conductivity of S.regeliana Nakai under drought stress
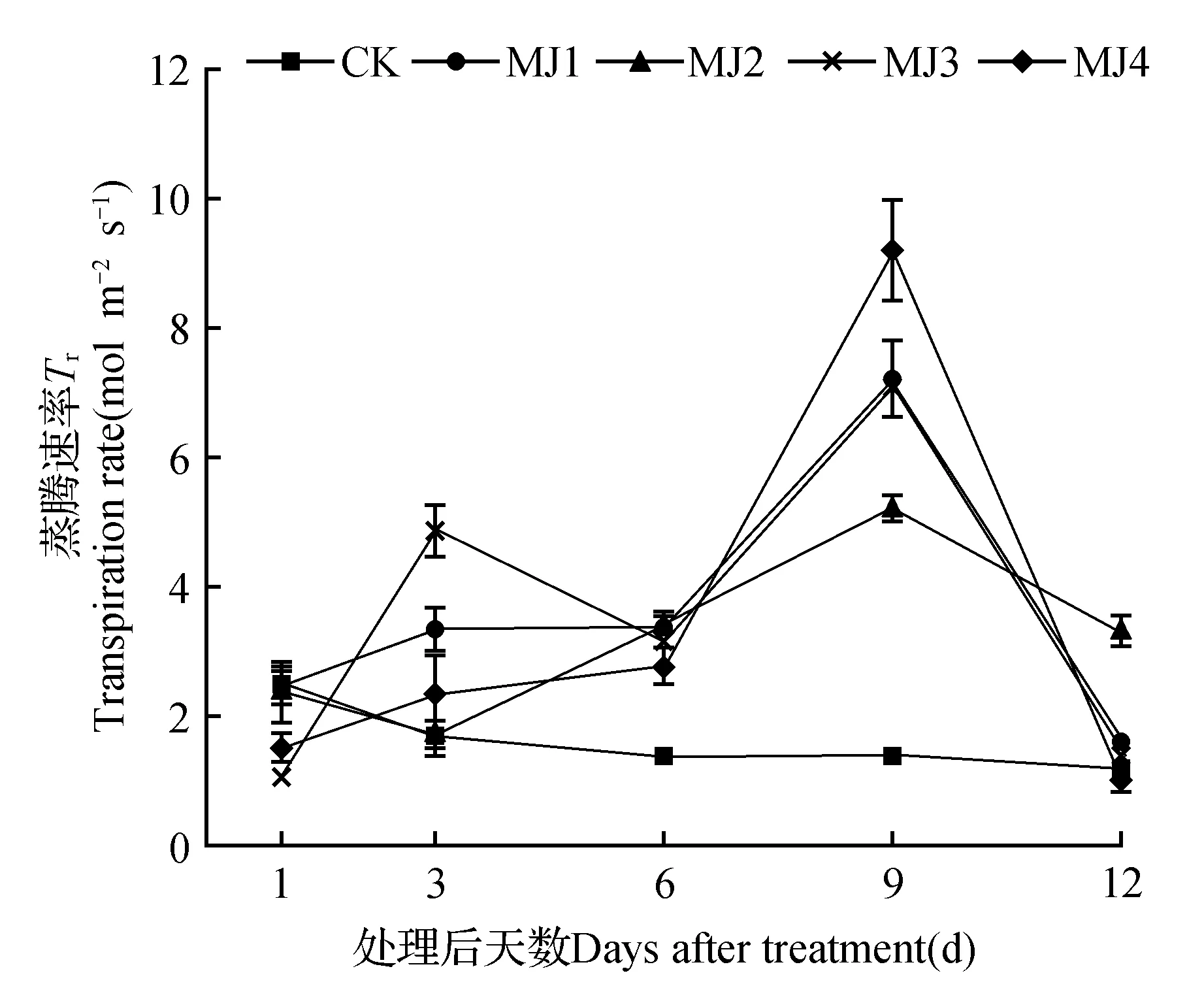
图4 干旱胁迫下茉莉酸甲酯对狭叶黄芩蒸腾速率的影响Fig.4 Effects of MeJA on transpiration rate of S.regeliana Nakai under drought stress

图5 干旱胁迫下茉莉酸甲酯对狭叶黄芩游离脯氨酸含量的影响Fig.5 Effects of MeJA on the free proline content of S.regeliana Nakai under drought stress
2.3 茉莉酸甲酯对干旱胁迫下脯氨酸和可溶性蛋白含量的影响
由图5可知,在干旱胁迫下,未处理组的狭叶黄芩的游离脯氨酸含量随着胁迫时间的延长其含量不断增加。用MeJA处理后1 d时,所有处理组的游离脯氨酸含量都高于CK,但都与CK无显著差异。处理后3 d时,除了MJ1相比CK增加了1.32%,其他处理组与CK相比均下降了。在MeJA处理6、9、12 d时,所有处理组的游离脯氨酸含量均显著低于CK。
由图6可知,在干旱胁迫下,未处理组的狭叶黄芩的可溶性蛋白随着时间的增加呈现先缓慢上升后下降的趋势,在MeJA处理后1和3 d时,所有处理组的可溶性蛋白均低于CK,6 d时MJ4显著高于CK3.54%,MJ1、MJ2、MJ3处理组则显著低于CK,处理9和12 d后,所有处理组均显著高于CK。MJ3在处理12 d后可溶性蛋白含量达到最大值,相比CK增加了88.54%。
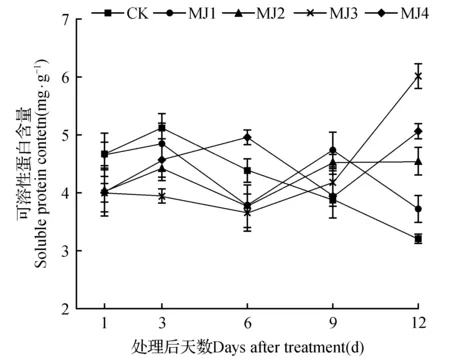
图6 干旱胁迫下茉莉酸甲酯对狭叶黄芩可溶性蛋白含量的影响Fig.6 Effects of MeJA on soluble protein content of S.regeliana Nakai under drought stress

图7 干旱胁迫下茉莉酸甲酯对狭叶黄芩SOD活性的影响Fig.7 Effects of MeJA on SOD activity of S.regeliana Nakai under drought stress
2.4 茉莉酸甲酯对干旱胁迫下过氧化酶活性的影响
由图7可知,在MeJA处理1、3、12 d后,所有处理组的SOD活性均高于CK,其中处理3 d的MJ3处理组的SOD活性为所有处理组中最高为242.98 U·g-1,相比CK增加了347.55%。处理后6和9 d时,MJ1、MJ3显著高于CK,MJ2、MJ4显著低于CK,并且两组的变化趋势相同。
由图8可以看出,干旱胁迫下所有处理组的POD活性均高于CK,并且与CK有显著性差异,在MeJA处理6 d时,MJ3处理组的POD活性为所有处理组中最高,为37.69 μg·min-1,比CK显著增加了147%。
由图9可知,在MeJA处理3、6、12 d时,所有处理组的CAT活性均高于CK。其中MJ2处理组在6 d时的CAT活性相比CK增加了1 255.76%,在12 d时的CAT活性最高,相比CK增加了797.1%。在9 d时,MJ1、MJ2、MJ4显著高于CK,MJ3则显著低于CK。

图8 干旱胁迫下茉莉酸甲酯对狭叶黄芩POD活性的影响Fig.8 Effects of MeJA on POD activity of S.regeliana Nakai under drought stress
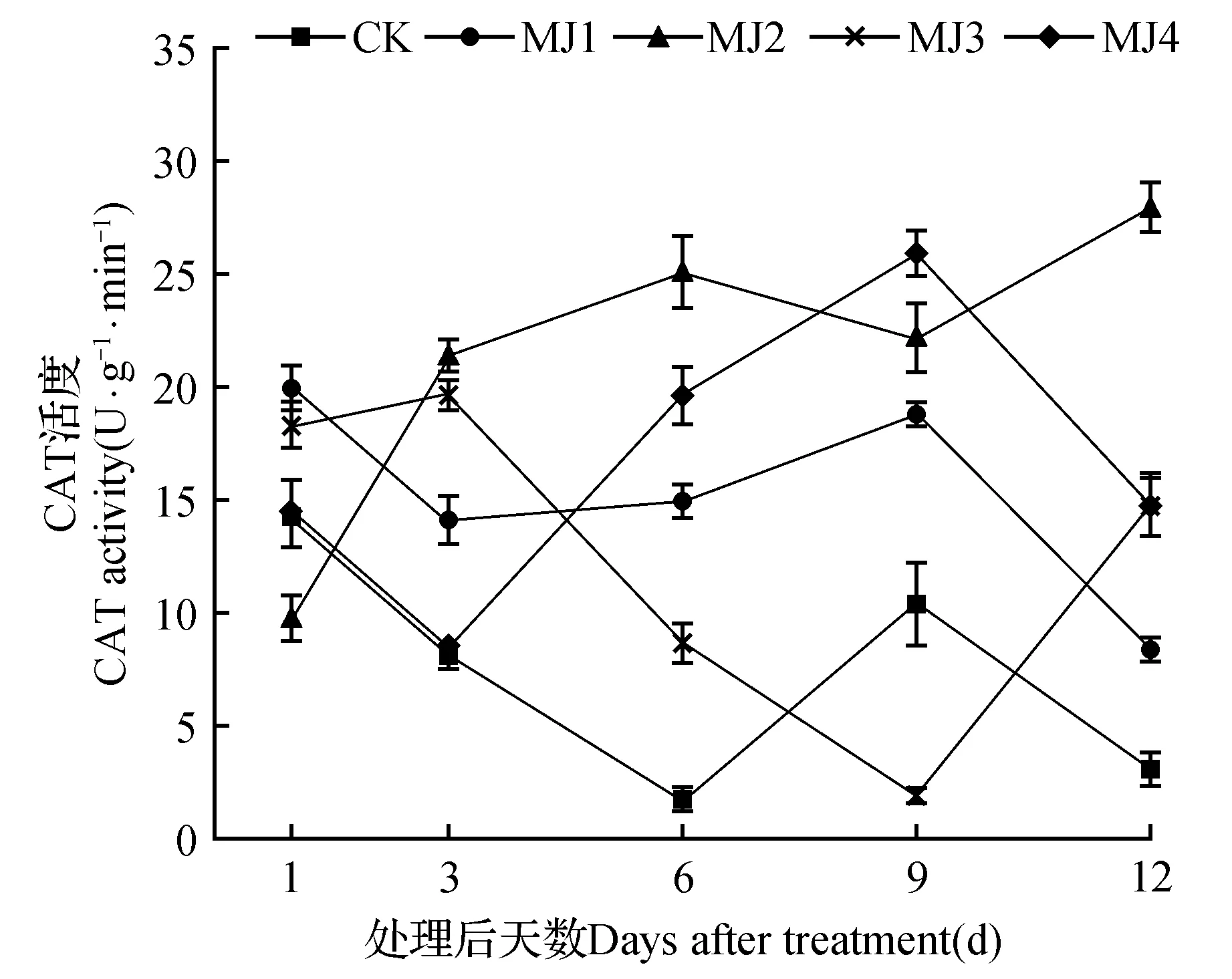
图9 干旱胁迫下茉莉酸甲酯对狭叶黄芩CAT活性的影响Fig.9 Effects of MeJA on CAT activity of S.regeliana Nakai under drought stress
3 讨论
随着干旱胁迫时间的增加,土壤含水率减少,狭叶黄芩自身的新陈代谢发生了变化,进而在形态上也表现出来,外部表现特征常常作为最直观的抗旱指标。如戴建良、王芳[20]通过对侧柏、掌叶木幼苗在干旱胁迫下植株表现的观察来研究植物的抗旱性。在本研究中,外源施加MeJA显著降低了狭叶黄芩受伤害等级。这一研究结果表明,MeJA能够在一定程度上缓解干旱胁迫对狭叶黄芩的伤害。
绿色植物光合作用强弱对植物生长和抗逆性大小都具有十分重大的影响,可用来作为判断植物生长和抗逆性强弱的指标。干旱胁迫下外源MeJA处理可显著提高水稻幼苗的叶绿素含量及光合作用[9]。在本研究中,干旱胁迫导致狭叶黄芩的净光合速率、气孔导度和蒸腾速率呈下降趋势。外源施加MeJA虽然降低了狭叶黄芩胞间CO2浓度。但下降较为缓慢。植物在干旱胁迫下,引起植物光合效率降低因素主要有两类,一是的植物自身气孔的部分关闭导致的气孔限制,二是叶肉细胞光合活性下降导致的非气孔限制[19]。许大全[21]认为,Ci和气孔导度同时降低才表明光合速率降低是以气孔限制为主。实验结果表明,干旱胁迫3 d时,未处理组Ci值和气孔导度都降低,因此在干旱胁迫3 d前狭叶黄芩光合效率降低原因是气孔导度下降导致的气孔限制。处理3 d后,Ci值和气孔导度增高,表明此时引起光合速率下降的主要原因则是以非气孔限制为主。在施加MeJA后,狭叶黄芩的净光合速率、气孔导度和蒸腾速率相比CK增加明显,并且随着时间的延长呈现先升高后下降的趋势,其中在9 d的MJ4处理组达到最大,说明MeJA能够在一定程度上缓解干旱胁迫对狭叶黄芩的伤害,促进叶片气孔张开。究其原因是施加MeJA在干旱胁迫初期,能提高狭叶黄芩的气孔开度,从而提高光合能力。而在胁迫后期则可能通过减轻干旱胁迫下狭叶黄芩光合机构所受的伤害,抑制狭叶黄芩光合速率的下降。因此MeJA在干旱胁迫初期可以改善由气孔限制引起的光合作用下降,而在胁迫后期主要是改善由非气孔限制导致的光合作用下降。
外源引入MeJA可通过诱导植物产生一些防御化合物及抗氧化剂来提高作物的抗旱性[22]。脯氨酸可保持细胞与环境渗透平衡,防止植物水分散失,是植物重要的渗透调节物质。在干旱胁迫下,植物体内脯氨酸的含量会显著增加。而刘欢等[23]研究发现不同程度的干旱处理,脯氨酸含量会有不同程度的积累,中度干旱含量最高,轻度干旱含量最低。在本研究中,未处理组狭叶黄芩体内的脯氨酸含量逐渐升高,而施加外源MeJA后各处理组脯氨酸含量先升高后降低,且所有处理组脯氨酸含量的积累程度均低于CK。表明CK相对于处理组来说受胁迫程度更大,因此外源施加MeJA可以缓解狭叶黄芩的干旱程度。植物体内积累的可溶性蛋白质越高,细胞的渗透势就会越低,从而抵抗干旱胁迫中水分对植物生命活动的威胁[24]。在本研究中,干旱胁迫下经MeJA处理后的狭叶黄芩可溶性蛋白含量在前6 d的处理中趋于或低于CK,可能是处理时间较短,MeJA还没起到缓解作用,而在6 d以后的处理中显著高于CK,说明MeJA处理6 d后可显著缓解干旱胁迫对狭叶黄芩的不良影响,研究结果与王燕芳等[25]和李顺欣等[26]的研究结果相似。
抗氧化酶系统能清除体内活性氧,从而保护植物免受氧化损伤。茉莉酸类物质作为一种植物信号转导分子,参与植物的各种生长发育和胁迫防御过程[27]。有研究发现,MeJA的作用类似植物激素,作用效果受植物种类和浓度的影响存在差异,浓度适宜能促进植物的生长或缓解胁迫对植物造成的伤害,高浓度则可能会起抑制作用[28]。苏金为[29]在对茶苗的研究中发现,施加外源MeJA能够有效保证茶苗植株体内氧自由基以及抗氧化物的平衡,缓解植株因缺水造成的氧化损伤。在本研究中,外源施加MeJA可以有效提高狭叶黄芩SOD、POD、CAT的活性,说明MeJA能够有效减轻干旱胁迫对狭叶黄芩带来的不良影响。这与苹果[30]幼苗在干旱胁迫下,其抗氧化酶活性在施加外源MeJA后,均表现出了不同程度的上升趋势的研究结果相似。狭叶黄芩在干旱胁迫下造成了膜脂过氧化加剧,植物的抗氧化系统遭到破坏。外源施加MeJA能够增强受旱狭叶黄芩体内SOD、POD和CAT等保护酶的活性,提高抵抗氧自由基和过氧化物的能力,使氧自由基和过氧化物代谢平衡维持在较高水平,减少细胞膜脂过氧化作用,从而有效缓解狭叶黄芩所遭受的干旱胁迫。这与汤日圣等[31]在水稻上的研究结论一致。在本研究中通过对狭叶黄芩的光合参数和生理指标的研究结果表明,经外源施加的MeJA处理能够有效减缓干旱胁迫对狭叶黄芩的不良影响。
猜你喜欢
中老年保健(2022年3期)2022-11-21
农业工程学报(2022年3期)2022-04-16
中国畜禽种业(2021年9期)2021-09-22
安徽化工(2021年3期)2021-05-29
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
棉花学报(2020年3期)2020-08-08
节水灌溉(2020年7期)2020-07-15
科学与财富(2019年15期)2019-10-21
润滑与密封(2019年3期)2019-03-22
