
2016-2017年我国华北地区猪繁殖与呼吸综合征病毒类NADC30毒株分子遗传演化分析
2020-05-11 03:35石青青李留安马吉飞孙英峰
中国动物传染病学报 2020年3期
石青青,姜 轩,张 奥,李留安,马吉飞,孙英峰
(天津农学院动物科学与动物医学学院,天津300384)
猪繁殖与呼吸综合征(Porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(Porcine reproductive and respiratory syndrome virus,PRRSV)引起的一种接触性传染病,临床主要表现为怀孕母猪繁殖障碍、仔猪呼吸系统症状和高死亡率等[1]。自20世纪80年代末,该病从美国和欧洲相继暴发以来,迅速在全球蔓延。1996年在国内首次分离报道[2],2006年在我国江西地区发生了高致病性猪繁殖与呼吸综合征病毒(Highly pathogenic PRRSV,HP-PRRSV)后,该病蔓延至全国大部分省份,给猪场造了成巨大经济损失[3]。
自2014年以来,国内多个实验室相继报道了一种与美国NADC30毒株高度同源,且在Nsp2存在不连续的30个氨基酸缺失模式的毒株,该类毒株极易与国内流行的其他亚群毒株(经典PRRSV、HPPRRSV、疫苗毒株或类疫苗毒株等)发生重组,经遗传变异产生了一类新病毒,国内学者将这类病毒统称为类NADC30毒株(NADC30-like)[4-5]。类NADC30感染猪场易造成种猪群和生长猪群生产极不稳定,且现有的疫苗保护效果不佳[6-7],目前已是猪场主要流行毒[8-9]。本实验室2016-2017年从华北四省市(北京、天津、河北、山东)通过对类NADC30毒株进行了ORF5基因的序列测定,并对其中ORF5部分核苷酸缺失毒株进行全基因组序列分析,研究其与参考毒株之间的遗传演化关系,揭示我国华北地区当前流行的PRRSV类NADC30的抗原特点和变异规律。
1 材料与方法
1.1 主要实验材料 236份疑似PRRSV感染的样品为2016-2017年采集自华北四省市(北京、天津、河北、山东)部分猪场。RNA提取试剂盒、反转录试剂盒(AMV-L)、DNA胶回收试剂盒、pMD18-T载体和质粒提取试剂盒购自天根生化科技(北京)有限公司;Buffer、dNTPs、TaqDNA聚合酶等购自宝生物(大连)有限公司。
1.2 引物设计及样品RT-PCR检测 根据参考文献[6]合成ORF5基因扩增引物,由生物生物工程(上海)有限公司合成。利用RNA提取试剂盒提取样品总RNA,经反转录成cDNA,放-20℃冰箱备用。PCR反应程序为:94℃预变性5 min;94℃变性30 s、56℃退火30 s、72℃延伸30 s,共35个循环;72℃再延伸5 min。PCR产物经1%琼脂糖凝胶电泳检测后,采用DNA胶回收试剂盒回收,克隆至pMD18-T载体,将阳性质粒送测序公司测序。
1.3 PRRSV的分离鉴定 取阳性样品血清或组织研磨病料用0.22 μm的滤器过滤后,分别感染PAMs细胞和Marc-145细胞,孵育2 h后PBS清洗,加入2 mL DMEM完全培养基,37℃、5%CO2连续培养72 h后,每天观察CPE并收取上清液,盲传3代后,然后对上清液进行RT-PCR检测。
1.4 PRRSV的ORF5基因的测序分析 参照GenBank中的18株PRRSV 参考序列(表1),利用Meg Align对阳性样品的ORF5序列进行核苷酸及推导氨基酸序列的同源性分析。利用 MEGA5.1对分离株 ORF5序列进行遗传演化分析。

表1 同源性序列分析用参考毒株名称及来源Table 1 The reference strains for blast analysis in this study
1.4 PRRSV缺失毒株的全基因组扩增及序列分析 针对类NADC30 PPRSV缺失TJjh1602株,参考文献[10]合成10对引物进行全基因组扩增,利用Megalign和Seqman对序列进行拼接;利用Megalign软件中的Cluster W程序对TJjh1602株全基因组序列进行分析;参照GeneBank中的13株参考序列(表1),利用MEGA5.1对TJjh1602株的全基因组序列进行遗传演化分析。
2 结果
2.1 病料样品中PRRSV的检测结果 分别提取采集疑似样品的RNA,进行反转录制备cDNA,以此为模板,利用设计的ORF5检测引物进行扩增,结果显示:106份样品呈PRRSV阳性,总阳性率为44.35%(106/236),部分阳性样品电泳结果见图1。分别将106份阳性PCR产物纯化后克隆至pMD18-T载体,将获得的阳性质粒送生工生物工程(上海)有限公司测序。
2.2 PRRSV的ORF5基因及氨基酸序列分析 将106株PRRSV的ORF5基因序列整理后进行核苷酸序列分析。结果显示:56株的ORF5 核苷酸基因与NADC30株的核苷酸序列同源性为98.8%~99.5%,表明56株PRRSV分离株为PRRSV类NADC30毒株,其中有18份流行毒株在ORF5基因99~101位(位置参考VR2332)缺失3个核苷酸,表现出相同特征性的核苷酸缺失模式,其余50株为经典PRRSV和HPPRRSV。氨基酸序列分析与核苷酸相同,18份样品在34位缺失1个氨基酸(位置参考VR2332 GP5氨基酸序列),与部分代表毒株的氨基酸位点分析如图2。
2.3 类NADC30毒株(TJjh1602)分离鉴定及全基因组扩增结果 针对性选取上述ORF5基因缺失的新鲜组织样品,研磨处理后分别接种Marc-145和PAMs细胞进行病毒分离,盲传3代后,细胞培养物经RT-PCR检测,成功分离到1株PRRSV类NADC30 TJjh1602株,该病毒不能在Marc-145细胞上增殖,仅感染PAMs细胞,且具有典型的CPE;对分离毒株TJjh1602进行噬斑纯化后,提取RNA反转录制备cDNA,进行分段扩增全基因组,结果显示,获得的PCR片段大小与预期大小相符(图3)。

图1 部分阳性样品ORF5基因的RT-PCR检测结果Fig.1 The RT-PCR detection results of ORF genes of the positive samples
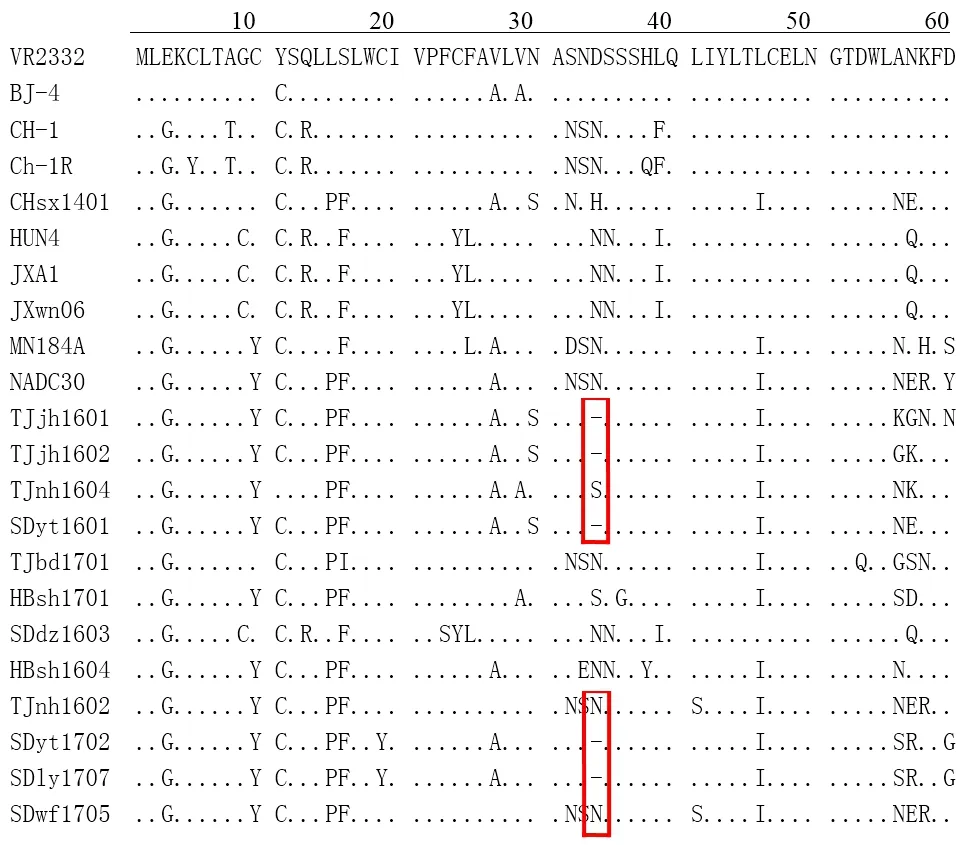
图2 2016-2017年华北地区PRRSV类NADC30毒株ORF5推导氨基酸序列比较Fig.2 Comparision of ORF5 amino acid sequences of NADC30-like strains in North China during 2016-2017
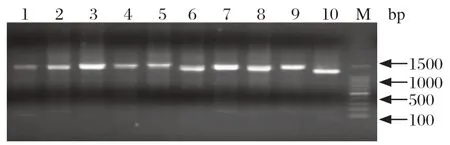
图3 PRRSV分离株TJjh1602全基因组各片段扩增结果Fig.3 The whole genome fragments amplification of PRRSV TJjh1602 isolate
2.4 类NADC30毒株TJjh1602核苷酸序列及推导氨基酸序列分析 对分离毒株TJjh1602的NSP2基因序列分析表明,在NSP2区存在131(111+1+19)个不连续氨基酸缺失,与NADC30毒株缺失序列一致;同源性分析结果表明,TJjh1602株的NSP2基因与NADC30毒株核苷酸和推导氨基酸同源性分别为92.2%和90.3%,ORF5基因与NADC30的核苷酸及氨基酸序列同源性分别为92.5%和91.4%。将分离毒株TJjh1602株的10条相互重叠的片段拼接成PRRSV全基因组,与NADC30株进行核苷酸及推导氨基酸序列的比较分析,结果显示,TJjh1602全基因长为15 018 nt,除缺失GP5 34位氨基酸外,无其他氨基酸插入或缺失,与NADC30株核苷酸序列同源性为95.2%。对NSP2基因、ORF5基因和全基因进化树分析结果表明,TJjh602毒株与NADC30毒株相近且位于系谱1(lineage 1)。
2.5 类NADC30 PRRSV的遗传演化分析 利用MEGA 5.1对56株分离株的ORF5 序列进行遗传演化分析显示,该56株PRRSV与NADC30毒株同属于一个分支,都属于谱系1(图4A);分离毒株TJjh1602与13株参考PRRSV毒株全基因组分析结果显示,分离毒株TJjh1602全基因组遗传进化与ORF5基因遗传演化分析结果一致,均属于NADC30分支(图4B)。
3 讨论
PRRS是危害我国生猪养猪业重要猪传染病之一,该病在我国猪群中已广泛流行。PRRSV具有毒株多样性和变异性强的特点,尤其流行新毒株不断出现,极易造成PRRS的大面积暴发。自2014年来国内学者检测到大量的在NSP2基因具有与NADC30同样的缺失特征PRRSV变异株[5-7,11],这类病毒可能已经成为主要流行毒株。因此,有必要对我国猪场的PRRSV类NADC30毒株的流行情况和相关基因型变化进行持续监测。
GP5是PRRSV表面主要的囊膜蛋白之一,存在遗传多样性[12-13]。因此,可通过监测ORF5基因的变异情况来分析PRRSV流行病学。本研究通过对2016-2017年间收集自我国华北四省市的236份PRRSV疑似样品进行RT-PCR检测,共检出PRRSV阳性样品106份,总阳性率达44.35%;通过对ORF5核苷酸序列分析发现,56株PRRSV分离株为PRRSV类NADC30毒株,为主要优势流行毒株,其余50株为经典PRRSV和HP-PRRSV。通过监测2015-2016年华东和华南地区的ORF5基因变异情况来对PRRSV进行分子流行病学分析,当前流行的PRRSV以HPPRRSV为主,但NADC30-like PRRSV毒株流行趋势明显增强[7]。由此可以推断,PRRSV类NADC30毒株已成为我国华北、华南、华东地区部分猪场优势流行毒株。
PRRSV的遗传变异一直是研究热点,而且有很多关于PRRSV变异的研究报道。在PRRSV的基因组中,ORF1a的NSP1β和NSP2、ORF3和ORF5的变异性相对较大,其中NSP2是突变率最高的部分,表现为基因缺失或插入[14],但鲜有关于ORF5基因缺失或插入报道。本研究通过对ORF5核苷酸及推导氨基酸序列分析发现,56份PRRSV类NADC30流行毒株中有18份存在GP5蛋白34位氨基酸的缺失模式,但在经典PRRSV和HP-PRRSV的GP5不存在氨基酸缺失;进一步对缺失毒株全基因组遗传进化分析表明,除在GP5 34位氨基酸缺失外,无其他氨基酸插入或缺失,且全基因进化树分析结果表明,缺失毒株与NADC30位于同一分支,属于系谱1(lineage 1)。以上研究结果表明,具有该新型缺失分子标签的流行毒株仅发生在类NADC30毒株,不存在经典PRRSV和HP-PRRSV,属国内首次报道。
由上可知,2016-2017年我国华北地区PRRSV流行毒株存在基因多样性,NADC30-like已成为优势流行毒株,且在遗传演化过程中出现一类具有新型ORF5缺失分子标签的毒株,这些新型变异毒株的流行病学特点、遗传变异情况以及临床的致病力等还有待于进一步研究证实,因此,有必要实时监测其流行情况,为临床疫病预警和研发更高效、安全的新型疫苗提供理论依据。
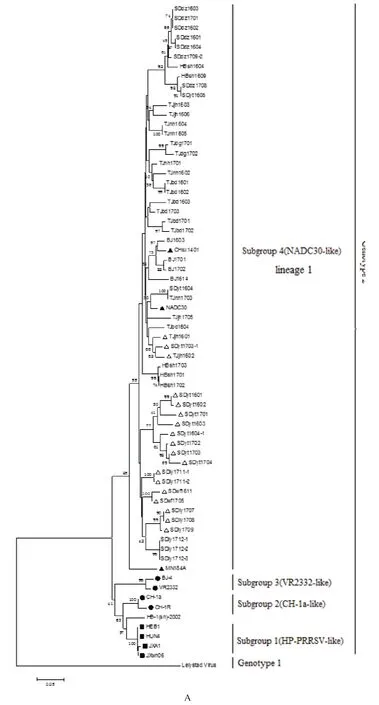

图4 基于ORF5基因和全基因序列的遗传进化树分析Fig.4 Phylogenetic tree based on the detected PRRSV isolates and reference strains
猜你喜欢
动物医学进展(2022年9期)2022-11-26
军事文摘(2022年16期)2022-08-24
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
