
一株野鸟源重组禽流感(H6N1)病毒基因组分子特征分析
2020-04-30 04:32杨晓宇吕欣孺陈丽霞天马沛然王玉龙侯志军曾祥伟王亚君杨思远张国钢柴洪亮
野生动物学报 2020年2期
杨晓宇 孙 静 吕欣孺 李 祥 陈丽霞 付 恬 马 天马沛然 李 仪 王玉龙 侯志军 曾祥伟 王亚君 杨思远张国钢* 孙 颖* 柴洪亮*
(1.东北林业大学野生动物与自然保护地学院,哈尔滨,150040;2.全国鸟类环志中心,北京,100091;3.黑龙江民族职业学院,哈尔滨,150066)
禽流感病毒(avian influenza virus,AIV)是属于正黏病毒科(Orthomyxoviridae)甲型流感病毒属(InfluenzavirusA)的甲型单链负向RNA病毒[1]。根据血凝素(HA)和神经氨酸酶(NA)糖蛋白的抗原性,将甲型流感病毒分为18种HA和11种NA亚型[2]。野生水禽,尤其是雁鸭类,被认为是AIV的天然宿主[3]。感染低致病性禽流感病毒的鸟类,包括家禽和野生鸟类,通常不会表现出临床症状,但是,它们为低致病性禽流感病毒间的基因重组提供了环境,而低致病性禽流感病毒可以作为高致病性禽流感病毒的前体[4]。禽流感病毒对家禽受体的适应可以增强其在人与禽之间传播的潜力,进而使它获得跨物种传播的能力[5]。近年来,禽类中陆续出现了不同的流感病毒亚型(例如H5,H6,H7和H10),它们越过了物种屏障并引起了人类感染的频繁暴发。禽流感的持续暴发已经引起人们的警觉,野生鸟类作为禽流感病毒主要储存库的作用越来越受到重视,它们保持了禽流感病毒的持久性和变异性,促进了病毒的传播和疾病的发生。
H6亚型禽流感病毒首次于1965年在马萨诸塞州的火鸡(Meleagrisgallopavo)体内分离到[6],目前已经在全球范围内流行,且宿主范围也较为广泛,包括各种水禽、家禽以及哺乳动物等。日前,在荷兰3个农场的鸡群中暴发H6N1亚型禽流感病毒已造成巨大的经济损失[7]。在中国,已有血清学证据表明人类以及其他哺乳动物都曾感染H6亚型AIVs[8],在台湾也已有人感染H6N1亚型流感病毒并引起明显流感症状的确诊病例[9],之前的研究还表明H6N1病毒有7个基因片段与1997年香港暴发的H5N1病毒高度同源[10]。虽然目前还未发现H6亚型禽流感病毒引起类似于H5、H7亚型病毒的高致病性和大面积暴发,但是它们的普遍存在和致病力增强的潜在可能一直令人担忧,特别是能否获得对哺乳动物的跨种感染能力与传播能力。
本试验对从江苏省海丰农场豆雁(Anserfabalis)粪便中分离的1株H6N1亚型毒株J746进行全基因克隆及遗传进化分析。
1 材料和方法
1.1 样品采集
本课题组于2017年1月19日在江苏省海丰农场小麦地采集新鲜豆雁粪便样品159份。用棉签蘸取少量粪便放入5 mL EP管中,内含2 mL病毒保存液(PBS缓冲液、青霉素、链霉素和10%甘油),-80 ℃储存备用。
1.2 病毒分离鉴定
将上述159份粪便样品震荡混匀后静置10 min,取0.1—0.2 mL上清液由尿囊腔途径接种2枚9日龄SPF 鸡胚,37 ℃全自动孵化器内孵育72 h后在无菌环境中收取尿囊液6—8 mL,分装于4个2 mL EP管中,取其中一管进行血凝(HA)实验。
HA呈阳性的尿囊液样品,按照QIAGEN公司的RNA提取试剂盒说明书要求对其进行总RNA的提取,随后用Unit12引物(5′-AGC AAA AGC AGG-3′)进行反转录,作为PCR模板。
利用特异性HA-640引物进行RT-PCR检测和HA亚型鉴定[11],PCR产物在1%琼脂糖凝胶中进行电泳后,挑选出呈预期长度条带的核酸样品,使用特异性引物进行RT-PCR实验扩增全基因组片段。
1.3 测序和序列分析
基因组扩增的PCR产物送往吉林库美生物科技有限公司测序,返回的测序结果用DNAStar 软件包中的Seqman 程序进行拼接,利用MegAlign 程序进行核苷酸同源性比较。使用Mega 7.0 进行序列比对以及氨基酸序列分析,利用IQTREE软件分别对8个基因片段最优进化模型的筛选,随后构建最大似然法遗传进化树。
2 结果
2.1 全基因RT-PCR扩增及测序结果
从鸡胚尿囊液中提取病毒核酸,并进行RT-PCR扩增,电泳结果显示,各条带与预期片段大小相符。对返回测序进行拼接,J746分离株PB2、PB1、PA、HA、NP、NA、M、NS基因蛋白质编码区(CDS)长度为2 280 bp、2 274 bp、2 151 bp、1 701 bp、1 497 bp、1 410 bp、982 bp、844 bp,GISAID序列号分别为EPI1619579、EPI1619611-EPI1619617。
2.2 同源性比较结果
如表1所示,J746的HA基因与韩国野生水禽病毒A/wild waterfowl/Korea/F14-5/2016(H6N1)同源性最高,同源性为99.4%,NA同样与在韩国野生水禽中分离出的A/white-fronted goose/Korea/H909-1/2017(H6N1),A/white-fronted goose/Korea/H952/2017(H6N1)和A/wild waterfowl/Korea/F14-5/2016(H6N1)同源性最高,同源性均为99.4%。6个内部基因中,PB2、PB1和NP均与蒙古家鸭的低致病性毒株同源性最高,分别为99.7%,99.9%和100%;与PA同源性最高的毒株中,除了分离于蒙古家鸭的低致病性H10N2毒株,还有分离于日本家鸭的低致病性H3N8毒株,同源性均为99.4%,与M同源性最高的毒株中,除了分离于蒙古家鸭的低致病性H4N6毒株,还有分离于荷兰野鸭的低致病性H4N6毒株,同源性均为99.7%,与NS同源性最高的为分离于中国武汉的家禽源A/chicken/Wuhan/WHJF/2014(H5N2)毒株,同源性为100%。
2.3 遗传进化分析结果
2.3.1 外部基因
如图1和图2所示,J746的HA基因与分离于韩国、日本野生水禽的低致病性H6N1、H6N8亚型毒株和孟加拉家鸭的低致病性H6N2亚型毒株聚集在一个分支上,其中与A/wild waterfowl/Korea/F14-5/2016(H6N1)亲缘关系最近。NA基因与分离于韩国野生水禽的低致病性H6N1、H7N1亚型聚集在一个分支上,同样与A/wild waterfowl/Korea/F14-5/2016(H6N1)的亲缘关系非常近。因此J746的HA和NA基因可能共同起源于韩国病毒A/wild waterfowl/Korea/F14-5/2016(H6N1)。
2.3.2 内部基因
如图3—图8所示,PB2基因与蒙古家鸭低致病性H10N3毒株、中国重庆家鸭H5N3毒株以及哈萨克斯坦野鸭H1N1毒株聚集在一个分支上。PB1基因和NP基因都与循环于东南亚的低致病性毒株处于同一分支,其中PB1基因与蒙古家鸭的H1N1 毒株A/duck/Mongolia/520/2015(H1N1)亲缘关系最近,NP基因与蒙古家鸭的A/duck/Mongolia/208/2015(H3N8)等H3N8毒株亲缘关系最近。PA基因和M基因都处于欧亚分支上,值得注意的是PA基因,虽然处于欧亚大分支上,但形成了独立的小分支,从进化关系上看,虽然PA基因与日本家鸭的A/duck/Fukui/181019/2016(H3N8)和A/duck/Yamaguchi/351201/2016(H4N6)两株毒亲缘关系最近,但可能经过了相对独立的进化过程。NS基因与分离于中国中南部和日本的多种亚型低致病性毒株聚集在一起,其中与中国安徽野生水禽的H6N2和H1N1两种亚型的分离株亲缘关系最近。
2.4 氨基酸位点分析结果
如表2所示,HA基因开放阅读框架有1 701个碱基,编码566个氨基酸,其裂解位点为PQIETR↓G,无连续碱性氨基酸的插入,表明该毒株符合低致病性禽流感病毒的典型特征[12]。第239(H3-226)位和241(H3-228)位氨基酸分别为Q和G,说明J746优先结合唾液酸α2-3 受体[13]。NA蛋白无颈部氨基酸缺失,但其出现H274Y突变,表明J746对神经氨酸酶类抗病毒药物具有一定的耐药性[14]。PB2蛋白中出现与增强对小鼠致病性有关的L89V突变[15],NS1蛋白中出现与增强对小鼠致病性、提高复制能力和改变宿主嗜性有关的P42S、L103F、I106M、N205S突变[16-17]。
表1 J746各基因片段的核苷酸同源性比较

Tab.1 Comparison of nucleotide homology of each gene fragment of J746
3 讨论
野生水禽是A型流感病毒重要的自然宿主,其每年规律性的迁徙活动对流感病毒的长期存在、持续变异和广泛传播起着十分重要的作用。在候鸟的8条主要迁徙路线中有3条经过我国,几乎覆盖了我国的全部领域,各条路线之间还有相当复杂的迁徙交汇区,更易造成不同鸟类种群之间的跨种传播和接力式、远距离跨洲传播。
本研究对2017年在江苏海丰农场采集的野生豆雁粪便中分离到的H6N1病毒株J746进行了全基因组测序,并对其进行了序列分析和遗传进化分析。结合同源性和遗传进化分析结果,J746的HA和NA基因均可能来源于韩国野鸟源H6N1病毒株A/wild waterfowl/Korea/F14-5/2016(H6N1)。内部基因来源较复杂,PB2基因可能来源于亚洲中东部地区低致病性H1、H5或H10毒株;PB1和NP基因可能分别源于蒙古家鸭低致病性H1N1、H3N8毒株;PA和M基因均源于欧亚低致病性毒株但PA基因进化过程相对独立;NS基因与中国安徽野生水禽的H6N2和H1N1两种亚型的分离株亲缘关系最近。虽然J746各基因片段的进化路径不尽相同,但它们的来源在总体上呈现一定的区域性,因此推测J746是由循环在亚洲及欧洲的低致病性毒株在候鸟沿东亚—澳大利亚迁徙路线进行迁徙或停歇时发生重组的。
此外,事实已经证明,禽流感的每次暴发都会导致家禽养殖业的巨大经济损失,J746虽然分离于野生水禽,但其所有的内部基因均与家禽源AIV毒株具有高度的同源性和密切的遗传关系,说明野生水禽在迁徙过程中与家禽发生了密切的基因交流,这大大提高了家禽养殖业的风险和AIV防控的难度。并且J746已经产生了多个与增强致病性和改变宿主适应性有关氨基酸突变位点,不排除其存在感染哺乳动物的可能。因此,加强对流感病毒的监测,尤其是对野生水禽携带AIV的主动预警监测研究,无论是对维护社会经济稳定,还是对提高社会公共卫生水平,都具有重要意义。
表2 J746氨基酸位点分析

Tab.2 Amino acid site analysis of each gene fragment of J746
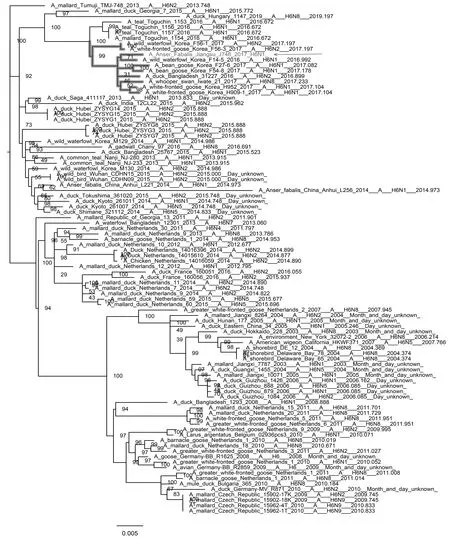
图1 HA基因进化树Fig.1 Phylogenetic tree of HA gene 注:图中加粗部分为J746的HA基因所在分支,箭头指向J746分离株 Note:The bold part in the figure is the branch of the HA gene of J746,and the arrow points to the J746 isolate
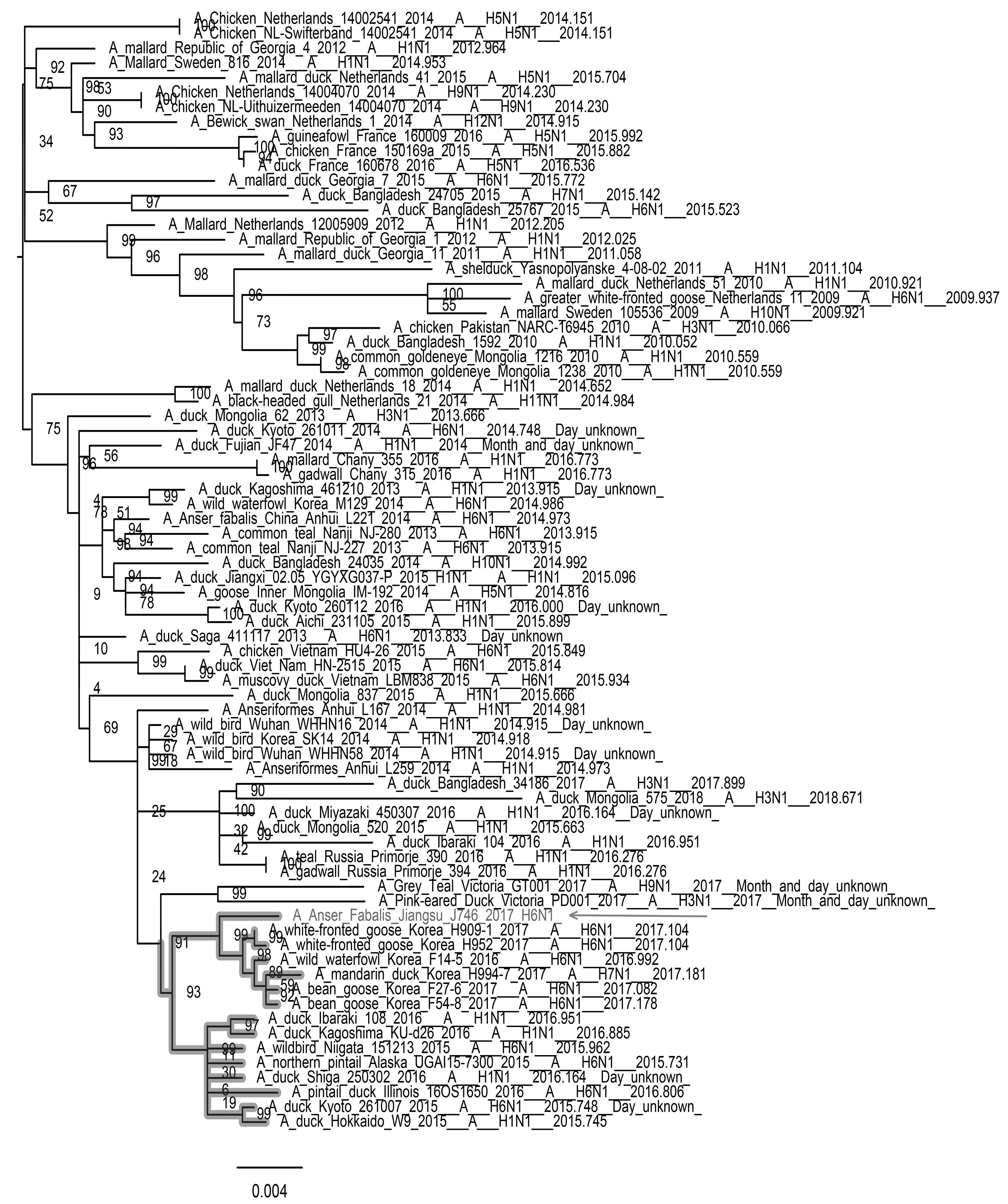
图2 NA基因进化树Fig.2 Phylogenetic tree of NA gene 注:图中加粗部分为J746的NA基因所在分支,箭头指向J746分离株 Note:The bold part in the figure is the branch of NA gene of J746,and the arrow points to the J746 isolate

图3 PB2基因进化树Fig.3 Phylogenetic tree of PB2 gene 注:图中加粗部分为J746的PB2基因所在分支,箭头指向J746分离株 Note:The bold part in the figure is the branch of PB2 gene of J746,and the arrow points to the J746 isolate
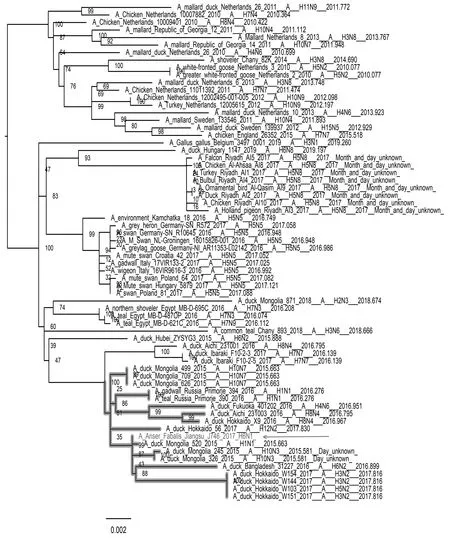
图4 PB1基因进化树Fig.4 Phylogenetic tree of PB1 gene 注:图中加粗部分为J746的PB1基因所在分支,箭头指向J746分离株 Note:The bold part in the figure is the branch of PB1 gene of J746,and the arrow points to the J746 isolate
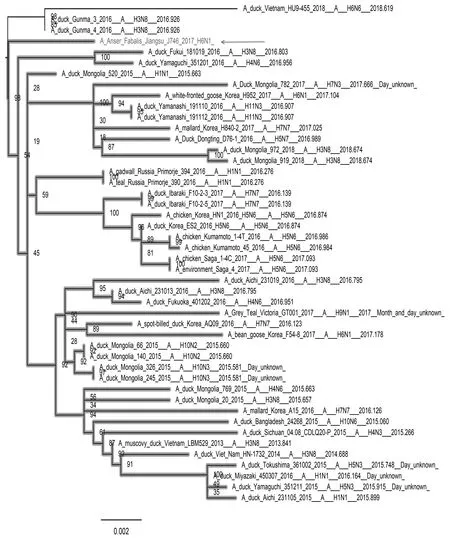
图5 PA基因进化树Fig.5 Phylogenetic tree of PA gene 注:图中加粗部分为J746的PA基因所在分支,箭头指向J746分离株 Note:The bold part in the figure is the branch of PA gene of J746,and the arrow points to the J746 isolate
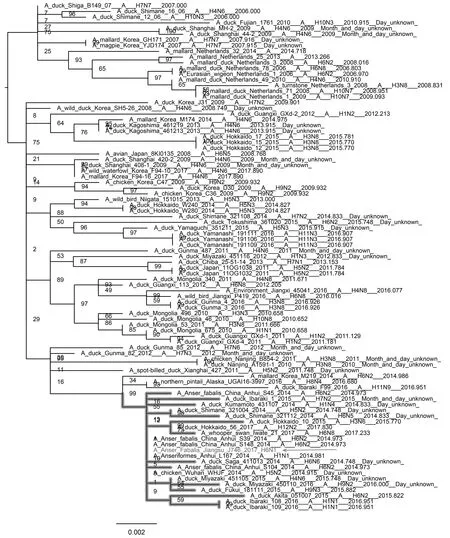
图8 NS基因进化树Fig.8 Phylogenetic tree of NS gene 注:图中加粗部分为J746的NS基因所在分支,箭头指向J746分离株 Note:The bold part in the figure is the branch of NS gene of J746,and the arrow points to the J746 isolate
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
奇妙博物馆(2022年9期)2022-09-28
小哥白尼(野生动物)(2022年6期)2022-08-17
中国土壤与肥料(2021年5期)2021-12-02
教师·中(2018年4期)2018-06-02
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
中国动物保健(2015年4期)2015-10-21
大众考古(2015年4期)2015-06-26
