
Komagataeibacter rhaeticus 3-15全基因组及葡萄糖脱氢酶基因缺失体的构建
2020-04-27 08:14李萧,王淼
工业微生物 2020年2期
李 萧,王 淼
江南大学食品学院,江苏 无锡 214122
细菌纤维素(Bacterial Cellulose,BC),是醋酸菌分泌合成的胞外多糖,由葡萄糖单体分子以β-1,4-糖苷键聚合而成的高分子聚合物,是一种天然形成的最细纤维[1,2]。与植物纤维素相比,BC具有均一性好、纯度高等[3]特点,可广泛用于医疗、影像器材、食品等众多领域[4]。如今,合成生物学的发展使模型微生物可以用模块化的DNA代码进行编程,以便执行各种各样的操作来达到目的[5]。这种方法可以通过基因调控来对BC合成进行更精细的调控。然而,关于BC生产菌株的基因信息较少,相关的基因改造几乎没有[6]。因此,获取各种BC生产菌株的基因组信息至关重要。
到目前为止,几种产BC的Gluconacetobacter和Komagataeibacter已被测序,如G.xylinusE25[7],K.medellinensisNBRC 3288[8],K.nataicolaRZS01[9]和GluconobacteroxydansDSM 3504。其中Gluconacetobacterxylinus(又称Acetobacterxylinum)是最常被研究的物种之一,因为它可以从各类的碳、氮源中产生大量的BC[10-12],对Komagataeibacter的研究较少。在本研究中,我们首先提出了K.rhaeticus315的完整基因组序列,为后续基因工程提供背景信息,后续可在代谢途径的基础上实现对BC生物合成的精确控制。其次,结合基因组序列,本研究对K.rhaeticus315的代谢途径及代谢物进行分析。最后,本研究通过对K.rhaeticus315进行分子改造,将葡萄糖合成葡萄糖酸途径中的关键酶-葡萄糖脱氢酶的相关基因敲除,使葡萄糖主要流向BC合成途径,降低副产物的生成。
1 材料与方法
1.1 菌种与培养基
莱蒂亚驹形杆菌(Komagataeibacterrhaeticus)3-15:本实验室从红茶菌中筛选并保藏。保藏号:CCTCC NO: M2016213。
种子培养基(g/L):葡萄糖20、蛋白胨5、酵母粉5、Nacl 5,115 ℃灭菌20 min后室温保存待用(固体培养基另加2%琼脂)。
发酵培养基(g/L):葡萄糖30、蛋白胨5、酵母粉5、2%(v/v)无水乙醇,115 ℃灭菌20 min后室温保存待用。
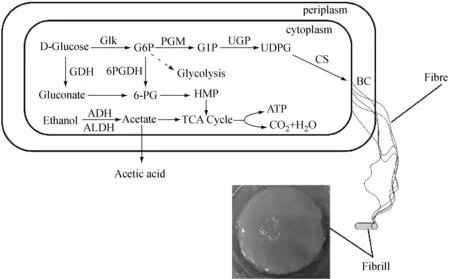
图1 细菌纤维素生物合成途径
1.2 试剂与设备
用于PCR反应的2×PrimeSTAR Max DNA高保真聚合酶、2×Taq低保真聚合酶均购买于宝生生物有限公司(大连,中国);Ezup柱式细菌基因组小量提取试剂盒、卡那青霉素(Km)以及琼脂糖均购于上海生工;标准样品葡萄糖酸(GA)和乙酸购自Sigma-Aldrich有限公司(美国);Tryptone、Yeast Extract够买自Oxoid公司(美国);Gold view、DNA Marker购自广州东盛生物科技有限公司(广州,中国);其他所需试剂均为国产级分析纯试剂。
PCR仪:美国Bio-Rad公司;DYY-60D 型核酸电泳仪:北京六一电泳仪厂;Gel Doc 凝胶成像系统:美国Bio-Rad公司;蛋白核酸定量测定仪:美国Thermo scientific公司;高效液相安捷伦1260色谱仪:美国Agilent公司;高速低温离心机:德国Eppendorf公司;恒温摇床:上海跃进医疗器械厂;ZXSD-B1160生化培养箱,上海智城分析仪器制造有限公司。
1.3 实验方法
1.3.1DNA提取及序列分析
将菌体在种子培养基里活化完成后,取培养液5 mL,用试剂盒提取DNA,电泳检测合格后,构建测序文库。取100 ng全基因组DNA,使用超声波破碎仪将其随机打断成小于500 bp的片段,然后进行末端修复,两端加测序接头,最后用P5和P7引物扩增纯化后目的片段。使用Agilent 2100生物分析仪检测文库质量,并且检测文库浓度,用检验合格的文库进行cluster制备和测序。取5 μg~10 μg高质量的基因组DNA,随机打断成10 kb左右的片段(根据基因组大小选择20 kb文库),随后用DNA Template Prep试剂盒构建SMRTbell文库。结合测序引物和聚合酶,以自由扩散的方式加入到PacBio Sequel平台的测序芯片中,准备测序。最后,对于质检合格的文库,根据项目数据量要求进行上机测序。
1.3.2基因组组装与注释
首先,基于第三代测序数据,使用HGAP4或Falcon软件获得初步装配结果。然后,将经过质量筛选的第二代测序数据与装配结果进行对比,通过软件Pilon对装配结果进行进一步校正,得到最终的装配结果。
基因功能注释主要的方法来预测基因编码蛋白序列与数据库包含蛋白质序列比对,如果某一个基因的蛋白质序列和蛋白质序列数据库中具有显著的序列相似性,可以推断出,基因和蛋白质数据库中相同或相似的功能。根据编码基因的预测蛋白序列,使用BLAST等比对软件与数据库中的蛋白序列进行比对。序列比对值设置为1e-5,序列比对长度不小于蛋白质长度的60%。选择最优匹配结果作为该基因的注释结果。
1.3.3葡萄糖脱氢酶基因的敲除
(1) 感受态的制作
①将-80 ℃保存的醋酸杆菌K.rhaeticus315进行平板划线,待长出单菌落后,挑取单菌落至3 mL种子培养基中过夜培养。
②取0.5 mL活化液接种至50 mL 种子培养基中,培养至OD600=0.5~0.6时,即达到制备感受态的要求。
③将培养液转入无菌离心管中,冰浴10 min~15 min,于4 ℃,4 500×g离心10 min收集菌体。
④弃上清液,用1 mL预冷的0.1 mol/L氯化钙溶液轻轻重悬菌体,于4 ℃,4 500×g离心10 min,弃上清。
⑤用1 mL预冷的超纯水重悬菌体,于4 ℃,4 500×g离心10 min,弃上清。此步骤重复两次。
⑥用1 mL 10%甘油重悬菌体,于4 ℃,4 500×g离心10 min,弃上清。
⑦按每管60 μL分装至冰上预冷的1.5 mL无菌离心管中可直接使用或冻存于-80 ℃。
(2) 电脉冲转化法
①将电脉冲杯从75%无水乙醇中取出,吹干,置于冰上预冷。
②从-80 ℃冰箱中取出感受态细胞,置于冰上融化后,加入10 μL片段。
③打开电脉冲仪,设置好参数(电压为2 kV),将混有片段的感受态细胞加入到预冷的电脉冲杯里。
④将电脉冲杯放入电脉冲仪中,按下“开始”按钮,当听到“叮”的一声后,立即取出电脉冲杯并迅速加入1 mL种子培养基,混匀。
⑤在30 ℃下缓慢振荡培养2 h,取100 μL涂布到卡那抗性平板上,30 ℃倒置培养。
(3)gcd基因的构建与克隆
根据gcd基因序列及km基因序列设计PCR引物,如表1所示。分别扩增出gcd基因的上臂、km片段、下臂并将其连接起来,采用电脉冲法对K.rhaeticus315感受态进行转化后用验证引物验证是否转化成功。
(4)gcd-重组菌株的发酵
将转化成功的菌株进行发酵,30 ℃下静置培养4 d,测BC干重及葡萄糖酸浓度。
1.4 分析方法
1.4.1BC干重的测定[13]

表1 PCR引物序列
取发酵后的培养液,经纱布过滤,将过滤后的BC用清水冲洗然后浸泡到1.5%(w/v)NaOH溶液中,80 ℃水浴2 h,用去离子水冲洗至pH中性,放入称量皿中,于105 ℃烘箱中烘干至恒重,称量,得BC的干重。
1.4.2残糖含量的测定
使用生物传感分析仪对发酵液中的葡萄糖浓度进行测定。
1.4.3葡萄糖酸、乙酸的测定
葡萄糖酸、醋酸的测定采用HPLC方法[14]。
样品前处理:培养液离心后过0.22 μm水膜过滤,待测。色谱条件:高效液相色谱Agilent 1 260,检测器为DAD,色谱柱:SB-AQ。流动相:0.025 mol/L磷酸二氢钾溶液,pH 2.5。流速:0.5 mL/min。检测器波长:210 nm,柱温:30 ℃,进样量:10 μL。
1.5 数据处理分析
采用Origin 2016进行数据分析,结果用均值±标准偏差来表示。
2 结果与讨论
2.1 K.rhaeticus 315全基因组测序
K.rhaeticus315是新筛选的一株BC生产能力较强的菌株,为了最大程度的发挥BC生产菌株的生产能力,了解与BC合成相关的基因,本研究对K.rhaeticus315的全基因组进行了测序注释。如表2所示,K.rhaeticus315的基因信息被组装成一条序列,总长度为3.70 Mbp,平均覆盖率为470X,GC含量63.98%。预测编码DNA序列(CDS)数为3 314条。K.rhaeticus315基因组完整圈图如图2所示。
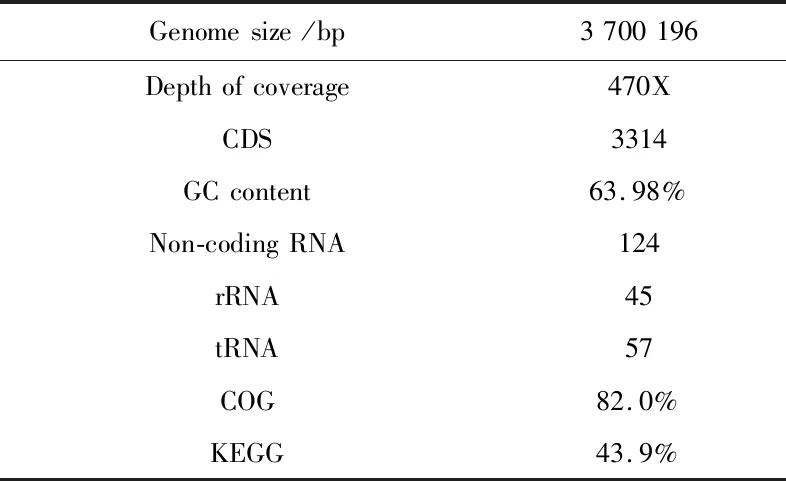
表2 K.rhaeticus 315基因组功能
图3是菌株的蛋白质功能图。在K.rhaeticus315基因组中,包含25个转运蛋白基因、9个同工基因和37个透性酶基因。他们主要负责运输糖、糖酸、氨基酸、磷酸盐、金属离子和脂多糖等。K.rhaeticus315可以利用多种碳源去合成BC,最常见的为葡萄糖。一般情况下,葡萄糖会通过EMP途径代谢。由于K.rhaeticus315基因组中缺乏编码磷酸果糖激酶的基因(PFK;EC 2.7.1.11),所以该菌株不能通过完整的EMP途径去代谢葡萄糖供能。这与类似菌株的研究结果一致[15,16]。此外,BC合成过程中另外两个关键酶是葡萄糖激酶(glk)和丙酮酸激酶(pyk)均存在于K.rhaeticus315基因组中,它们的编码基因座标记为WP_019084814.1和WP_110095141.1。
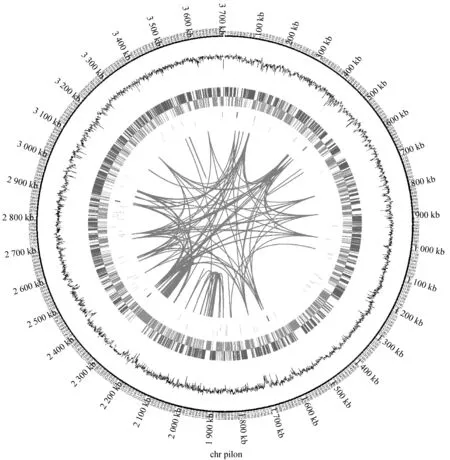
图2 K.rhaeticus 315基因组圈图
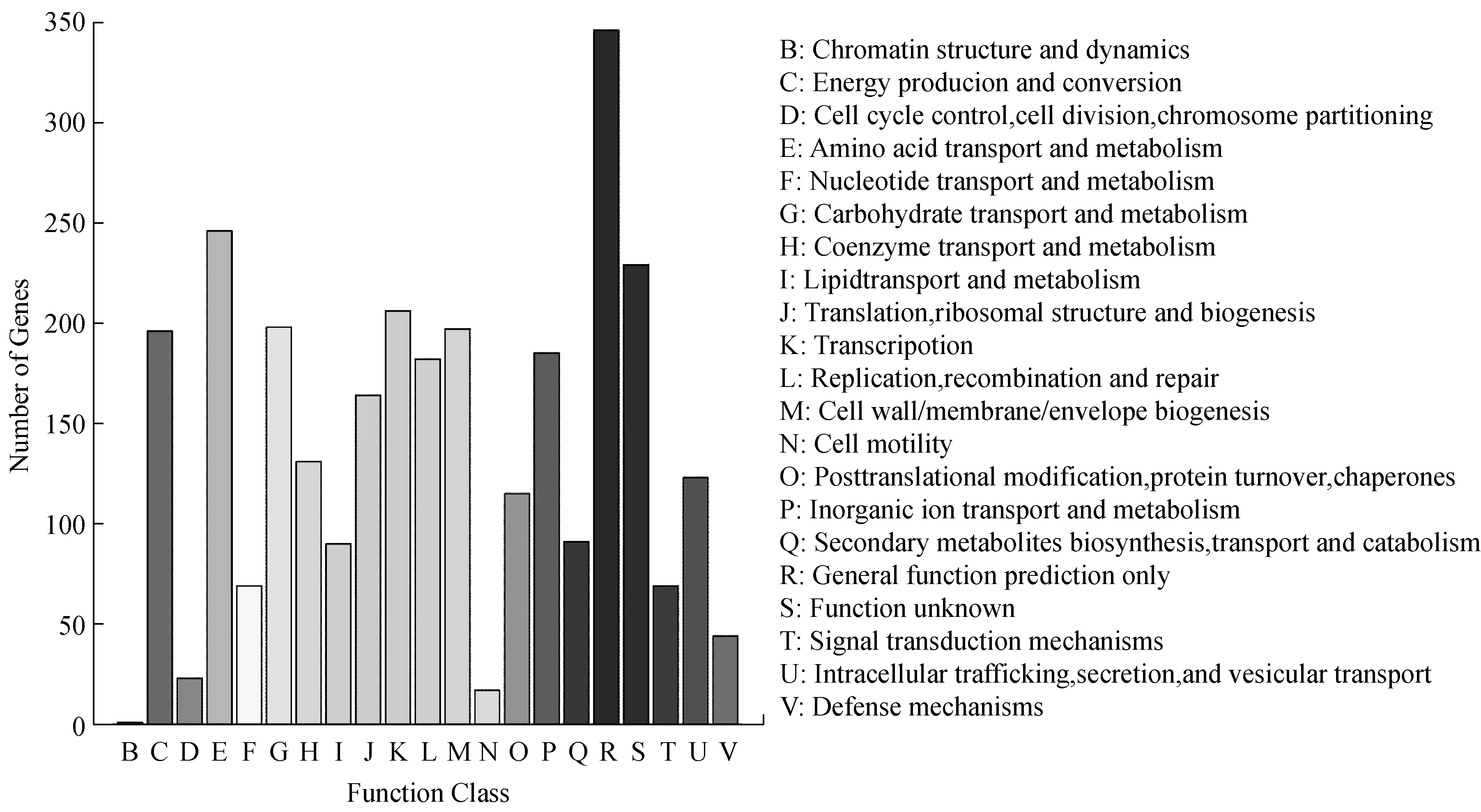
图3 蛋白质功能的同源群(COG)分类
戊糖磷酸途径(PPP)是K.rhaeticus315中另一种有效的葡萄糖代谢途径,在K.rhaeticus315基因组存在编码PPP途径中的关键酶基因:葡萄糖-6-磷酸脱氢酶的编码基因是gcd(WP_110094965.1和WP_003620475.1),6-磷酸葡糖酸内酯酶的编码基因是pgl(WP_110094851.1),6-磷酸葡萄糖酸脱氢酶的编码基因是pgd(WP_110094848.1),核糖磷酸3-表异构酶的编码基因是rpe(WP_110912266.1),转酮醇酶的编码基因是tktA,tktB(K00615)。有编码三羧酸循环(TCA)中三个关键酶的基因:柠檬酸合酶(K01647),异柠檬酸脱氢酶(K00031,K00030),2-氧戊二酸脱氢酶(K00164,K00658)。所以K.rhaeticus315可以通过PPP和TCA途径进行代谢来维持菌体生长和BC合成。
2.2 代谢产物分析
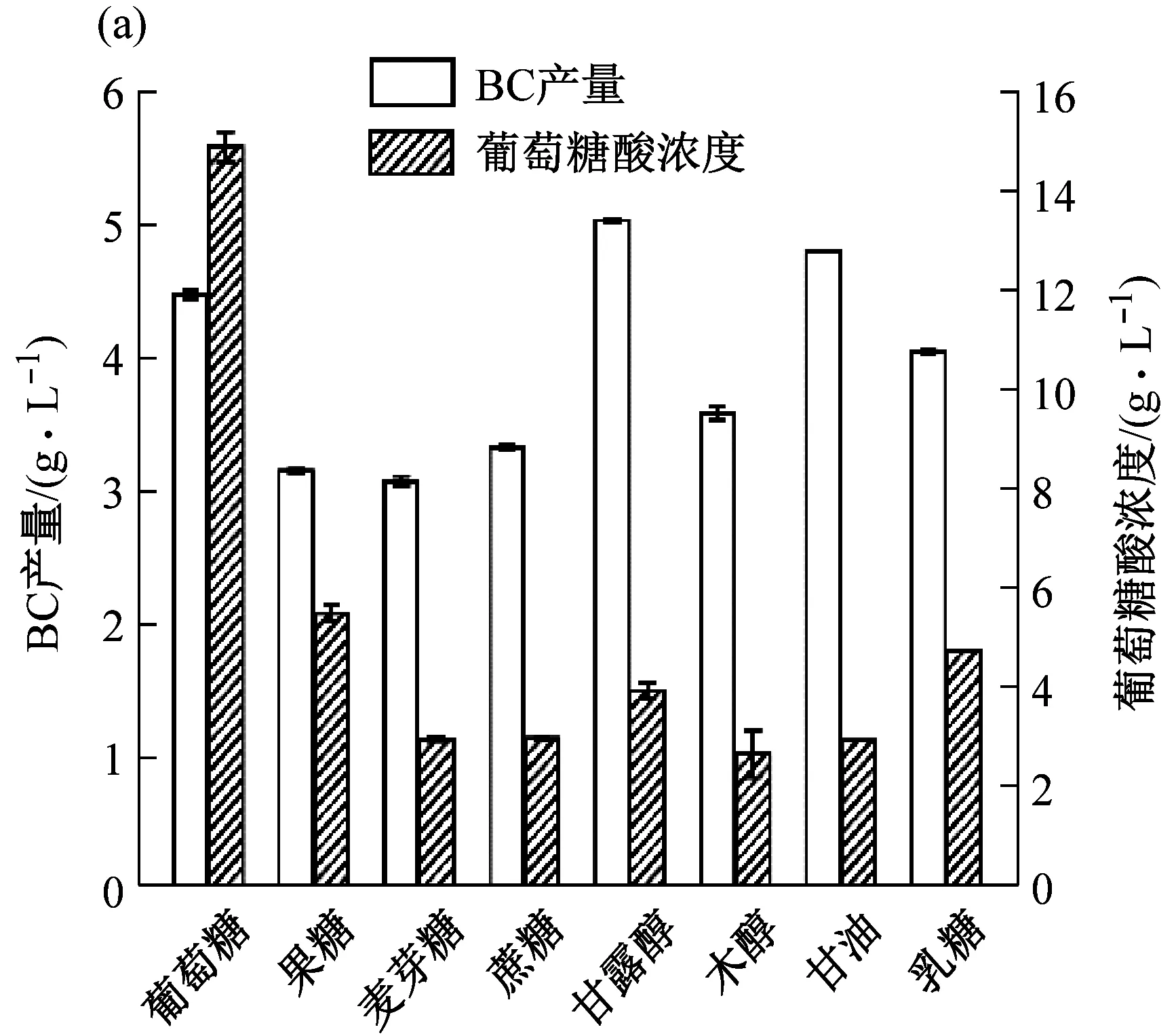
据报道,碳源是影响BC产量和结构差异的主要因素之一[17]。本研究发现K.rhaeticus315可以较好的利用葡萄糖、甘油和甘露醇,且三种碳源所合成的BC产量相似(图4a)。菌株对果糖的利用不好,从基因组中分析,由于不存在编码果糖激酶的基因(frk),所以果糖无法被转化为果糖-6-磷酸以及6-磷酸-葡萄糖。K.rhaeticus315对于双糖的利用也不佳,主要是因为K.rhaeticus315基因组中缺乏编码蔗糖酶、麦芽糖酶等双糖酶的基因。由于该菌株不具备EMP途径,菌体主要通过PPP途径和TCA途径来代谢碳源。此时碳源流向三个途径:菌体生长代谢、合成BC、合成酸类副产物。从图4b可以看出,发酵20 d后,耗糖量为36.73 g/L,BC产量为12.81 g/L,葡萄糖酸生成量为32.25 g/L。进一步说明菌体在合成BC时,有大量的葡萄糖酸副产物生成[18],造成碳源的浪费。并且,大量的酸会降低发酵液的pH,对菌体生长不利[19]。因此,对菌体进行改造,减少副产物的生成对提高BC产量至关重要。
2.3 K.rhaeticus 315葡萄糖脱氢酶基因缺失体的构建
葡萄糖酸(GA)是葡萄糖在葡萄糖脱氢酶(gcd,WP_110094965.1和WP_003620475.1)的作用下产生的。为了减少GA的生成,使葡萄糖代谢流向BC合成途径,本研究通过敲除GA合成关键酶的基因,来减少副产物GA的生成。首先选择敲除gcd(WP_003620475.1)这一基因,构建gcd-重组菌株。
2.3.1片段融合
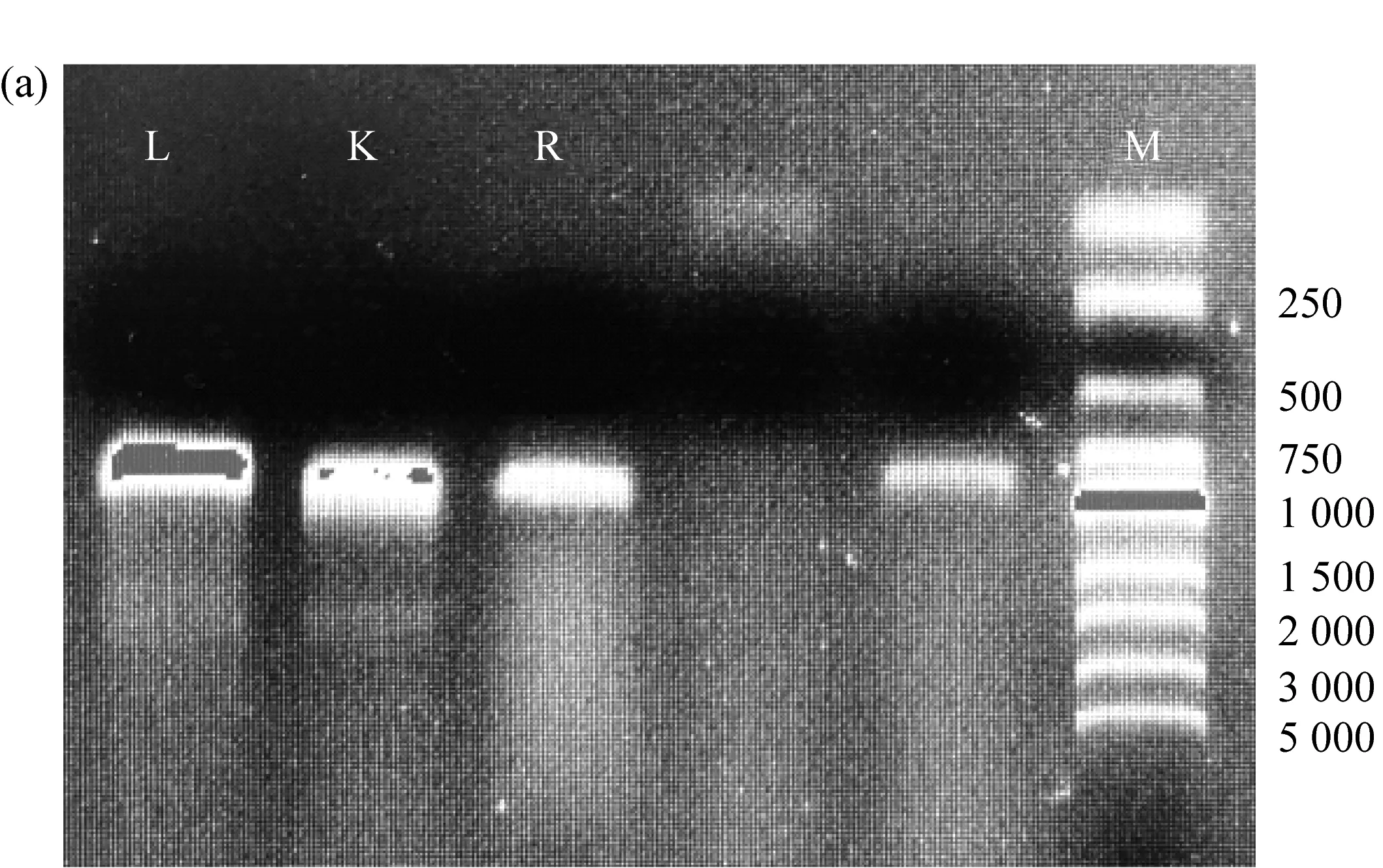
首先利用PCR分别扩增gcd基因(WP_003620475.1)的左臂、右臂以及Km基因。如图5a所示,电泳条带显示三段基因全都扩增成功,条带大小为900 bp。再将已扩增完成的三段基因融合。图5b说明三段基因连接成功,融合片段大小为2 500 bp。
2.3.2同源重组
采用电脉冲法,将2.3.1融合成功的片段转入K.rhaeticus315感受态中进行同源重组。利用卡那霉素抗生标记筛选目的菌株gcd-重组株,并用验证引物(表1)进行验证,验证片段的大小为1 000 bp左右。如图6所示,片段转化成功,即gcd-重组菌株构建成功。
2.3.3gcd-重组菌株的发酵
将未电转的原始菌株与gcd-重组菌株静置发酵4 d,观察BC干重及GA生成量。如表3所示,gcd-重组菌株的GA生成量显著下降,比原始菌株降低了7.93 g/L,降低了77%。但BC干重无明显提高,推测是单纯敲除一个gcd基因在有限的发酵时间内体现不出其节约BC合成前体—葡萄糖的优势;另一方面基因的敲除可能会影响菌体的其他代谢功能。
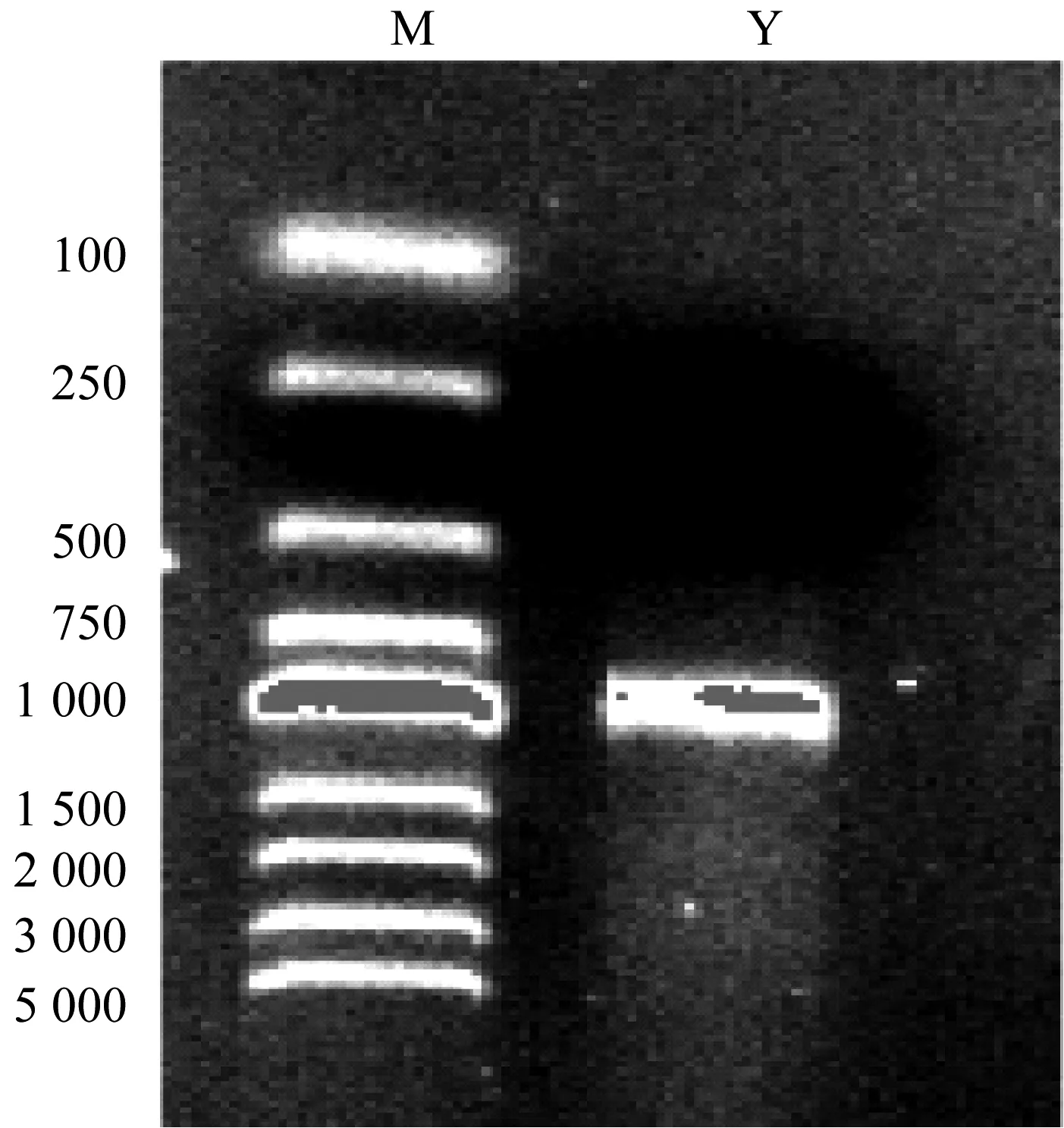
M: DNA marker;Y: 验证片段图6 验证片段琼脂糖凝胶电泳图

表3 gcd基因敲除后对发酵的影响
3 结论
本研究对新筛选的BC生产菌株K.rhaeticus315进行了全基因组测序及简单分析。通过基因信息及代谢途径分析,K.rhaeticus315是通过PPP和TCA途径来代谢碳源,维持菌体生长和代谢。当葡萄糖作为碳源时,菌体会产生大量的葡萄糖酸,造成葡萄糖的浪费。因此,通过全基因组序列找到葡萄糖合成葡萄糖酸途径中的关键基因gcd,对该基因进行敲除,可使葡萄糖酸降低了77%。本研究仅对K.rhaeticus315做了简单分析和改造,后续可利用该基因组信息进行进一步的分析和操作。
猜你喜欢
金属加工(热加工)(2022年7期)2022-07-12
当代水产(2022年1期)2022-04-26
今日农业(2021年11期)2021-08-13
电力大数据(2021年4期)2021-07-29
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
中成药(2018年7期)2018-08-04
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
创新科技(2014年12期)2014-07-27
