
ADV VP2主要抗原表位基因核酸疫苗与IL-12、IFN细胞因子佐剂质粒的构建及鉴定
2020-04-26 02:59柏玲朱言柱胡博朱翔宇张海玲廉士珍张蕾
特产研究 2020年2期
柏玲,朱言柱,胡博,朱翔宇,张海玲,廉士珍,张蕾※
(中国农业科学院特产研究所,吉林 长春 130112;2.农业农村部经济动物疫病重点实验室,吉林 长春 130112)
水貂阿留申病(Aleutian disease,AD)又称为浆细胞增多症,是由水貂阿留申病毒(Aleutian disease virus,ADV)引起的一种免疫抑制性疾病[1-2]。AD临床上以免疫复合物沉积导致的肾小球肾炎、高丙种球蛋白增多和浆细胞增多为主要特征[3-4];AD 广泛存在于世界各养貂国家和地区,在我国主要养貂地区感染率达70%以上,患病水貂消瘦,毛皮质量差,生育能力下降,严重危害水貂养殖业[5-6]。
ADV为无囊膜、二十面体、单股、线状DNA病毒,ADV属于细小病毒科、细小病毒亚科、阿留申病毒属[7]。ADV 全长4.8 kb,ADV 基因组主要有2 个开放阅读框(open reading frame,ORF):L-ORF 和 R-ORF。LORF 编码 3 种非结构蛋白(NS1、NS2、NS3),R-ORF编码 2 种结构蛋白(VP1 和 VP2)[8-10]。与 VP2 蛋白相比,VP1 在N 端多42 个氨基酸,VP2 占病毒衣壳的90%[11-12]。目前尚无疫苗防控AD,其主要原因是动物感染AD后,虽然会产生高水平抗体,但该抗体并无中和病毒的能力,反而会介导抗体依赖性增强作用(antibody dependent enhancement,ADE),进一步诱导病毒侵染机体,并且产生的抗原抗体复合物沉积,从而导致肾小球肾炎及动脉炎。研究者尝试制备ADV 灭活疫苗,虽然可以产生抗体,但会进一步增强病毒侵染。Aasted等[13]通过表达ADV 病毒结构蛋白VP2 及非结构蛋白NS1,将纯化的蛋白对动物进行免疫,结果发现,VP2免疫并无保护作用,反而导致动物死亡;NS1 免疫只有部分保护作用。近年来,核酸疫苗成为热点,核酸疫苗又称为基因疫苗,是将外源基因插入至真核表达载体中,构建成重组表达质粒,将质粒通过肌注等方式导入动物体内,使动物细胞表达外源蛋白,进而诱导机体产生免疫应答。核酸疫苗在机体内表达的蛋白更接近于天然状态,其免疫原性更好。与常规疫苗相比,它解决了主要组织相容性复合体(major histocompatibility complex,MHC)限制性问题,不仅能产生体液免疫,还能产生较强的细胞免疫,这对抵抗病毒及胞内寄生菌的侵染尤为重要。同时,核酸疫苗具有制备简单、生产周期短、成本低、易于构建成复合疫苗(多价苗或嵌合疫苗或联合苗)和便于储存运输等特点。2005 年,Castelruiz 等[14]将ADV的NS1 基因连接至真核表达载体Pvr1012,构建核酸疫苗免疫水貂,发现可以起到部分保护作用,这为阿留申病的疫苗研究提供了思路。
Bloom 等[12-15]将VP2 分成9 个非折叠的片段进行原核表达,证实VP2 上存在明确的与宿主致病性和宿主选择性密切相关的抗原决定簇,在探究衣壳蛋白柔性区的短肽序列是否是ADV 致病相关抗体的靶点过程中,发现 VP2 蛋白 428~446 aa 及 487~501 aa 为抗原抗体复合物主要的结合位置,介导ADE,将VP2中上述的2 个肽段去除将有望降低ADE。IL-12 及IFN-具有诱导机体产生细胞免疫的功能,被证明可作为佐剂增强疫苗的细胞免疫应答[16-17]。本研究通过比对国内外强毒株氨基酸序列,保留强毒株关键性位点,去除上述2 段导致ADE 的肽段后,进行核酸疫苗的构建,同时构建与细胞免疫相关细胞因子IL-12 及IFN-的真核表达质粒作为免疫佐剂,在体外,通过Western Blot及间接免疫荧光验证上述真核表达质粒的表达,旨在为后期用于免疫,研制出对水貂阿留申病有效的疫苗及未来应用于水貂阿留申病的防控提供借鉴和参考。
1 材料与方法
1.1 细胞株、质粒
293T细胞株及pVAX1 质粒为农业农村部经济动物疫病防控研究室保存。
1.2 主要试剂及仪器
T4 DNA连接酶(上海赛默飞世尔科技有限公司);Fugene HD 转染试剂(北京普洛麦格生物技术有限公司);FITC 标记的兔抗鼠IgG、Anti-6 His tags 抗体(上海艾博抗贸易有限公司);HRP 标记的山羊抗小鼠IgG、HRP 标记的山羊抗猫IgG、FITC 标记的羊抗猫IgG(美国KPL 生物公司);PrimeSTAR Max Polymerase、DH5感受态细胞(北京宝日医生物技术有限公司);ECL 显色液、蛋白分子量标准及核酸分子量标准(北京全式金生物有限公司);RIPA、蛋白酶抑制剂及小鼠IFN-ELISA 检测试剂盒(武汉博士德生物有限公司);小鼠IL-12 ELISA 检测试剂盒(上海江莱生物科技有限公司);聚丙烯酰胺凝胶配制试剂盒(上海碧云天生物技术有限公司);蛋白电泳仪(美国伯乐生物公司);PCR仪(美国应用生物系统公司);ADV阳性血清由农业农村部经济动物疫病重点实验室制备保存。
1.3 VP2主要抗原表位区及IL-12、IFN-序列确定和引物设计
以ADV-G 株VP2 基因序列为骨架,通过对实验室现有34 株国内ADV 强毒株的VP2 基因序列的比对,保留强毒株共有的关键性氨基酸位点,替换ADVG 株VP2 的相应表位位置,同时去除抗原抗体复合物形成的关键区域,将整合好的序列按照真核基因偏好性密码子进行优化后,交由南京金斯瑞生物科技有限公司进行基因合成。IL-12 的亚基(p35)序列参照Gen-Bank:NP_001152896.1,亚基(p40)序列参照Gen-Bank:NM_001303244.1亚基与 亚基之间的柔性肽为(G4S)4。IFN-序列参照 GenBank:XM_021205593.1,同时在氨基酸序列的C 端添加6 个组氨酸标签,交由南京金斯瑞生物科技有限公司进行基因合成,各基因分别连至克隆载体pMD18T 的多克隆位点中,命名为pMD18-M、pMD18-IL、pMD18-IFN-,为了与真核表达载体pVAX1 相连及后续表达,通过PCR 在引物的5'端增加kozak序列和酶切位点EcoR I,在3'端增加酶切位点XhoI及终止密码子,同时构建带有EGFP标签的pVAX1-EGFP的阳性质粒对照,VP2 主要抗原表位区(命名为M)的引物为M1、M2,IL-12 的引物为IL1、IL2,及 IFN-的引物为 IF1、IF2,EGFP 的引物为 EP1、EP2(表1),引物交由生工生物工程(上海)股份有限公司进行合成。

表1 引物序列Table 1 Primer sequences
1.4 VP2基因疫苗及IL-12、IFN-细胞因子佐剂真核表达质粒构建
用质粒提取试剂盒提取各克隆质粒,以此为模板,用设计的引物及高保真酶进行PCR,PCR 反应体系50L:PrimeSTAR Max Polymerase 25L,上下游引物各1.5L,模板 50 ng,ddH2O 补齐至 50L;PCR 扩增程序为 95 ℃预变性 10 min;95 ℃变性 45 s,55 ℃退火45 s,72 ℃延伸 1 min,35 个循环;72 ℃延伸 10 min。经1.0%琼脂糖凝胶电泳鉴定,胶回收后将产物连接于pVAX1 载体上,经酶切鉴定及测序后,确认克隆成功,将阳性质粒及pVAX1 空载体用EcoR I 及Xho I 进行双酶切,连接后,转化入DH5感受态细胞,接种于含卡那抗性的LB 琼脂平板,挑取阳性克隆接种于含卡那抗性LB 液体培养基,提取质粒进行双酶切鉴定及测序,将连接正确的VP2 阳性质粒命名为pVAX1-M,IL-12 阳性质粒命名为pVAX1-IL,IFN阳性质粒命名为pVAX1-IF,同时,连有EGFP 标签的阳性对照质粒命名为pVAX1-EGFP。
1.5 细胞转染
利用去内毒素质粒大提试剂盒提取重组质粒pVAX1-M、pVAX1-IL、pVAX1-IF、pVAX1-EGFP 及pVAX1 空载体,通过分光光度计检测浓度,保存于-20 ℃备用;取出培养至单层的293T 细胞,倒出原培养基,用PBS 清洗,加入适量0.25%的胰酶进行消化,镜下观察,当细胞有分散迹象时,加入DMEM培养基中和,轻柔吹打细胞,并进行计数。向6 孔细胞培养板中加入1 106个细胞,并用DMEM 培养基补齐至2 mL,置于37 ℃、CO2 培养箱进行培养,待细胞长至70%左右,按照Fugene HD 转染试剂说明书进行细胞转染,转染30 h,在倒置荧光显微镜下观察转染效果。
1.6 真核表达产物鉴定
1.6.1 间接免疫荧光 在细胞转染30 h 后进行间接免疫荧光试验。吸去6 孔细胞培养板中的DMEM 培养基,沿侧壁缓缓加入PBS清洗,将PBS吸净,加入预冷的4%的多聚甲醛1 mL,置于4 ℃冰箱固定30 min后,弃掉多聚甲醛,PBST 漂洗 3 次,每次 5 min;加入0.5%的TritonX-100 进行细胞打孔,于室温静置10 min,弃掉后,PBST 漂洗3 次,每次5 min;加入5%脱脂乳1 mL,37 ℃恒温培养箱封闭1 h;弃掉封闭液,PBST漂洗3 次,每次5 min;以ADV 阳性血清(稀释倍数1∶10)及His标签鼠单克隆抗体(稀释倍数1∶1 000)分别作为一抗,37 ℃孵育 1 h,PBST 漂洗 3 次,每次5 min,以FITC 标记的羊抗猫多克隆抗体(稀释倍数1∶5 000)及FITC 标记的兔抗鼠多克隆抗体(稀释倍数1∶5 000)分别作为二抗,37 ℃孵育 1 h,PBST漂洗3 次,每次5 min,在荧光显微镜下观察并进行拍照。
1.6.2 Western Blot 在细胞转染30 h后进行Western Blot。吸去6 孔细胞培养板中的DMEM 培养基,沿侧壁缓缓加入PBS清洗,将PBS吸净,每孔加入200L 的RIPA(含有终浓度为 1 mmol/L 的蛋白酶抑制剂PMSF),冰上放置10 min,待细胞脱落后,回收至1.5 mL离心管,4 ℃、12 000 r/min,离心 10 min,小心吸取上清,按比例加入6 protein loading buffer,煮沸5 min,每孔20L 样品,SDS-PAGE 电泳后,转印至 NC 膜,用含5%脱脂乳的TBST 缓冲液封闭,4℃过夜。pVAX1-IL、pVAX1-IF 的C 端带有6 His 标签,分别以小鼠抗His标签抗体(稀释倍数1∶1 000)和ADV 阳性血清(稀释倍数1∶200)为一抗,山羊抗小鼠HRP-IgG(稀释倍数1∶10 000)和山羊抗猫HRP-IgG(稀释倍数1∶4 000)为二抗,进行Western Blot 分析目的蛋白的表达及反应原性。
1.6.3 ELISA (1)IL-12 的检测:细胞转染 48 h 后,利用细胞刮将细胞刮下,4 ℃、2 000 r/min,离心20 min,取上清10L 按照1∶10 进行稀释,即15L 的细胞上清+135L 的样品稀释液,每孔50L 进行 ELISA检测。取出试剂盒中的酶标板恢复至室温,标准品孔各加不同浓度(80、40、20、10、5、2.5pg/mL)的标准品 50L,除空白孔外,标准品孔和样品孔中每孔加入辣根过氧化物酶(HRP)标记的检测抗体100L,用封板膜封住反应孔,37 ℃恒温培养箱孵育60 min。弃去液体,吸水纸拍干,每孔加满洗涤液350L,静置 1 min,甩去洗涤液,吸水纸拍干,如此重复5 次。每孔加入底物A、B各50L,37℃避光孵育15min。每孔加入终止液50L,15 min 内,在 450 nm 波长处测定各孔的OD 值。利用ELISA Calc回归计算程序-v0.1 绘制标准曲线,并计算样品中IL-12 的浓度。(2)IFN-的检测:细胞转染48h后,利用细胞刮将细胞刮下,4 ℃、2 000 r/min,离心20 min,取上清25L 按照1∶2 进行稀释,即120L 的细胞上清+120L的样品稀释液。IFN-标准品稀释按照博士德小鼠IFN-检测试剂盒说明书进行,分别为2 000、1 000、500、250、125、62.5、31.2 pg/mL。将各梯度的标准品及样品各100L 依次加入孔中,每个做1 个复孔,酶标板加上封板膜,37 ℃孵育90 min;甩去液体,吸水纸拍几下,不洗;将准备好的生物素抗小鼠IFN-抗体工作液每孔100L依次加入(TMB显色孔除外)。酶标板加上封板膜,37 ℃孵育60 min;洗板3 次,每次浸泡1 min;将准备好的ABC 工作液按每孔100L依次加入(TMB 空白显色孔除外),酶标板加上封板膜,37 ℃孵育 30 min;洗板5 次,每次浸泡1 min;每孔加入TMB 显色液90L,37 ℃避光孵育 20 min;按每孔100L 加入TMB 终止液,此时蓝色立即变为黄色。用酶标仪在450 nm 波长处测定OD 值。利用软件CurveExpert 1.4,输入数据,绘制标准曲线,得到回归方程,计算样品中IFN-的浓度。
1.6.4 统计学分析 ELISA检测获得的结果采用平均数±标准差的形式表示。采用SPSS 20.0 统计分析软件和单因素方差分析方法进行统计分析。
2 结果与分析
2.1 真核表达质粒酶切鉴定结果
通过PCR 和双酶切方法,将VP2 抗原表位M、小鼠细胞因子IL-12 片段、小鼠细胞因子IFN-片段及荧光标签EGFP 基因分别连接于真核表达载体pVAX1,获得目的质粒 pVAX1-M、pVAX1-IL、pVAX1-IF 及pVAX1-EGFP,经 EcoR I 及 Xho I 双酶切鉴定,琼脂糖凝胶电泳均显示有2 条特异条带,大小与预期一致(图1~4),目的基因的条带大小分别为 1 866、1 748、518、720 bp;进一步测序结果表明,序列正确,各真核表达质粒构建成功。

图1 pVAX1-M 双酶切鉴定结果Fig.1 pVAX1-M double digestion identification result

图2 pVAX1-IL 双酶切鉴定结果Fig.2 pVAX1-IL double digestion identification result

图3 pVAX1-IF 双酶切鉴定结果Fig.3 pVAX1-IF double digestion identification result
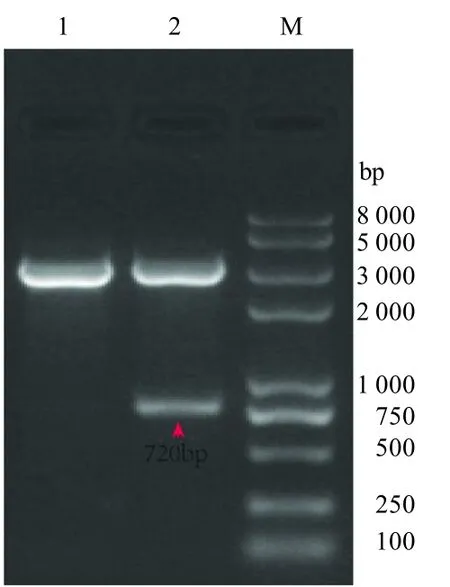
图4 pVAX1-EGFP 双酶切鉴定结果Fig.4 pVAX1-EGFP double digestion identification result
2.2 细胞转染结果
为观察转染效果,将pVAX1-EGFP同步转染入293T细胞中,30 h 后观察转染效果(图5),结果显示,转染成功,镜下可见明显的荧光,证明转染体系合理,且真核表达载体pVAX1 可正常表达蛋白。
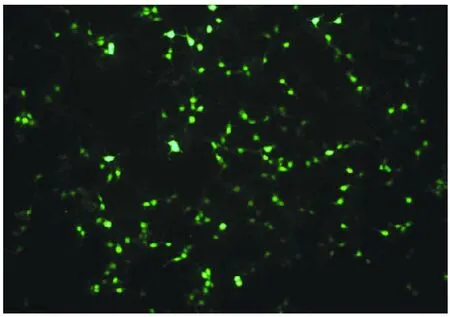
图5 pVAX1-EGFP 转染结果Fig.5 Transfection result of pVAX1-EGFP
2.3 间接免疫荧光鉴定
2.3.1 重组蛋白反应原性的鉴定 将 pVAX1-M 及pVAX1 转染至293T 细胞,培养30 h 后,利用间接免疫荧光,对其表达情况进行验证,一抗为ADV 阳性血清,结果显示,pVAX1-M质粒孔可见特异性荧光,空载体pVAX1 无(图6),证明pVAX1-M 质粒在体外可正常表达蛋白,且该蛋白具有反应原性。

图6 pVAX1-M质粒及空载体质粒pVAX1 体外转染间接免疫荧光结果Fig.6 Indirect immunofluorescence results of transfection of pVAX1-M plasmid and empty vector plasmid pVAX1
2.3.2 细胞因子表达情况 将各细胞因子真核表达质粒转染至293T 细胞,培养30 h 后,利用间接免疫荧光,对其表达情况进行验证,一抗为His 标签鼠单抗,结果显示,pVAX1-IL 质粒孔可见特异性荧光(图7),pVAX1-IF质粒孔可见特异性荧光(图8),而对照空载体pVAX1 质粒孔无特异性荧光(图9),证明各细胞因子真核表达质粒可在体外正常表达蛋白。



2.4 Western Blot鉴定
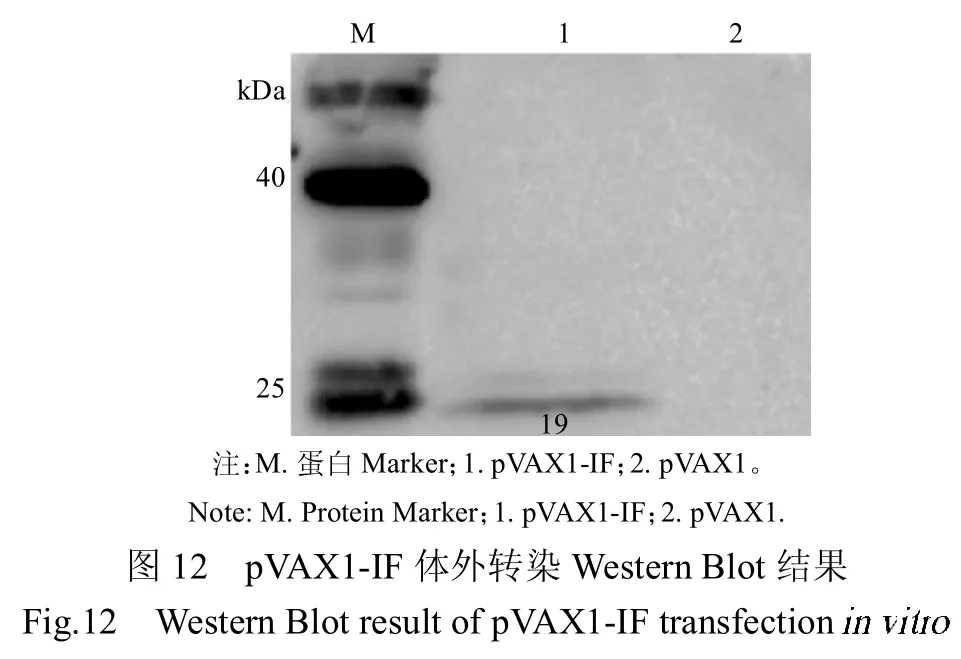
将含目的基因的质粒转染至293T细胞,培养30 h后,将细胞刮下,提取蛋白,加入蛋白protein loading buffer 进行制样,利用Western Blot 对其表达情况进一步验证,一抗分别为ADV 阳性血清及His 单抗,结果显示,各真核表达质粒可正常表达蛋白,pVAX1-M 表达的蛋白大小约为70.5 kDa(图10),且具有反应原性,pVAX1-IL 表达蛋白大小约为 60.0 kDa(图11),pVAX1-IF 表达的蛋白大小约为19.0 kDa(图12)。


2.5 ELISA鉴定
细胞转染48 h 后,将细胞刮下,离心后取上清进行ELISA 检测,利用各细胞因子标准品的OD 值及对应浓度得到的回归方程绘制出标准曲线,其中,IL-12的标准曲线回归方程为 y=0.106+0.027x(r2=0.997);IFN-的标准曲线回归方程为 y=(11.823+179.623x)/(1 33.912x 318.513x2)(r2=0.999)。标准曲线见图13、14。
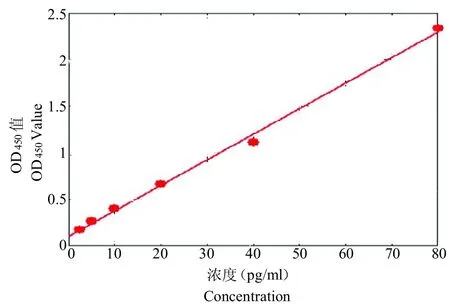
图13 IL-12 标准曲线Fig.13 IL-12 standard curve

图14 IFN-标准曲线Fig.14 IFN-standard curve
根据检测样品的OD 值换算其浓度,结果显示,pVAX1-IL 及 pVAX1-IF 中 IL-12 或 IFN的水平明显高于pVAX1 空载对照组,差异极显著(P <0.01),结果见表2 和图15,证明构建的真核表达质粒pVAX-IL及pVAX1-IF在体外可有效表达目的蛋白,且具有活性,后期可作为免疫佐剂使用。

表2 IL-12 及IFN- 表达情况Table 2 IL-12 and IFN- expression (pg/mL)

图15 IFN与IL-12 浓度柱形图Fig.15 Histogram of IFN-and IL-12 concentration
3 讨论
AD 是典型的免疫复合物疾病,动物在感染AD后,机体会产生高水平的抗体,却不能中和病毒,反而会与抗原形成复合物,沉积于肾脏上皮或动脉并发炎症,抗体进一步介导ADV 的侵染,产生抗体依赖性增强作用(ADE)。作为水貂养殖业的三大疫病之一,研发出有效的疫苗进行防控为首选手段,前人也曾尝试研制常规的疫苗,如灭活疫苗、减毒疫苗及亚单位疫苗等,但由于该病特殊的致病特点,均未有良好的保护效果,甚至因抗体依赖性增强导致病毒的进一步侵染,ADV 的防控亟需研制一种安全有效的疫苗。
近年,新型疫苗的涌现给疾病防控带来疫苗研发新角度,其中作为引发第三次疫苗革命的核酸疫苗被研究者青睐。由于核酸疫苗在进入体内后,是由机体细胞来表达外源蛋白,进而引发机体免疫反应,这种内源性的抗原可突破传统疫苗只能产生较强的体液免疫的限制,可产生较强的细胞免疫反应。而AD由于其ADE作用,传统疫苗产生的抗体并非保护性抗体,反而加重疾病,立足于体液免疫的传统疫苗显然不适合阿留申病疫苗的研发,细胞免疫对其免疫效果尤为重要。为增强疫苗的免疫效果,佐剂常被应用于疫苗免疫中,而佐剂的选择取决于疫苗的类型和免疫目的,有研究者为增强疫苗细胞免疫的效果,利用与细胞免疫相关的细胞因子IL-12 及IFN的联合免疫增强机体的细胞免疫应答[18-19]。IL-12 由异源二聚体p30 和p45 通过共价键连接而成,小鼠IL-12 的分子量大小约为60 kDa,具有重要的免疫调节功能,主要诱导机体产生细胞免疫应答,促进Th0 细胞增殖分化成Th1 细胞,诱导其产生IFN-,从而增强细胞免疫,2 个亚基共同表达才可发挥其生物学功能[20-22]。IFN-即II型干扰素,小鼠IFN-分子量大小约为19 kDa,是一种具有抗病毒、抗肿瘤并且具有免疫调节作用的细胞因子,可促进抗原递呈细胞表达组织相容性复合物,从而增强抗原递呈能力,同时可诱导Th0 细胞分化为Th1 细胞,从而增强细胞免疫水平[23-24]。登革热病是一种导致人死亡率及发病率很高的烈性传染病,在其疫苗的研发中同样受ADE 效应困扰,国外有学者通过构建登革热病毒(DENV)E 蛋白的DIII 结构域的核酸疫苗,可引起该病毒 4 种血清型的免疫反应[25]。2015 年,Tang等[26]在定位DENV2 型的E 蛋白I~II 结构域中导致抗体依赖性增强的表位后,构建去除导致ADE的表位基因片段的DNA 疫苗,发现与野生型相比,该DNA 疫苗在体内可产生良好的中和保护效应。由此可见,构建去除ADE 片段的核酸疫苗对ADE 相关疾病的疫苗研发具有参考价值,这也为 ADV 疫苗的研发提供了思路。VP2 作为ADV 衣壳蛋白的主要构成蛋白,成为疫苗研发的首选抗原,虽然表面有丰富的抗原决定簇,但由于ADE 作用,前人曾将VP2 免疫动物,并不能起到期望的保护力。Bloom 等[15]研究发现抗原抗体复合物结合和产生 ADE 作用的主要位置为 VP2 蛋白的428~446 aa 及 487~501 aa。在 ADV 核酸疫苗的构建中,选择含有丰富抗原决定簇的VP2 的基因片段为骨架,去除上述导致抗原抗体复合物结合和产生ADE作用的2 段肽段,将降低VP2 的ADE 作用,同时有望在免疫后产生中和保护效应,这种新型的AD 核酸疫苗为解决目前AD 无疫苗防控带来了希望。
本研究以DNA 疫苗专用载体pVAX1 为载体,构建了不含抗原抗体复合物表位的 VP2 基因疫苗及IL-12 和IFN细胞因子佐剂,通过转染293T细胞,分别进行间接免疫荧光、Western Blot 和ELISA 验证目的基因在体外表达情况,结果表明,表达的蛋白具有良好的反应原性,为下一步应用奠定了基础。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
交通医学(2022年2期)2022-06-14
昆明医科大学学报(2021年5期)2021-07-22
江西农业学报(2021年4期)2021-04-20
科学导报(2021年3期)2021-02-22
昆明医科大学学报(2021年1期)2021-02-07
三农资讯半月报(2020年11期)2020-06-21
江苏农业学报(2019年1期)2019-09-10
生物学教学(2018年10期)2018-11-29
