
黄波椤生长性状种内遗传变异
2020-04-16 05:53:54唐晓杰潘树百王德秋王连福程广有
北华大学学报(自然科学版) 2020年2期
唐晓杰,潘树百,王德秋,张 悦,王连福,程广有
(1.北华大学林学院,吉林 吉林 132013;2.吉林省临江林业局,吉林 临江 134600)
黄波椤(PhellodendronamurenseRupr.)为芸香科黄檗属落叶乔木[1-2],是东北亚特有种,阳性树种,耐严寒,不耐荫,在林冠下自我更新不良,但能在空旷地完成更新.黄波椤是古老的孑遗植物,被列为国家二级重点保护树种[3-6].黄波椤是我国东北阔叶红松林的重要伴生树种,是重要的国防和工业用材,为“东北三大硬阔”之一[7-9],韧皮部即关黄柏,经炮制后入药,是临床应用广泛的药物[10-11].黄波椤广泛应用于园林景观中[12],可做杀虫剂[13],可制成美容类产品、天然防腐剂[14].
黄波椤以种子繁殖为主.种子有休眠,春季播种前需要层积处理50~60 d,也可在11月进行秋播.黄波椤韧皮部含有多种生物碱,植株中的生物碱含量因树龄、季节、栽培措施不同而异[15-20],不同群体、个体间活性成分含量也不同[21-23].随着对黄波椤生物碱药理作用研究的深入,以黄波椤为原料生产的生物药品种越来越多.由于过度采伐,黄波椤资源处于衰竭状态,木材和关黄柏供不应求.遗传变异是良种选育的理论基础,到目前为止,有关黄波椤生长性状变异规律的报道甚少.本文主要研究无性繁殖条件下黄波椤的生长性状变异规律,以期为黄波椤优良品种选育提供依据.
1 材料与方法
1.1 试验材料
黄波椤无性系对比林建于1999年,位于临江林业局金山林场,N 126°54′,E 41°48′,海拔793 m,年均气温2~4 ℃,全年降水量750~1 000 mm,属中温带大陆性季风气候.2017年7月,在黄波椤无性系测定林中选择黑龙江种源1、临江种源1、临江种源2、临江种源3,调查树高、胸径、冠幅、树皮厚度和枝条连年生长量.其中,黑龙江种源1雌无性系20个、雄无性系10个;临江种源1雌无性系22个、雄无性系15个;临江种源2雌无性系18个、雄无性系10个;临江种源3雌无性系24个、雄无性系5个.上述雌无性系各25个分株,雄无性系各10个分株.文中用字母代表种源和无性系,比如,H1M8代表黑龙江种源1第8号雄无性系,L2F9代表临江种源2第9号雌无性系.
1.2 试验方法
测量并记录各种源无性系树高、胸径、冠幅、树皮厚度以及枝条连年生长量.
1.3 数据统计方法
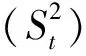
利用离差平方和法对不同种源雌、雄无性系生长性状进行系统聚类分析.按照欧氏距离D2将各无性系划分为不同组别.
假设所有对象的X的均值为m,标准差为s,则标准化后的值=(标准化前的值-各属性的均值)/各属性的标准差,所以加权的欧式距离为
式中:Distance(Ui,Uj)为标准化后的(距离)值;Uik为某个性状(k)第i个标准化前的值;Sk为该性状标准差;n为该性状观测个体数量.
2 结果与分析
2.1 黄波椤生长性状
黄波椤无性系树高、地径、冠幅、树皮厚度和连年生长量统计结果见表1.由表1可知:种源内,树高和冠幅平均值雌、雄株间差异不大,变异系数较小,说明树高和冠幅种源内单株间变异较小.黑龙江种源内地径和树皮厚度平均值雌、雄株间差异较大,变异系数亦较高,表明种源内个体间径向生长变异大.树高和冠幅的标准差较小,而连年生长量和地径标准差较大.标准差是个体偏离平均值距离的体现,说明黄波椤树高和冠幅的个体观测值与平均值接近,连年生长量和地径的观测值偏离平均值较远.黄波椤无性系对比林是采用随心形成层对接方法繁殖的嫁接苗,砧木和嫁接技术不同会影响嫁接植株生长发育.
2.2 黄波椤种源间生长性状变异
黄波椤种源间各生长性状方差分析结果见表2.由表2可知:黄波椤树高、地径、冠幅和连年生长量种源间差异均达到极显著水平,雄无性系树皮厚度种源间差异极显著,雌无性系树皮厚度种源间差异不显著,说明黄波椤生长性状种源间变异较大.
进一步对种源间生长性状进行多重比较,结果见图1~3.结果表明:同一种源黄波椤树高、地径和冠幅雌、雄株间差异不显著;不同种源间差异显著性方面,黑龙江种源株高、地径和冠幅均显著高于临江种源,临江种源1显著高于临江种源2和3,临江种源2和3间差异不显著.
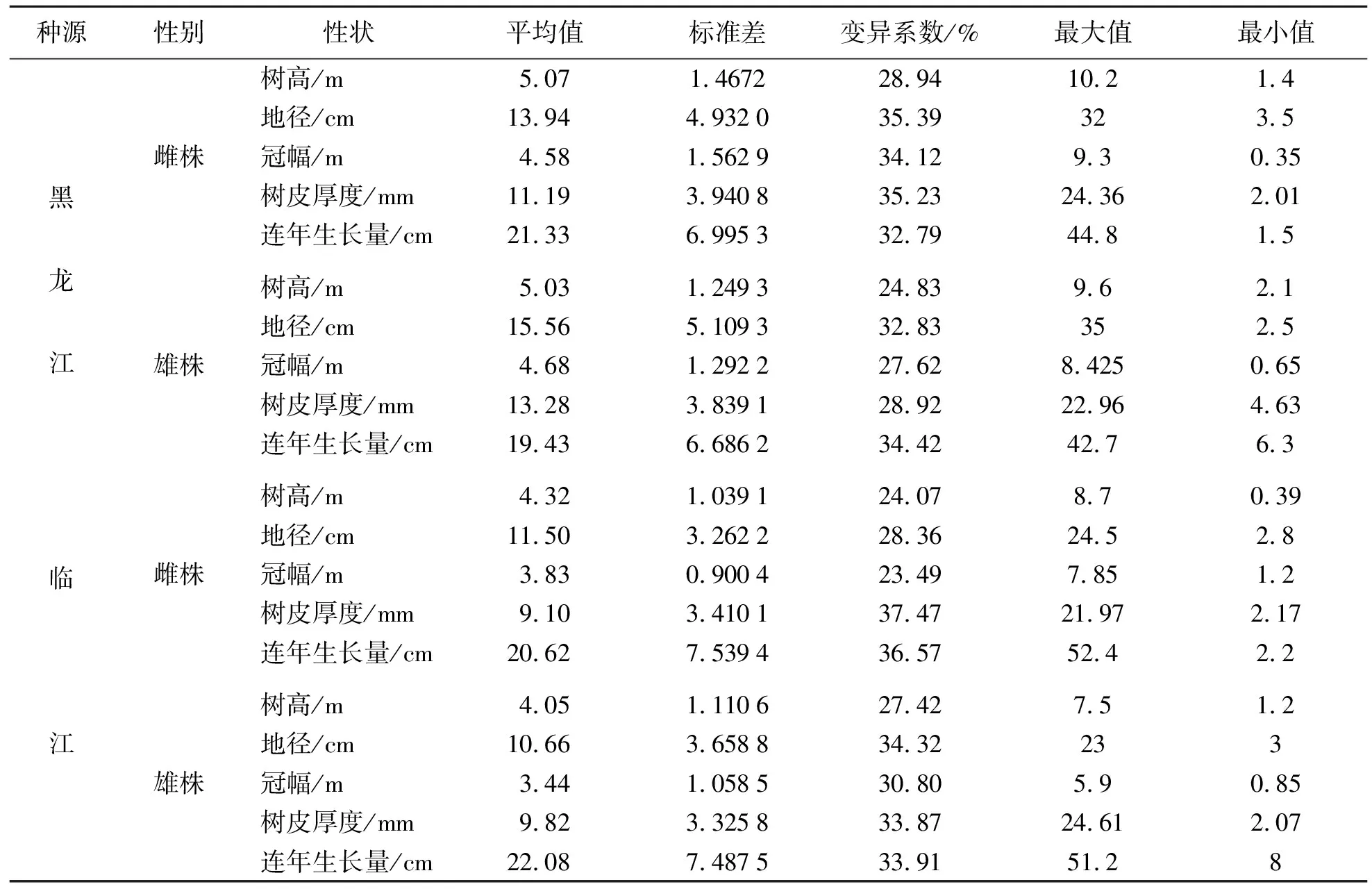
表1 黄波椤无性系生长性状统计Tab.1 Statistical results of growth traits of asexual lines of Phellodendron amurense Rupr.
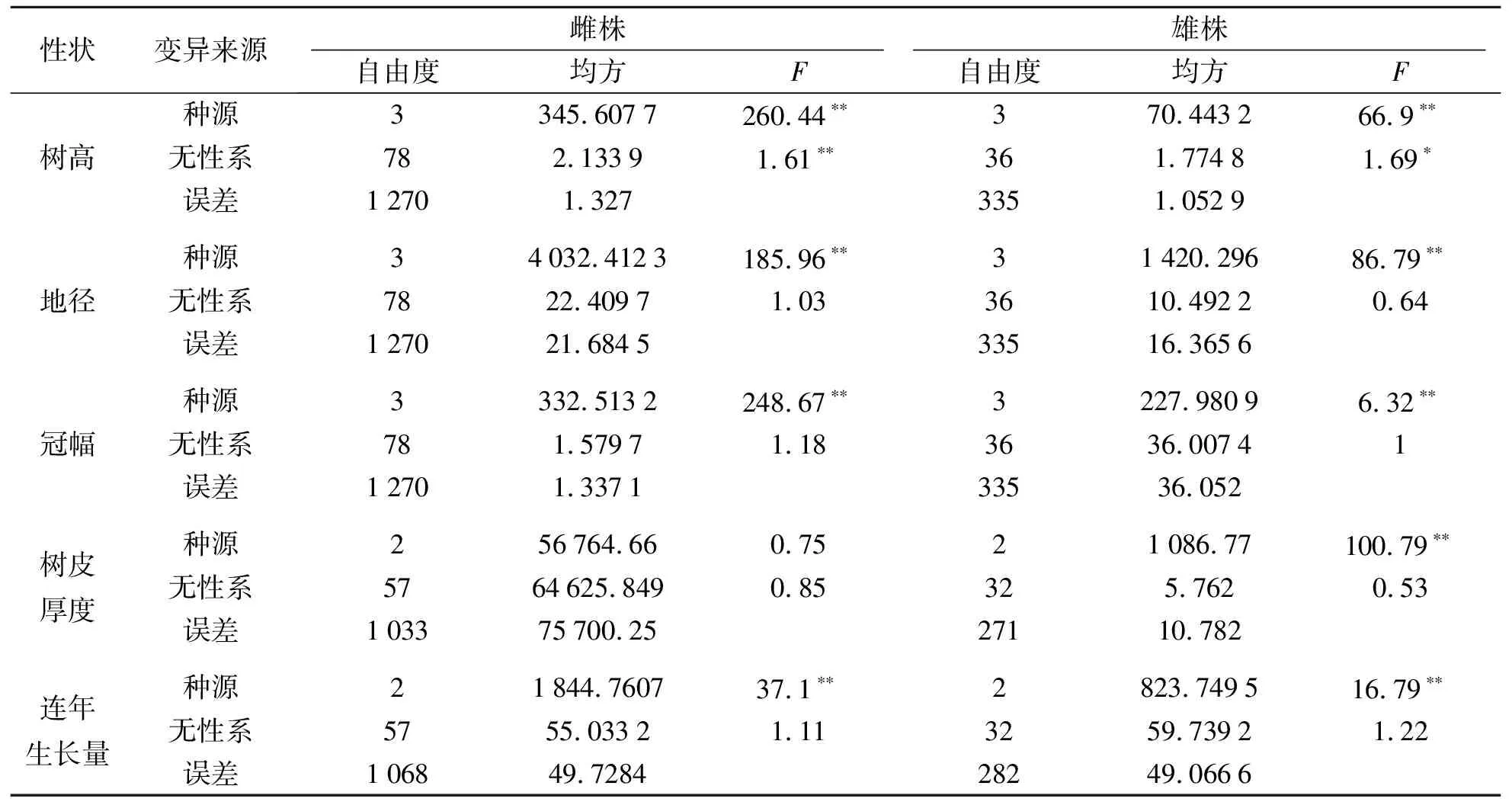
表2 黄波椤种源间各生长性状方差分析Tab.2 Variance analysis of growth traits among provenances of Phellodendron amurense Rupr.
注:**.差异极显著;*.差异显著.下同.
2.3 黄波椤无性系间生长性状变异
黑龙江种源内无性系间生长性状方差分析结果见表3.由表3可知:雌性树高无性系间差异显著,雄性树高无性系间差异极显著,无性系间地径、冠幅和树皮厚度差异不显著.
临江种源1无性系间生长性状方差分析结果见表4.由表4可知:临江种源1雌性树高和冠幅无性系间差异极显著,雌无性系间地径和树皮厚度差异不显著,树高、地径、冠幅和树皮厚度雄无性系间差异均不显著.
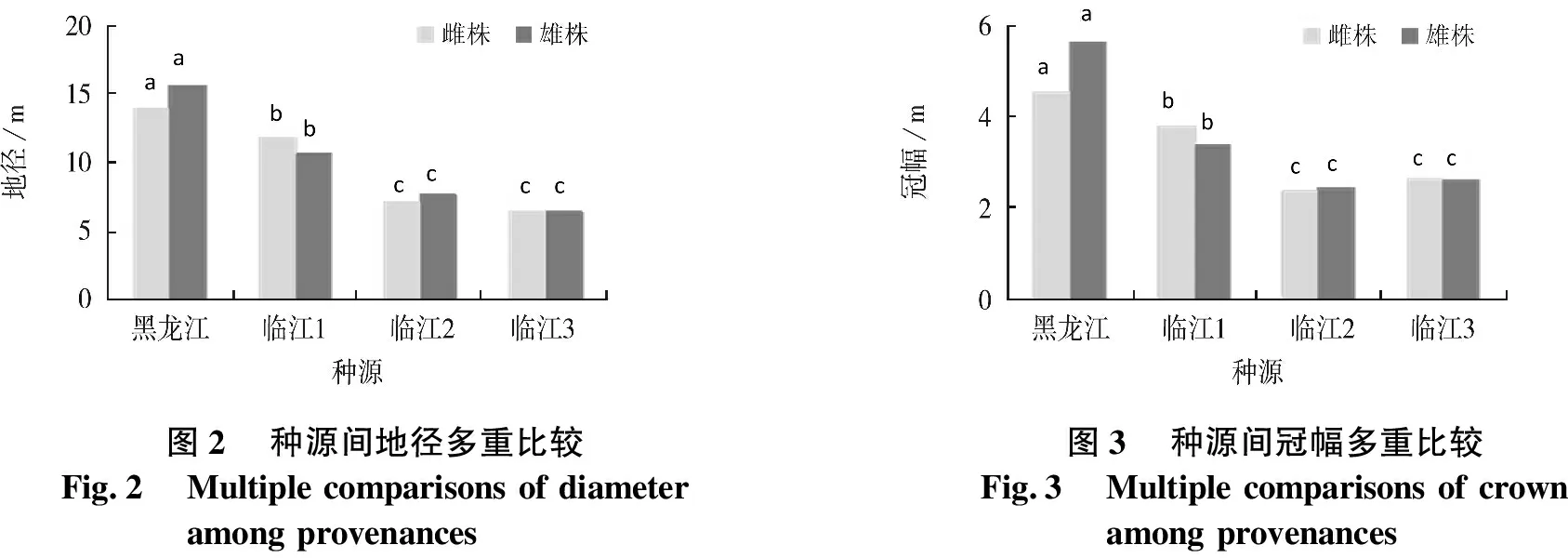
图2种源间地径多重比较Fig.2Multiple comparisons of diameteramong provenances 图3种源间冠幅多重比较Fig.3Multiple comparisons of crownamong provenances
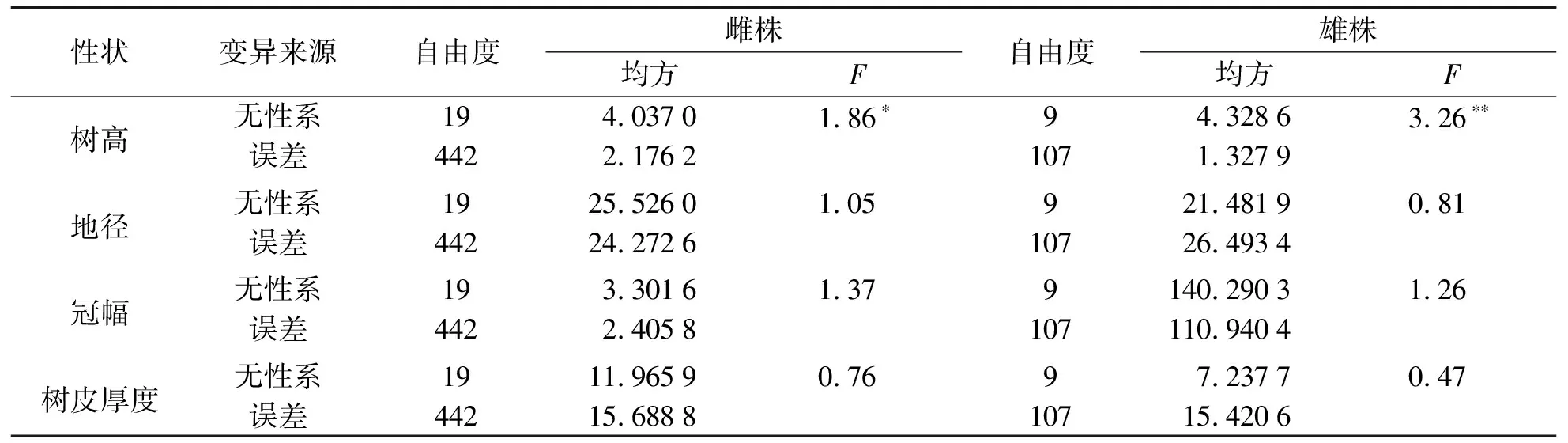
表3 黑龙江种源无性系间各生长性状方差分析Tab.3 Variance analysis of growth traits among the asexual lines of Heilongjiang provenance
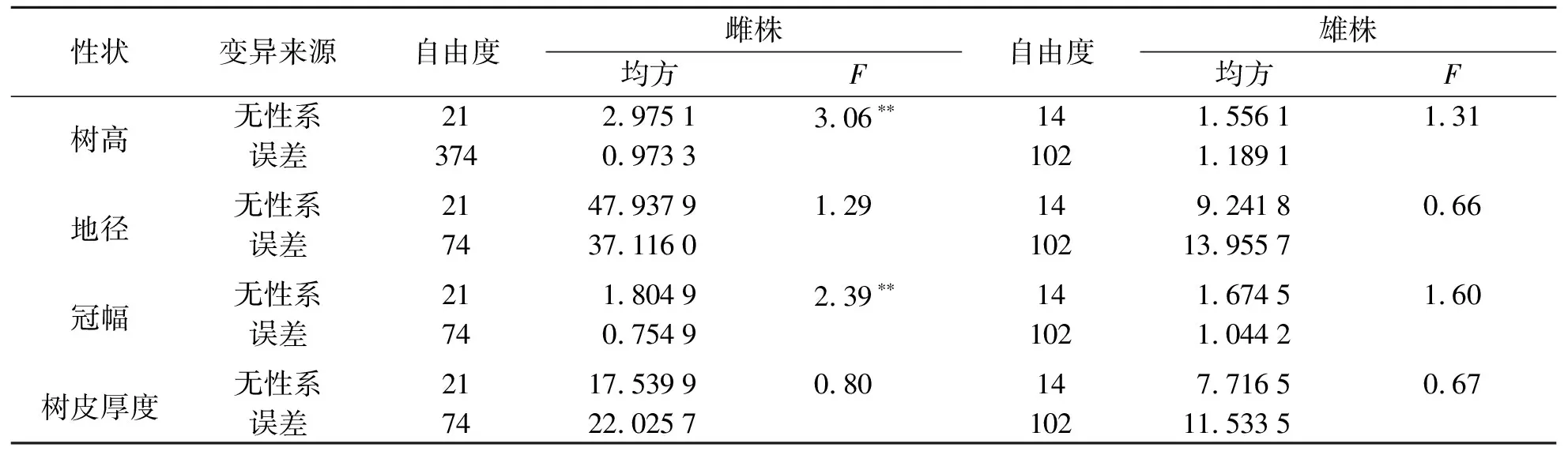
表4 临江种源1无性系间生长性状方差分析Tab.4 Variance analysis of growth traits among the asexual lines of Linjiang Ⅰ provenance
黑龙江种源无性系间聚类分析结果见图4、图5.由图4、图5可知,当欧氏距离D2=7.5时,将10个雄无性系聚为3类:H1M8、H1M9和H1M2聚为一类;H1M1、H1M6和H1M5聚为一类;其余4个无性系聚为一类.其中,H1M3自成1个亚类;H1M4、H1M7和H1M10被聚为第2个亚类.当D2=8.5时,将20个雌无性系聚为4类:H1F14、H1F15、H1F13、H1F5、H1F7和H1F16聚为一类;H1F1、H1F4、H1F9和H1F12聚为一类;H1F17自成一类;其余9个无性系聚为一类.进一步分类可知:H1F18和H1F20被聚为第1个亚类;H1F3、H1F19、H1F2、H1F8、H1F11、H1F10和H1F6聚为第2个亚类.
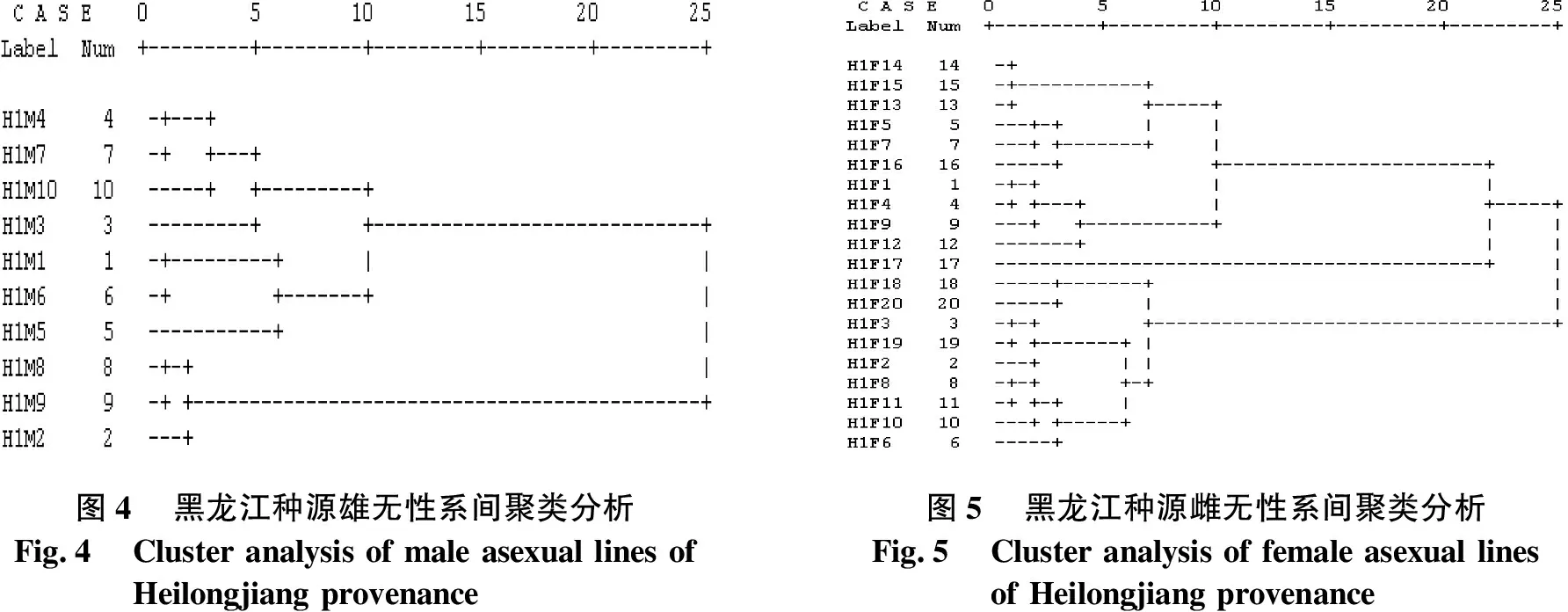
图4黑龙江种源雄无性系间聚类分析Fig.4Cluster analysis of male asexual lines ofHeilongjiang provenance图5黑龙江种源雌无性系间聚类分析Fig.5Cluster analysis of female asexual linesof Heilongjiang provenance
临江种源1无性系间聚类分析结果见图6、图7.由图6、图7可知,当D2=10时,将15个雄无性系聚为3类:L1M2、L1M9、L1M4和L1M13被聚为一类;L1M1、L1M15、L1M10和L1M11被聚为一类;其余7个无性系聚为一类.其中,L1M7和L1M14被聚为1个亚类;L1M5、L1M12、L1M8、L1M3和L1M6被聚为第2个亚类.当D2=9.5时,将22个雌无性系聚为3类:L1F19和L1F20被聚为一类;L1F17、L1F21和L1F14被聚为一类;其余17个无性系被聚为一类.其中,L1F4、L1F16、L1F15、L1F18、L1F9、L1F6、L1F12和L1F11被聚为1个亚类;L1F5、L1F7、L1F13、L1F3、L1F22、L1F1、L1F10、L1F2和L1F8被聚为第2个亚类.
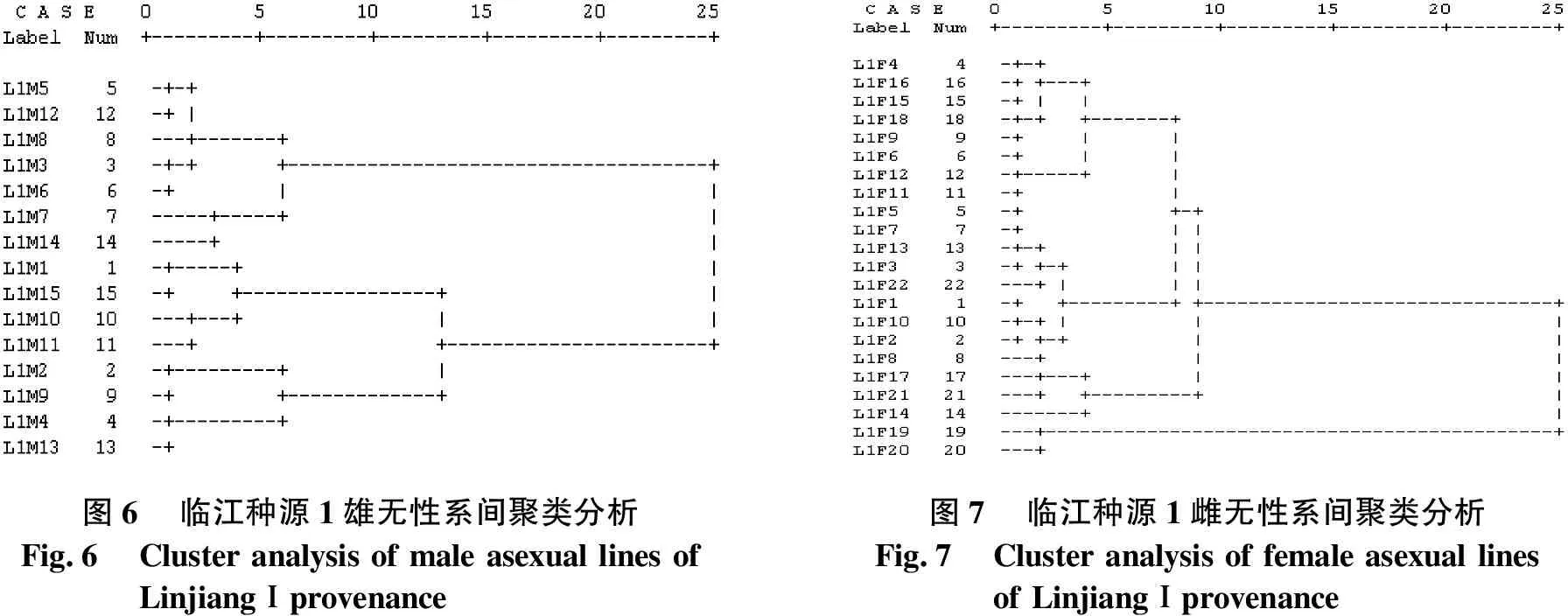
图6临江种源1雄无性系间聚类分析Fig.6Cluster analysis of male asexual lines ofLinjiangⅠprovenance图7临江种源1雌无性系间聚类分析Fig.7Cluster analysis of female asexual linesof LinjiangⅠprovenance
临江种源2无性系间聚类分析结果见图8、图9.由图8、图9可知,当D2=15时,将10个雄无性系聚为3类:L2M1、L2M4和L2M7被聚为一类;L2M3、L2M10和L2M5被聚为一类;L2M2、L2M8、L2M9和L2M6被聚为一类.当D2=10时,将18个雌无性系聚为4类:L2F14自成一类,L2F3、L2F13和L2F6被聚为一类;L2F1、L2F10、L2F2、L2F15、L2F18和L2F4被聚为一类;其余8个无性系聚为一类.其中,L2F7自成1个亚类;L2F9、L2F12和L2F5被聚为第2个亚类;L2F16、L2F17、L2F8和L2F11被聚为第3个亚类.
临江种源3无性系间聚类分析结果见图10、图11.由图10、图11可知,当D2=12.5时,将5个雄无性系聚为2类:L3M2和L3M4被聚为一类;其余3个无性系聚为一类.其中,L3M5自成1个亚类;L3M1和L3M3被聚为第2个亚类.当D2=15时,将24个雌无性系聚为2类:L3F14、L3F19、L3F15、L3F20和L3F12被聚为第一类的第1个亚类,L3F5、L3F8、L3F21和L3F24被聚为第2个亚类,L3F13和L3F23聚为第3个亚类;L3F10自成第2类的第1个亚类,L3F2、L3F11、L3F22、L3F1和L3F18聚为第2个亚类,L3F6、L3F9、L3F16、L3F7、L3F3、L3F4和L3F17聚为第3个亚类.
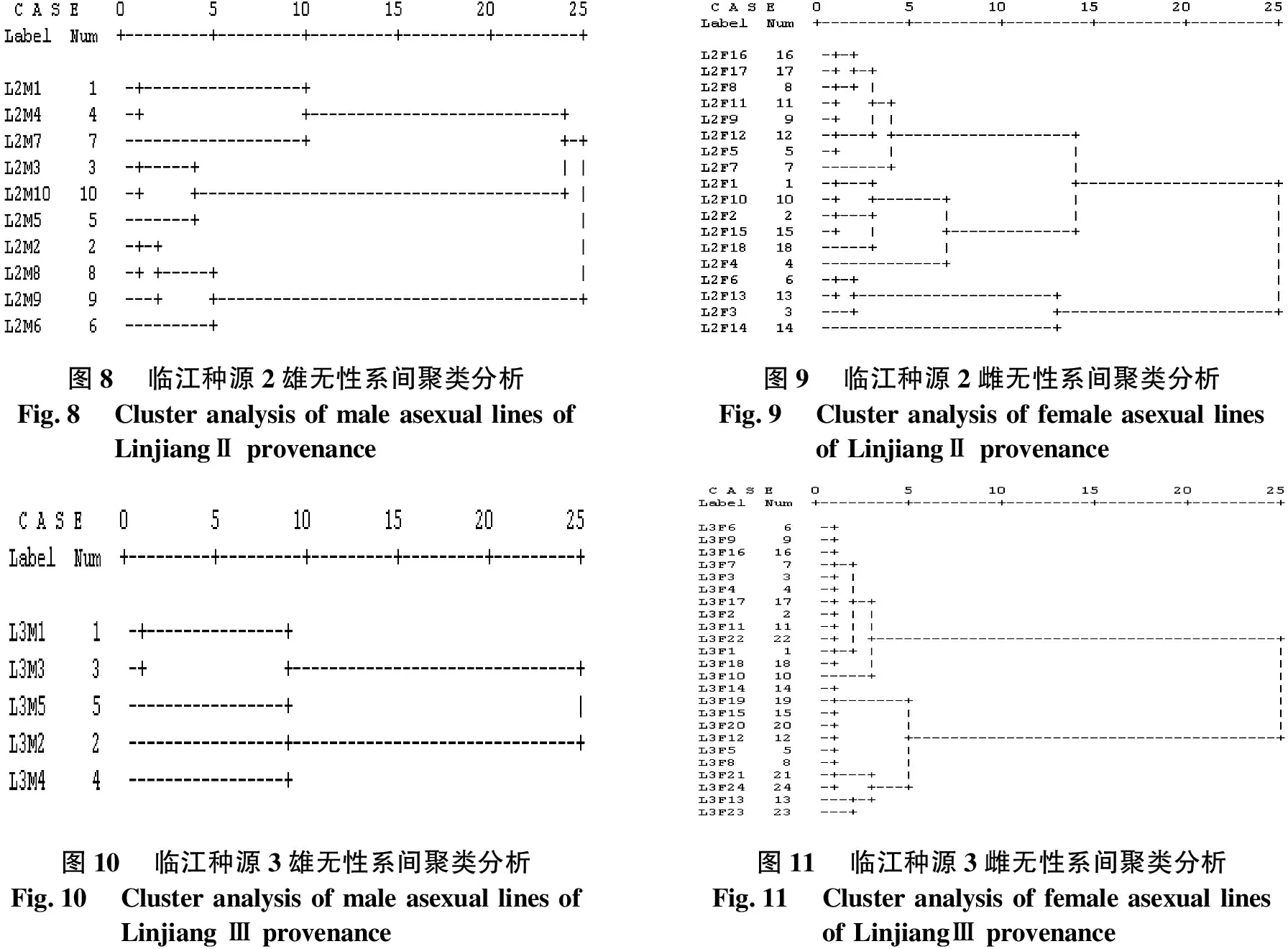
图8临江种源2雄无性系间聚类分析Fig.8Cluster analysis of male asexual lines ofLinjiangⅡ provenance图9临江种源2雌无性系间聚类分析Fig.9Cluster analysis of female asexual linesof LinjiangⅡ provenance图10临江种源3雄无性系间聚类分析Fig.10Cluster analysis of male asexual lines ofLinjiang Ⅲ provenance图11临江种源3雌无性系间聚类分析Fig.11Cluster analysis of female asexual linesof LinjiangⅢ provenance
2.4 黄波椤生长性状特征方差分量分析
计算无性系对比林内雌、雄株生长性状特征的方差分量比,用方差分量占遗传总变异的比例表示种源间的表型分化系数,结果见表5.由表5可知:树高、地径、冠幅、树皮厚度和连年生长量表型分化系数的变异幅度为45.74%~99.68%.雌株树高、地径、冠幅、树皮厚度和连年生长量的表型分化系数均值分别为93.36%、98.30%、97.21%、97.16%和94.61%,雄株树高、地径、冠幅、树皮厚度和连年生长量的表型分化系数均值分别为87.29%、45.74%、99.68%、94.67%和97.22%.由各生长性状的表型性状平均值可以发现,种源间平均表型方差分量占总变异的37.09%,种源内无性系间平均表型方差分量占总变异的1.64%,机误的方差分量占总变异的61.27%.树高、地径、冠幅和树皮厚度种源间方差分量比例均大于30%,且均大于林分内无性系间的方差分量比例.进一步分析发现,黄波椤无性系测定林各生长性状种源间变异对总变异的贡献为95.77%,种源内无性系间的贡献为4.23%,说明黄波椤群体间的遗传变异大于个体间.
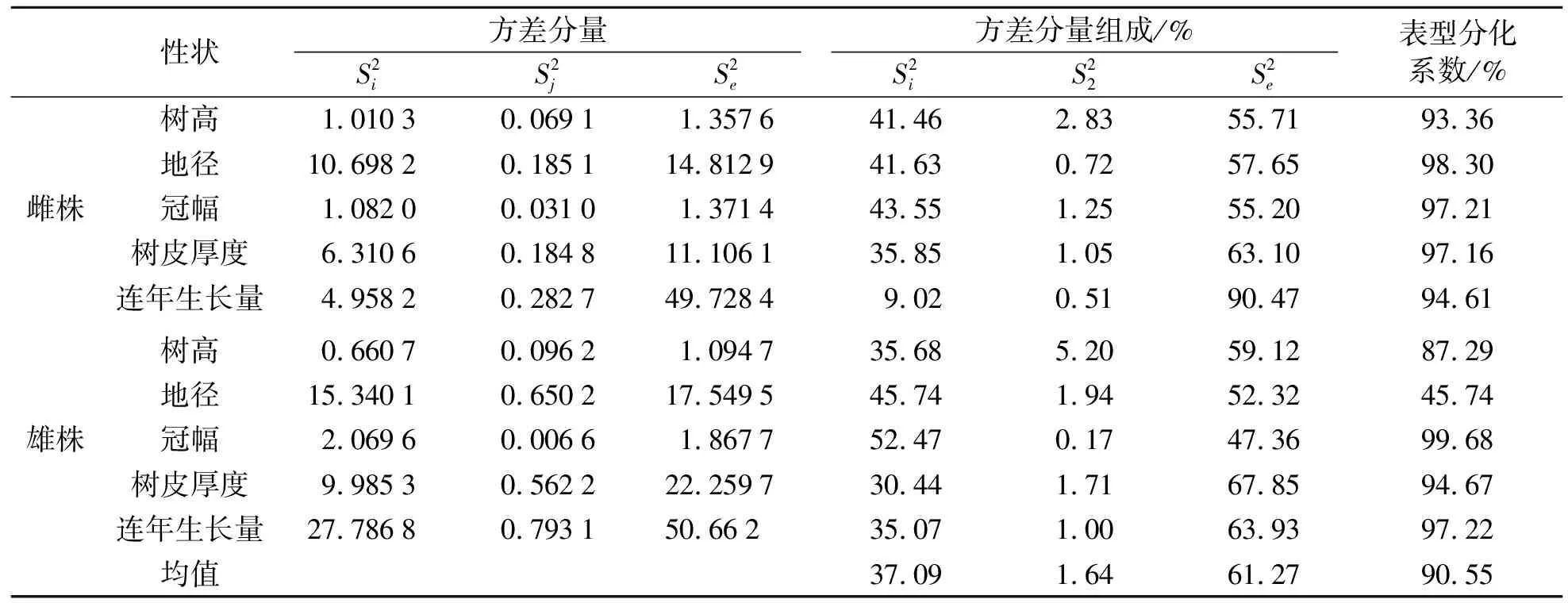
表5 黄波椤生长性状遗传参数Tab.5 Genetic parameters of growth traits of Phellodendron amurense Rupr.
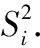
3 小 结
1)生长性状种源间变异.同一黄波椤种源树高、地径和冠幅雌、雄株间差异较小.树高、地径和冠幅种源间差异均达到极显著水平,雄性树皮厚度种源间差异极显著,雄性连年生长量种源间差异极显著.黑龙江种源株高、地径和冠幅均显著高于其他种源.
2)生长性状无性系间变异.黑龙江种源内黄波椤树高雌性无性系间差异显著,雄性无性系间差异极显著,无性系间地径和冠幅差异不显著.临江种源1内树高和冠幅雌性无性系间差异极显著.黑龙江种源内,当D2=7.5时,将10个雄无性系聚为3类;当D2=8.5时,将20个雌无性系聚为4类.临江种源1内,当D2=10时,将15个雄无性系聚为3类;当D2=9.5时,将22个雌无性系聚为3类.临江种源2内,当D2=15时,将10个雄无性系聚为3类;当D2=10时,将18个雌无性系聚为4类.临江种源3内,当D2=12.5时,将5个雄无性系聚为2类;当D2=15时,将24个雌无性系聚为2类.
3)生长性状变异来源.树高、地径、冠幅、树皮厚度和连年生长量表型分化系数的变异幅度为45.74%~99.68%,雌株树高、地径、冠幅、树皮厚度和连年生长量的表型分化系数均值分别为93.36%、98.30%、97.21%、97.16%和94.61%,雄株树高、地径、冠幅、树皮厚度和连年生长量的表型分化系数均值分别为87.29%、45.74%、99.68%、94.67%和97.22%.种源间平均表型方差分量占总变异的37.09%,种源内无性系间平均表型方差分量占总变异的1.64%,机误的方差分量占总变异的61.27%;黄波椤无性系测定林各生长性状种源间变异对总变异的贡献为95.77%,种源内无性系间的贡献为4.23%,说明黄波椤群体间的遗传变异大于个体间.
猜你喜欢
东北林业大学学报(2023年6期)2023-05-31 02:56:16
丽水学院学报(2022年2期)2022-04-19 01:22:10
草业学报(2022年3期)2022-03-26 02:27:28
林业科技情报(2021年3期)2021-09-01 02:01:26
当代陕西(2019年7期)2019-04-25 00:22:54
中南林业调查规划(2017年1期)2017-12-19 00:58:07
中国实验诊断学(2017年5期)2017-06-05 15:03:23
学习月刊(2016年4期)2016-07-11 02:54:26
上海精神医学(2014年6期)2014-12-08 08:14:51
大众考古(2014年3期)2014-06-26 08:30:44
