
罗伊氏乳酸杆菌SL001对AD模型小鼠和C57BL/6小鼠肠道微生物的影响
2020-04-15 13:33刘梅梅胡汝倩郭耀文孙卫忠李建涛范明亮王燕杜华茂唐志如柴春利
生物工程学报 2020年9期
刘梅梅,胡汝倩,郭耀文,孙卫忠,李建涛,范明亮,王燕,杜华茂,唐志如,柴春利
1 西南大学 生物技术学院,重庆 400715
2 西南大学 动物科技学院,重庆 400715
阿尔茨海默症 (Alzheimer’s disease,AD) 是一种神经退行性疾病,具有年龄相关性,其临床表现为进行性认知障碍,始于记忆力的恶化,影响正常生活并最终导致残疾甚至死亡[1-3]。目前对AD的发病机制主要有两个假说:Aβ沉积假说和Tau蛋白过度磷酸化假说[4-5]。令人遗憾的是,目前以这两种假说为中心的治疗策略持续失败。最近,越来越多的研究表明,肠道微生物 (Gut microbes,GM) 与AD之间存在紧密的联系[6-8]。与常规饲养的AD模型小鼠相比,无菌AD模型小鼠表现出异常的肠胃蠕动,并且脑中Aβ沉积急剧降低。将常规饲养的AD模型小鼠的粪便微生物菌群定殖到无菌AD模型小鼠中,小鼠脑中的Aβ水平增加。相比之下,来自对照野生型小鼠的微生物群定殖却不会造成Aβ水平的增加[9]。长期的广谱抗生素治疗也可使AD模型小鼠的肠道微生物多样性发生紊乱,进而引发脑部Aβ斑块沉积和小胶质细胞形态发生改变[10]。近来的临床试验表明,幽门螺杆菌Helicobacter pylori感染的AD患者具有更严重的认知障碍,根除幽门螺杆菌可以延长AD患者的生存期[11]。GV-971药物已通过临床Ⅲ期试验,发现其可以重塑AD患者的肠道微生物,降低大脑的炎症反应,进而改善了认知功能的下降[12],该结果进一步证明了肠道微生物在AD发展中的重要作用,因此可以将GM作为AD治疗的潜在新靶标。
益生菌作为生物治疗剂具有改善肠道环境、抗肥胖、预防癌症和感染等功效[13-14]。一些益生菌也可以通过微生物-肠-脑轴 (Microbe-gut-brain axis,MGB) 影响中枢神经系统 (Central nervous system,CNS)。MGB轴是一种双向通讯系统,而作为MGB轴核心的微生物,可通过神经、内分泌、代谢和免疫途径影响中枢神经系统[15]。已有研究结果显示一部分乳酸杆菌可以影响肠道神经元的兴奋性和结肠运动性,并且降低血浆中炎症因子水平,缓解小鼠的焦虑行为[16]。在AD患者脑内检测到γ-氨基丁酸 (γ-aminobutyric acid,GABA) 浓度的降低,而短乳杆菌和齿状双歧杆菌可通过谷氨酸代谢途径产生GABA,增加其在脑内的含量,减少神经损伤[17]。益生菌治疗还可以挽救AD模型小鼠的认知缺陷,改善某些代谢状态[18]。综上所述,某些益生菌可能能够安全有效地治疗AD。
在本研究中,我们对AD模型小鼠与野生型小鼠的肠道微生物群落差异进行了调查,并分析添食罗伊氏乳酸杆菌SL001 (Lactobacillus reuteriSL001,L.reuteriSL001) 对两种小鼠肠道微生物菌群结构的影响。
1 材料与方法
1.1 罗伊氏乳酸杆菌SL001菌液制备
L.reuteriSL001 (NCBI登录号KU255544.1;菌种保藏号CCTCC M 2019122) 为本实验室前期从家兔胃液内容物中筛选所得。该菌株为革兰氏阳性杆状细菌,在MRS培养基上呈白色圆形,中央凸起,边缘整齐。同时,该菌株具有耐酸、耐胆盐、分解胆固醇的特点。在MRS培养基中培养L.reuteriSL001,离心收集细胞沉淀,并用PBS (pH 7.4) 洗涤。将收集的沉淀物以5×1011CFU/mL的浓度重悬于PBS中以供实验使用。
1.2 动物试验
APP/PS1双转基因小鼠 (C57BL/6 background)购于南京大学模式动物研究所,野生型 (C57BL/6品系) 小鼠购自重庆中医药研究院。将7月龄20 g左右的雄性APP/PS1双转基因小鼠和野生型小鼠随机分为4组,每组5只:AD小鼠添食组 (PT)、AD小鼠未添食组 (PC)、野生型小鼠添食组 (NT)、野生型小鼠未添食组 (NC)。添食组小鼠每天用0.2 mL的L.reuteriSL001菌悬液灌胃,未添食组则每天接受等量的无菌PBS,每天早上9点通过口服管饲法进行给物处理,周期为45 d。将小鼠分笼饲养,室内适度通风,室温25 ℃,12 h光照/12 h黑暗循环,自由饮用食物和水。
1.3 小鼠粪便16S rRNA
添食结束后所有小鼠禁食12 h,经乙醚麻醉后解剖,取直肠粪便,液氮冻存以备使用。采用TIANamp基因组提取试剂盒 (TIANGEN Biotech LTD) 提取小鼠粪便DNA,PCR扩增16S rRNA V3–V4片段,引物序列为341-F (5′-CCTAYGGG RBGCASCAG-3′) 和806-R (5′-GGACTACNNGG GTATCT-3′)[19-20]。反应条件:预变性98 1 min℃;98 ℃ 10 s,50 ℃ 30 s,72 ℃ 30 s,30个循环;72 ℃最终延伸5 min。PCR产物经琼脂糖凝胶电泳分离,胶回收试剂盒 (Qiagen,Germany)纯化目的片段,送北京诺禾致源生物有限公司用Illumina MiSeq 250平台测序。
1.4 小鼠肠道菌群分析
16S rRNA测序后拼接成Tag,并根据Tag进行物种分类、OTU分析、多样性分析和多样品的比较分析等。利用Mothur对测序序列进行去冗余处理,使用基于Naive Bayesian的分类器RDP Classi-fier工具对Tag进行物种注释。计算97%相似度下的OTU数量,并根据众数原则对OTU进行物种注释。采用PICRUSt进行OTU pathway注释,以Mothur进行样品α多样性分析;采用R软件进行热图分析、β多样性分析、样品聚类分析和维恩图的绘制;采用SPSS 21软件中的双因素方差分析检验两个处理组之间的统计学差异,*P<0.05,**P<0.01,***P<0.001。
2 结果与分析
2.1 测序数据
经整理,4个处理组20只小鼠肠道微生物的16S rRNA基因扩增子的Illumina MiSeq 250测序对样品产生84 694–100 129个原始读数。在质量过滤和去除嵌合序列后,每个样品保留80 010–92 264个读数,以97%相似度划分,可以分配给1 176个OTUs。根据OTU数据,绘制稀疏曲线,由图1可以看出,稀疏曲线的末端趋于平坦并且在0.03相异截止值下,所有样品的测序深度指数覆盖率均高于99.7% (表1)。这些结果表明我们的抽样量充分,并且测序结果捕获了小鼠肠道微生物群落的大部分多样性。根据物种注释的结果,将来自小鼠粪便样本中所有的OTU分为对应的域,包括了19个门、40个纲、64个目、104个科和175个属。由表1可以看出,OTU数目NT 组>PT组>PC组>NC组,表明添食L.reuteriSL001可以增加小鼠肠道微生物种类。

图1 样品物种的稀释曲线Fig.1 Rarefaction curve analyses of sample.
2.2 小鼠肠道细菌群落的多样性分析
2.2.1 Alpha多样性分析
由表1可知,在ACE、Chao1、Shannon、Simpson等α多样性指数中,NT>NC,PT>PC,这揭示了添食L.reuteriSL001可以增加小鼠肠道菌群的多样性。在PC与NC中,前者ACE、Chao1、Shannon、Simpson等α多样性指数更高,表明未添食AD模型小鼠的肠道微生物多样性比野生型高;在PT与NT中,ACE、Chao1、Shannon、Simpson等指数差异很小 (甚至后者更大),说明野生型小鼠对L.reuteriSL001添食更敏感。AD模型小鼠的肠道微生物多样性比野生型丰富,添食L.reuteriSL001可以略微增加小鼠肠道菌群的多样性,并且在野生型小鼠中的涨幅较大。

表1 小鼠肠道微生物丰度和多样性指数Table 1 Intestinal microbial abundance and alphadiversity index in mice
2.2.2 Beta多样性分析
由图2可以看出,NT组和PT组表现出了较明显的组内聚集,样品根据所处组别相互聚集到一起,NC组和PC组并没有依据其组别明显聚集,添食组的组内微生物结构与组成的相似性高于未添食组,说明添食L.reuteriSL001可以在一定程度上减少组内个体间的差异。

图2 PCoA聚类分析Fig.2 Cluster analysis by principal co-ordinates analysis.
由图3可知,PC组和NC组 (图3A) 之间,组内差异大于组间差异,但不显著 (R=–0.052,P=0.538)。PT组与NT组 (图3B) 之间,组间差异大于组内差异 (R=0.184,P=0.11),表明添食L.reuteriSL001可以使同种小鼠的肠道微生物更趋于一致,保持更稳定的状态。

图3 小鼠肠道细菌群落的Anosim分析 (A:NC组与PC组间比较;B:NT组与PT组间比较)Fig.3 Anosim analysis of intestinal bacterial community in mice.(A) Comparison between NC group and PC group.(B)Comparison between NT group and PT group.
如图4所示,4个处理组小鼠共聚为两大分支,PC组和NC组之间的细菌群落结构相似性较高,共同聚集在上方,PT组和NT组之间的细菌群落结构相似性较高,共同聚集在下方,说明添食L.reuteriSL001可以造成不同基因型小鼠肠道微生物群落结构发生相似的改变。

图4 小鼠肠道细菌群落聚集分析图Fig.4 Clustering analysis of mice intestinal bacterial community.
2.3 样品群落组成分析
2.3.1 物种维恩图分析
由维恩图 (图5) 可知,4个处理组之间共有的OTU数目为667个,占绝大多数。NT组独有OTU数目最多,为93个,PC组独有的OTU数目最少,仅有28个,PT组和NC组独有OTU数目分别为50个和54个,即AD模型小鼠初始肠道菌群丰富度小于野生型小鼠,而添食L.reuteriSL001能增加肠道菌群丰富度,野生型小鼠尤为显著。另外,AD模型小鼠添食后,与野生型小鼠未添食组之间的共有OTU数目增加。这可以说明添食L.reuteriSL001能够改变小鼠肠道微生物结构组成,增加独有物种,丰富物种多样性,也可以使AD模型小鼠的肠道微生物组成更接近于正常小鼠。

图5 小鼠肠道微生物OTU程度的维恩图Fig.5 Venn diagram of OTU degree of intestinal microorganisms in mice.
2.3.2 细菌门分类水平的比较
4组小鼠粪便微生物中的优势菌群均为拟杆菌门 (Bacteroidetes) 和厚壁菌门 (Firmicutes),这两个菌门序列约占序列总数92% (图6A)。AD模型小鼠中的拟杆菌门丰度低于野生型,厚壁菌门的丰度高于野生型,添食L.reuteriSL001会增加拟杆菌门所占比例,且PT组与PC组之间 (68.54%vs49.20%,P<0.05)、PT与NC之间 (68.54%vs54.38%,P<0.01) 存在显著性差异。相反,添食L.reuteriSL001会降低厚壁菌门所占的比例,而且PT与PC之间 (24.15%vs42.80%,P<0.05)、PT与NC之间 (24.15%vs38.12%,P<0.05) 存在显著性差异。为了更直观地展现两种菌门的占比差异,统计了厚壁菌门与拟杆菌门的比值 (图6B)。其中PC组小鼠的F/B比值显著高于NC组,添食L.reuteriSL001会降低比值且PT组比值最小,并且PT组分别与PC组、NC组之间有显著性统计学差异。
2.3.3 细菌目分类水平的比较
从图7可以看出,在目的水平上,拟杆菌目(Bacteroidales) 的相对丰度最大,并且AD模型小鼠的相对丰度低于野生型小鼠,添食L.reuteriSL001会增加其相对丰度,与图6结果相一致。AD模型小鼠中的乳酸杆菌目 (Lactobacillales)、芽胞杆菌目 (Bacillales) 和双歧杆菌目 (Bifidobacteriales)的相对丰度均低于野生型,添食L.reuteriSL001会增加这3个目在AD模型小鼠中的相对丰度,但在野生型中则是降低的效果。AD模型小鼠中的丹毒丝菌目 (Erysipelotrichales) 相对丰度高于野生型,添食L.reuteriSL001会降低其在两种小鼠中的相对丰度。
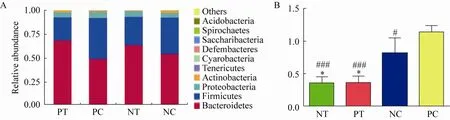
图6 门类群落结构组成分布图 (A:门水平前10种细菌的相对丰度;B:Firmicutes/Bacteroidetes (F/B) 在不同组中的比值情况.*:P<0.05;**:P<0.01;***:P<0.001 vs NC;#:P<0.05;##:P<0.01;###:P<0.001 vs PC)Fig.6 Distribution structure of community structure.(A) Relative abundance of the top 10 bacterial at the level of phylum.(B) The ratio of Firmicutes/Bacteroidetes (F/B) in different groups (*:P<0.05;**:P<0.01;***:P<0.001 vs NC;#:P<0.05;##:P<0.01;###:P<0.001 vs PC).

图7 目水平上前10种细菌的相对丰度Fig.7 Relative abundance of the top 10 bacterial at the level of order.
2.3.4 细菌属分类水平的比较
从图8可以看出,在属水平上,4组小鼠的Allobaculum、毛螺旋菌属 (Lachnospiraceae_NK4A136_group)、拟杆菌属 (Bacteroides) 和乳酸菌属 (Lactobacillu) 所占比例较大,相对丰度高。AD模型小鼠的拟杆菌属、乳酸菌属、双歧杆菌属 (Bifidobacterium) 和拟普雷沃菌属(Alloprevotella) 的相对丰度低于野生型小鼠,添食L.reuteriSL001会增加这些属在AD模型小鼠中的丰度,降低在野生型小鼠中的丰度。AD模型小鼠的丁酸弧菌属 (Butyrivibrio) 相对丰度也低于野生型,但是添食L.reuteriSL001后,其相对丰度在两种小鼠中均出现降低。
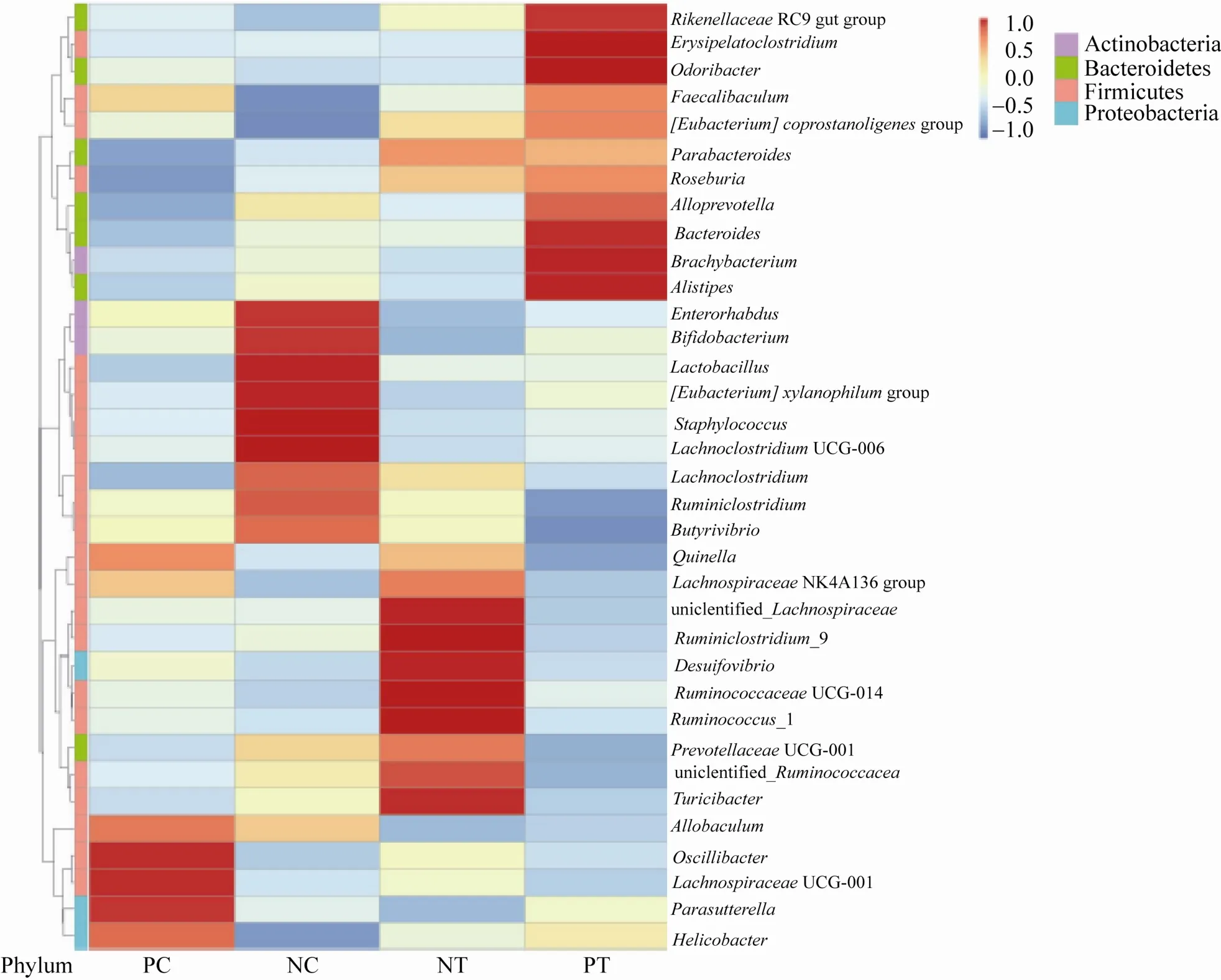
图8 含系统发育进化树的热图(属水平)Fig.8 Heatmap graph with phylogenetic tree (genus level).
2.4 肠道菌群LEfSe分析
通过对PC组和NC组进行LEfSe分析可知(图9A–B),野生型小鼠的优势菌种为杆菌纲 (Bacilli)中的乳酸杆菌科 (Lactobacillaceae),在AD模型小鼠中未发现优势菌群。在PT组和NT组的比较中(图9C),均未发现丰度差异显著的菌种,表明添食L.reuteriSL001可减小AD模型小鼠与野生型小鼠之间的差异。

图9 LEfSe分析 (A:PC组与NC组组间LDA值分布柱状图;B:PC组与NC组组间进化树分支图;C:PT组与NT组组间进化树分支图)Fig.9 LEfSe analysis.(A) Histogram of LDA value distribution between PC group and NC group.(B)Phylogenetic tree branch diagram between PC group and NC group.(C) Phylogenetic tree branch diagram between PT group and NT group.
2.5 物种分类树分析
由4个处理组不同比例的物种分类树 (图10)可以看出,野生型小鼠在杆菌纲 (Bacilli) 以及乳酸杆菌目、芽孢杆菌目、乳酸杆菌科中的比例占有绝对优势,AD模型小鼠占有比例较小。添食L.reuteriSL001后,AD模型小鼠在以上菌群中的占有比例增加。

图10 小鼠肠道微生物物种分类树Fig.10 Species classification tree of intestinal microorganisms in mice.
3 讨论
在本研究中,我们调查了AD模型小鼠(APP/PS1双转基因小鼠)和野生型小鼠 (C57BL/6)之间的肠道菌群差异,分析了L.reuteriSL001对两种小鼠肠道微生物菌群结构的影响。α多样性分析的结果表明,AD模型小鼠的α多样性大于野生型小鼠,但是两者之间的差异较小。添食L.reuteriSL001后,两种小鼠的α多样性均增加,并且AD模型小鼠的增幅较小。这可能是因为AD模型小鼠自身独有的某些微生物,会对外来微生物的生长产生抑制作用,也可能是因为某些微生物的代谢产物改变了小鼠肠道环境,使部分微生物更加不易定殖繁殖,这也说明AD模型小鼠的肠道菌群不易调节,而这也成为一些治疗手段的阻碍因素。β多样性分析结果显示,AD模型小鼠和野生型小鼠没有出现明显的组内聚集,组内差异大,但是添食L.reuteriSL001后均表现出较明显的组内聚集,说明L.reuteriSL001可以在一定程度上减少组内个体间的差异,使同种小鼠的肠道微生物更趋于一致,保持更稳定的状态。
在我们的测序数据中,所有样品的优势菌门都为拟杆菌门和厚壁菌门,两种菌门约占序列总数92%。厚壁菌门和拟杆菌门与体重有着密切联系[21],而且厚壁菌门与拟杆菌门的比值 (F/B) 增加可能会导致过多的免疫反应和过量饮食,促进能量在体内的富集,造成肥胖[22-23],所以F/B的比值也是肥胖和2型糖尿病 (T2DM) 的明显标志[24]。有趣的是,阿尔兹海默症被称作为3型糖尿病[25],并且其他研究也表明,相对于野生型,AD模型小鼠的F/B比值增加[12]。因此,F/B的比值增加可能指示着患AD风险也在增加[26]。由图5B可以看出,AD模型小鼠中的拟杆菌门丰度低于野生型,厚壁菌门的丰度高于野生型,添食L.reuteriSL001会造成两种小鼠的F/B比值下降,而且AD添食组与AD未填食组存在显著性差异,表明L.reuteriSL001对AD模型小鼠有潜在的有益作用。
在目水平上,AD模型小鼠中的拟杆菌目、乳酸杆菌目、芽胞杆菌目和双歧杆菌目的相对丰度均低于野生型,添食L.reuteriSL001可增加这4种目在AD模型小鼠中的相对丰度。拟杆菌、乳酸杆菌和双歧杆菌可以产生短链脂肪酸 (Short-chain fatty acid,SCFAs),包括乙酸盐、丁酸盐和丙酸盐。研究表明乙酸盐可以增强全身葡萄糖稳态,也能穿过血脑屏障,向大脑发出饱足感[26]。丁酸盐是一种多功能分子,可以保护神经,恢复血脑屏障的功能,具有明显的认知和抗抑郁作用[27]。在AD模型中,丁酸钠的使用可显著改善小鼠的学习记忆能力[28-29]。丙酸盐能影响小胶质细胞的活化并降低血脑屏障的渗透性,也可防止不适当的免疫反应[30-31]。综上所述,SCFAs水平的降低可能与AD的发展密切相关。添食L.reuteriSL001可增加产生SCFAs的细菌,所以L.reuteriSL001可能对AD的发展具有缓解抑制作用。在AD患者的额叶、颞叶和顶叶皮质均发现GABA 水平降低,而肠道中的某些双歧杆菌和乳杆菌可能将谷氨酸盐转化为GABA,调节机体的认知行为[17]。另外,服用某些乳酸杆菌和双歧杆菌可以恢复AD模型小鼠脑海马中神经营养因子的水平,抑制Aβ斑块的产生并改善学习记忆能力[32-33]。芽胞杆菌具有产生乙酰胆碱的能力,而乙酰胆碱是AD患者大脑中减少的一种神经递质,所以芽胞杆菌可能会通过迷走神经帮助胆碱信号的传递[34-36]。我们的研究表明,L.reuteriSL001可以增强以上细菌在AD模型小鼠中的相对丰度,这可能会降低脑部的免疫反应、Aβ斑块的沉积和小胶质细胞的过度活化,进而改善学习记忆能力。L.reuteriSL001是否会通过以上途径缓解AD病症还需要进一步的实验研究。
在属水平上,AD模型小鼠的拟普雷沃菌属和丁酸弧菌属相对丰度均低于野生型小鼠,添食L.reuteriSL001会增加拟普雷沃菌属在AD模型小鼠中的丰度,降低丁酸弧菌属的丰度。拟普雷沃菌属也可以产生短链脂肪酸,发挥对生物体有益的作用。丁酸弧菌属可以产生内毒素,内毒素能够引起炎症反应、损害肠屏障、引起人体代谢与免疫失调,从而诱导肥胖、胰岛素抵抗、糖尿病等代谢性疾病[37-38]。当机体衰老时,胃肠粘膜和血脑屏障的渗透性增加,内毒素更容易到达大脑,诱发神经炎症和Aβ增加,进而引起AD疾病的发展[39-40]。有研究表明 AD 患者血浆中的内毒素含量远远高于正常人[41],所以减少内毒素的产生可能会对AD的治疗起到积极作用。添食L.reuteriSL001增加了对改善病症有益的细菌,减少了加剧AD病理的细菌,表明L.reuteriSL001可以调节AD模型小鼠的肠道微生物结构,可能对生物体的生理功能产生积极影响。
通过LEfSe分析可知,野生型小鼠的优势菌种为杆菌纲中的乳酸杆菌科和乳酸杆菌属,在AD模型小鼠中未发现优势菌群,这一结果也与物种分类树结果相符。在添食L.reuteriSL001后,在两种小鼠中均未发现丰度差异显著的菌种,说明L.reuteriSL001干预可以改善AD模型小鼠的肠道微生物结构,使其更接近野生型小鼠。有其他研究报道AD模型小鼠肠道中丰度差异显著的菌种为中紫单胞菌科的帕拉普氏菌 (Paraprevotellaceae)和普氏菌属 (Prevotella)[42],与我们本次实验结果不一致,这有可能是因为选择的AD模型小鼠以及鼠龄不同等多方面原因所导致。我们的结果可以丰富AD模型小鼠的肠道微生物研究,并具有一定的参考价值。若益生菌干预能够重塑肠道平衡,减少患AD的风险,可为以后的AD治疗提供一种安全可靠的策略。
猜你喜欢
安徽农业科学(2022年20期)2022-11-11
科学技术与工程(2022年26期)2022-11-01
今日农业(2022年14期)2022-09-15
中老年保健(2022年2期)2022-08-24
新疆医科大学学报(2022年6期)2022-07-02
中国农学通报(2022年14期)2022-06-01
油气田环境保护(2022年2期)2022-05-09
中国饲料(2022年5期)2022-04-26
昆明医科大学学报(2022年3期)2022-04-19
科教新报(2020年21期)2020-06-05
