
Toll样受体通路调节Tregs功能的研究进展
2020-04-15 13:32:50郭鹏张含李长菲孟颂东
生物工程学报 2020年9期
郭鹏,张含,李长菲,孟颂东
1 中国科学院微生物研究所 中国科学院病原微生物与免疫学重点实验室,北京 100101
2 中国科学院大学,北京 100049
1 Toll样受体
1.1 Toll样受体及其配体
Toll是果蝇中决定胚胎背腹轴线发育、参与抗感染的一组基因。通过数据库搜寻同源物,科学家们很快在哺乳动物中发现结构相似的一个跨膜分子家族,主要包括IL-1R和Toll类似物,后者又被称为Toll样受体 (Toll like receptors,TLRs)。TLRs是Ⅰ型跨膜蛋白,细胞外结构域含有形似马鞍状的亮氨酸富集重复序列 (Leucine rich repeat,LRR),该重复区域构成配体结合区。细胞质结构域为TIR(Toll/IL-1R),是所有TLRs及IL-1R分子胞内段所特有的。TIR结构域与细胞内其他带有相同TIR结构域的分子相互作用,由后者启动下游信号传递。TLRs主要表达于免疫细胞,包括巨噬细胞、树突状细胞 (Dendritic cell,DC)、B细胞和T细胞。在一些非免疫细胞如上皮细胞、内皮细胞[1]和成纤维细胞中也有表达。目前在哺乳动物中已发现13种TLRs,其中小鼠和人均表达TLR1–9,TLR10仅存在于人体内,TLR11–13仅在小鼠体内被发现[2]。TLRs是一种模式识别受体,不同的TLRs识别不同的病原相关分子模式 (Pathogen-associated molecular pattern,PAMP)[3]。TLRs与PAMPs相互作用主要表现为两个特点:1) 在细胞中因表达部位、蛋白组成等差异,不同的TLRs分子识别不同的PAMPs。2) 胞膜TLRs通常以二聚体形式发挥作用,相组合的TLRs一般识别同类配体分子。在体内除了外源性配体外,一些TLRs结合内源性配体,如TLR2和TLR4可以与细胞内高表达的热休克蛋白70、热休克蛋白gp96和HMGB1相互作用,这与坏死细胞释放内容物作为危险相关分子 (Damage-associated molecular patterns,DAMP) 有关[4]。此外,有研究报道TLR4还与体内纤连蛋白、纤维蛋白原、肺表面活性蛋白A、肝素、透明质酸和鼠的β防御素相互作用[2]。在系统性红斑狼疮 (Systemic lupus erythematosus,SLE) 中,TLR9通过与染色质免疫球蛋白G复合物结合被激活。这些内源性配体的存在,增加了TLRs参与自我耐受和免疫检测的可能性,提示TLRs对于自身免疫病和癌症的治疗至关重要。
1.2 Toll样受体信号通路
TLRs是连接天然免疫和获得性免疫的桥梁,它对于PAMPs的识别及下游信号通路的激活是机体免疫应答的重要途径之一。TLRs介导的信号通路激活需要衔接蛋白的参与,衔接蛋白主要包括髓样分化因子88 (Myeloid differentiation factor 88,MyD88)、MyD88样衔接蛋白(MyD88 adapter-like,MAL)、诱导β干扰素的TIR结构域衔接蛋白(TIR-domain-containing adaptor protein inducing IFN-β,TRIF)、TRIF相关衔接分子(TRIF-related adaptor molecule,TRAM) 和含不育α和犰狳基序的蛋白 (Sterile alpha and armadillo-motif-containing protein,SARM)[5]。TLRs信号通路可分为MyD88依赖和非依赖途径(图1)。TLR5、TLR7和TLR9等分子直接与MyD88作用激活下游信号通路,而TLR2和TLR4则需要MyD88和含TIR结构域的衔接蛋白 (也称MAL,TIR domain containing adaptor protein,TIRAP/Mal) 的共同作用,促进下游通路的激活。此外,TLR3和TLR4都可介导非MyD88依赖途径的信号转导,其中TLR3只能利用该途径。TLRs信号通路激活主要引起NF-κB(核转录因子)活化,诱导IL-1β、IL-6、TNF-α等炎性细胞因子的分泌和Ⅰ型干扰素的表达,参与炎症反应或炎症病理反应[6],同时调节免疫反应。除此之外,TLRs还能通过激活丝裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPKs)、胞外信号调节激酶(Extracellular regulated protein kinases,ERK)、p38和c-JUN N端激酶 (c-Jun N-terminal kinase,JNK)等调节细胞的增殖、转化和死亡[7]。

图1 Toll样受体介导的信号通路Fig.1 Toll like receptors mediated signaling pathway.TLRs signaling pathways can be divided into MyD88-dependent and MyD88-independent pathways.Different from TLR5,TLR7,and TLR9,which can directly interact with MyD88,TLR2 and TLR4 require coordination of MyD88 and TIRAP/Mal for downstream signaling.TLR3 and TLR4 mediate signal transduction in a MyD88-independent pathway,of which TLR3 solely utilize this pathway.TLRs signaling mainly causes activation of NF-κB that induces the secretion of inflammatory cytokines including IL-1β,IL-6,TNF-α and the expression of type I interferon.In addition,TLRs can also activate p38 and JNK.
2 Treg细胞
调节性T细胞 (Treg) 又称抑制性T细胞,是一类负反馈性T细胞亚群。Treg主要分成两类:一类是源自胸腺的自然调节性T细胞 (Natural Treg,nTreg),代表亚群为CD4+CD25+Treg,占CD4+T细胞亚群的5%–15%。CD4+CD25+Tregs激活后能够抑制T细胞增殖、细胞因子的分泌和抗原呈递细胞 (Antigen presentation cell,APC)的功能[8]。它们可以识别自身和非自身的抗原肽表位。这类Tregs通过TGF-β诱导,表达转录因子叉头框蛋白3 (Forkhead box protein 3,Foxp3)[9],Foxp3是Treg的标志性分子。除此之外小鼠和人的CD4+CD25+Treg还表达很多标志分子,如细胞毒性T淋巴细胞相关蛋白4 (Cytotoxic T-lymphocyte-associated protein 4,CTLA-4)[10]、糖皮质激素诱导的肿瘤坏死因子受体(Glucocorticoid-induced tumor necrosis factor receptor,GITR)[11]、CD103[12]、CCR8、淋巴细胞活化基因-3分子 (Lymphocyte activation gene-3,LAG-3)、神经纤毛蛋白-1 (Neuropilin-1,Nrp1)、高表达CD5分子和低表达IL-7受体分子CD127。第二类Treg是分布在外周中的Treg,也被称为诱导型调节性T细胞 (Inducible Treg,iTreg),可在外周通过TGF-β或IFN-γ单独诱导初始T细胞产生,这类细胞也称为适应性调节T细胞 (Adaptive Treg,aTreg),主要产生IL-10和TGF-β。aTreg有两种类型:Tr1和Th3细胞。Tr1细胞可以在IL-10、不成熟DC反复刺激或抗CD3和CD46单抗联合刺激等条件下产生[13]。Tr1细胞可以大量分泌IL-10和IFN-γ,并且通过IL-10来发挥负调节作用[14];Th3是由自然的CD4+T细胞在抗原和TGF-β刺激下分化而来,同时Th3通过分泌TGF-β抑制免疫反应,在诱导外周耐受和黏膜免疫中发挥作用[15-16]。Tr1和Th3等诱导型调节性T细胞主要通过分泌可溶性细胞因子或细胞接触依赖机制发挥免疫抑制功能,而自然调节T细胞主要通过细胞接触依赖机制发挥抑制功能。
Treg在免疫系统中扮演了重要角色,正常生理条件下,Treg可以维持细胞外周耐受和调节自身免疫反应[17]。但在急性感染期和肿瘤消除期,Treg会抑制效应T细胞的活力,阻碍免疫反应[18-19]。因此,研究如何调节Treg的功能对于维持机体自身免疫平衡、改善疾病治疗效果具有重要意义。近年来研究者们发现Treg表达不同种类的TLRs分子[20],TLRs介导的信号转导显著影响Treg的分化、增殖与功能,接下来重点介绍一下TLRs和Treg之间的联系。
3 TLRs和Treg
TLRs信号通路以直接或间接的方式调控CD4+CD25+Treg的功能。间接调控主要通过APC介导的TLRs信号通路调节Treg功能。而Treg上的TLRs也可以通过与相应配体的结合直接影响自身的功能。相比于其他CD4+T细胞,Treg细胞中的TLRs表达水平更高[21-22],这提示Treg细胞更容易受到TLR配体的调控。Caramalho等比较分析了C57BL/6小鼠Treg和Tconv (Conventional T)细胞中TLRs的表达情况,结果表明两者均表达TLR1、TLR2和TLR6,而TLR4、TLR5、TLR7和TLR8在Treg细胞中选择性表达,TLR3和TLR9 mRNA在小鼠CD4+T细胞亚群中没有检测到[20]。Gelman等发现在BALB/c小鼠中活化的CD4+T细胞表达TLR3、TLR5和TLR9,但不表达TLR2和TLR4[23]。这种差异可能是由于使用的检测方法、T细胞纯度或者小鼠的亚系不同所造成的。Chiffoleau等发现TLR9 mRNA在大鼠CD4+T细胞中表达,TLR5 mRNA在CD4+CD25+T细胞中高度表达[24]。人类CD4+CD25+Treg表达更高水平的TLR2、TLR5和TLR8[25]。不同的TLRs对Treg的影响不同,作用方式也不同。
3.1 TLR2对Treg细胞的调节
近来很多研究表明,TLR2在调节Treg功能方面发挥了关键作用。TLR2是T细胞的共调控受体[1],在体外,天然免疫细胞TLR2分子的活化可以促进细胞分泌IL-10[26],而且与体内抑制炎症反应有直接的关系[27]。APC和Treg细胞上的TLR2都会调控Treg的功能。APC上的TLR2活化后会促进细胞分泌IL-1和IL-6,逆转CD4+CD25+Treg的低反应性[28]。Treg上的TLR2激活后直接调控Treg功能。研究表明TLR2基因缺陷小鼠的Treg数目与TLR4基因缺陷小鼠相比明显减少[22],并且用TLR2配体Pam3Cys刺激野生小鼠,不但会使Treg数目增加、CD25分子表达上调,而且在此期间Treg的抑制显型也会短暂消除[29]。随后的研究表明小鼠Treg细胞中的TLR2信号激活会增加Treg糖酵解和增殖并降低其抑制能力[30]。而且Lal等发现肽聚糖 (Peptiglycan,PGN) 能够激活TLR2-MyD88-IRF1 (Interferon regulatory factor 1,干扰素调节因子1)信号通路。IRF1活化后与Foxp3基因座的近端启动子、内含子、增强子区域中存在的IRF1反应元件结合,负调控Foxp3的转录,以抑制Treg的功能[31]。此外,不同的TLR2配体对Tregs功能有不同的影响。研究表明HSP60可以通过Treg上的TLR2,活化信号分子PKC、PI3K和P38上调Treg的抑制功能[32]。然而人工合成的脂肽Pam3Cys、FSL-1和Pam2Cys通过增强AKT/PKB (Protein kinase B,蛋白激酶B) 磷酸化减弱Treg的抑制功能[33]。Treg在受到TLR2配体刺激后,会促使自身分泌IL-6和TGF-β,不仅会使Treg细胞的抑制功能减弱,还会促进Treg细胞向分泌IL-17的表型Th17分化[34]。人体内存在IL-17+Foxp3+循环记忆样Treg细胞,当机体受到外界病原体感染时,会激活这类细胞,增强免疫反应,更有效地应对感染[35]。因此TLR2活化是一把双刃剑,一方面限制Treg抑制活力,促进免疫反应的开始;另一方面增加Treg的数量,阻碍病原体和癌细胞的最终消除。
TLR2在很多自身免疫病中发挥着重要的作用,TLR2信号活化提高Treg的免疫调节功能进而预防Ⅰ型糖尿病[36];在类风湿性关节炎小鼠模型中,TLR2敲除鼠Treg的抑制功能降低,T细胞的IFN-γ产生大大增加,显示出更严重的关节炎[37]。这些研究表明TLR2介导的通路在活化Treg抑制功能方面起重要作用。
3.2 TLR4对Treg细胞的调节
目前,TLR4分子对Treg细胞功能的影响仍未完全确定。Caramalho等的研究表明TLR4配体脂多糖 (Lipopolysaccharide,LPS) 可以直接促进Treg的生存和增殖,使Treg的免疫抑制功能增强10倍。此外LPS还能上调如CD69、CD44、CD38等Treg活性标记分子的表达水平[20]。随后他们发现用LPS处理非肥胖性糖尿病小鼠可以增加Foxp3+和CD103+Treg的数量和活力,有助于糖尿病的防护[38]。此外,用不同剂量的LPS预先处理小鼠,会诱发小鼠出现迟发型超敏反应、移植物抗宿主反应、移植排斥反应,并且显著降低正常免疫应答的水平,这可能也是由于LPS活化Treg导致的。随后研究发现在小鼠肠炎模型中,TLR4和IL-2缺陷小鼠的Treg表达高水平的炎性细胞因子,包括GM-CSF、IFN-γ和IL-17,并且其抑制活力也有所降低[39]。这些研究都提示TLR4通过与配体的作用直接介导Treg的增殖与活化。
TLR4分子还可以通过间接作用的方式调控Treg的功能。Cao等在肠炎模型中发现TLR4以MyD88依赖的方式负调节Treg的产生[40],Treg减少的原因是在受到肠内配体刺激后,会促进Foxp3+Treg向IL-17+和IFN-γ+Foxp3-细胞分化。然而2018年有研究发现热休克蛋白HSP60通过激活TLR4-Mal (MyD88衔接样蛋白) 信号转导通路,促进巨噬细胞中TGF-β的表达,进而诱导Treg的产生[41]。先前也有报道称,LPS能够通过TLR4刺激树突状细胞产生IL-6,IL-6直接作用于效应T细胞,使其对于Treg的抑制功能产生抗性[28]。还有研究提示高度纯化的LPS并不影响小鼠Treg功能[42]。上述不同的实验结果可能是由于采用的动物模型、实验方法等差异导致的,同时也提示了TLR4对Treg功能影响可能还受其他免疫细胞和微环境的制约,因此,TLR4介导的Treg细胞调控机制还需进一步研究。
3.3 TLR5对Treg细胞的调节
Crellin等证明人类CD4+CD25-T细胞和CD4+CD25+T细胞均表达TLR5,在CD4+CD25+T细胞中TLR5mRNA水平明显高于CD4+CD25-T细胞,但二者TLR5分子蛋白水平无明显差异,其具体机制还有待探索[21]。TLR5配体鞭毛蛋白(Flagellin)[43]会直接影响人类CD4+CD25+T细胞和CD4+CD25-T细胞的功能,在黏膜免疫调控中发挥重要作用。多克隆刺激低反应性是Treg细胞的主要特征。研究表明TLR5配体鞭毛蛋白不改变Treg的低反应性。相反,它作为共刺激分子促进CD4+CD25-效应T细胞的增殖和IL-2的产生。然而鞭毛蛋白在不存在APC的情况下,会增强Treg细胞的免疫抑制功能和Foxp3的表达,这种TLR5介导的Treg的抑制功能增强可能与Foxp3表达升高有关[44]。在小鼠移植肿瘤模型中,使用鞭毛蛋白处理小鼠会加速肿瘤的生长,这主要与IFN-γ和IL-4比率降低以及Treg细胞频率增加有关。但是在肿瘤移植8–10 d后,再用鞭毛蛋白处理会抑制肿瘤的生长,此时小鼠IFN-γ和IL-4比率增加,但Treg细胞频率却降低[45]。这种TLR5配体对于Treg细胞和效应T细胞的影响差异机制还需要进一步的研究。另有研究报道重组鞭毛蛋白rFliC给药可延长同种异体移植物的存活时间,这可能与TLR5激活Treg有关[46]。综上所述,TLR5对Treg的调节功能主要以激活为主。
3.4 TLR7对Treg细胞的调节
TLR7在抗病毒防御[47]和几种系统性红斑狼疮小鼠模型自身免疫中起着重要作用[48-49]。TLR7激活可能破坏由Treg介导的外周耐受。有研究表明TLR7通过促进APC分泌IL-6的方式使得应答性T细胞对Treg产生抗性[50]。Hackl等发现在小鼠DC存在的情况下,TLR7配体咪喹莫特类似物S-27609、CL-076和R848可以活化DC上的TLR7,减少分化的Treg数目,这一过程依赖于DC细胞分泌的IL-6,它可能诱导Treg分化成促炎性辅助性T细胞[51]。因此通过干扰TLR7激活或阻断下游效应细胞因子 (如IL-6) 来逆转Foxp3下调可能是治疗SLE的有效策略。Van等发现在OVA诱导的过敏性哮喘小鼠模型中,TLR7激动剂R848能够靶向肺部Treg,使其数目增加,减缓哮喘症状,TGF-β在这一过程发挥主要作用[52]。上述研究提示TLR7配体如果直接作用于Treg上的TLR7可能增强其功能,但如果作用于其他免疫细胞则可能抑制Treg的功能。
3.5 TLR8对Treg细胞的调节
近来,Peng等发现人类Treg表达高水平的TLR8。Poly-G10寡聚核苷酸和类似的TLR8配体ssRNA40、ssRNA33与阳离子脂质的复合物都能直接逆转Treg的抑制功能,促进初始CD4+T细胞的增殖。应用siRNA技术敲除TLR8、MyD88和白介素-1受体相关激酶4 (Interleukin-1 receptor-associated kinase 4,IRAK4) 后,Treg的免疫抑制功能不再被Poly-G10逆转,该实验进一步表明TLR8对于Treg抑制功能的逆转直接依赖于上述效应分子。在T、B细胞都缺陷的Rag1-/-小鼠肿瘤模型中,转输TLR8配体刺激的Treg实验发现TLR8介导的Treg抑制功能的逆转对于抗肿瘤免疫应答有很大的影响[22],提示通过TLR8信号通路抑制Treg,改变其和效应T细胞之间的功能平衡,可能有助于增强针对肿瘤的免疫治疗效果。最近研究表明,人Treg细胞和肿瘤细胞中TLR8信号的激活可以预防细胞衰老[53-54]。不仅如此,TLR8信号对Treg的能量代谢也有调节作用,它能选择性地抑制人Tregs细胞中葡萄糖摄取和糖酵解过程,逆转Treg的抑制作用。在黑色素瘤过继转移T细胞治疗模型中,TLR8信号介导的人Treg细胞内葡萄糖代谢和功能的重编程可以增强体内的抗肿瘤免疫力[55-57]。这些结果均表明TLR8介导的信号通路主要抑制Treg的功能。
3.6 TLR9对Treg细胞的调节
大鼠T细胞与TLR9配体预培养实验证实相比于CD4+CD25+T (Treg) 细胞,CD4+CD25-T(Tconv) 细胞对TLR9配体更敏感。并且经CPG-ODN2006处理的Tconv对Treg细胞的抑制作用产生抵抗,从而间接消除了Treg细胞的抑制活力[58]。在小鼠模型中,TLR9与自身免疫病SLE密切相关,TLR9缺失导致SLE病情加重,这主要与Treg抑制能力受损、效应T细胞活化有关[59]。Hall等发现固有层树突状细胞 (Lamina propria dendritic cell,LpDC) 中TLR9激活后通过抑制Treg细胞来促进肠道炎症。肠道菌群DNA激活TLR9可以破坏肠道稳态,促进Th1和Th17反应,同时抑制CD4+Foxp3-T细胞向CD4+Foxp3+T细胞的转化并完全激活LpDC[60],该结果可以作为治疗肠炎的新型疗法,并且对于口服疫苗的发展也提供了启示[61]。通过共生DNA或合成的TLR9激动剂抑制Treg细胞转化可能改善通过口服途径递送的疫苗的免疫原性。这提示TLR9对Treg的作用受其他免疫细胞和免疫微环境的影响。
4 热休克蛋白直接调节Treg的增殖和抑制功能
热休克蛋白家族通过TLRs信号通路对于Treg的功能有着显著的影响,除了HSP60和HSP70家族成员[62]外,HSP90家族成员gp96(Glycoprotein 96),即GRP94 (Glucose-regulated protein 94)[63]也能够直接增强Treg的功能。首先,内源gp96是Treg谱系维持和抑制功能所必需的分子,gp96缺失会导致Treg谱系不稳定性和体内抑制功能降低。在gp96缺失条件下,Treg无法维持Foxp3的表达水平,导致产生IFN-γ和IL-17的T细胞数量增加[64]。其次,研究表明gp96能够通过TLRs信号通路影响Treg的功能,Dai等发现将转基因小鼠细胞表面的gp96表达量提高,赋予了其对LPS的高反应性,并通过TLR4增强了Treg的抑制功能[65]。更重要的是,gp96作为分子伴侣蛋白,在天然免疫和获得性免疫中都发挥着重要的作用。Brent等发现gp96能够选择性地结合细胞表面的TLR2和TLR4[66]。笔者发现用低剂量 (20 μg) 的gp96免疫小鼠能同时激活效应CD4+T细胞、CD8+T细胞和Treg,但整体以活化效应T细胞为主。随着免疫剂量的提高,gp96介导的免疫活化逐渐向免疫抑制方向转化,小鼠体内IL-10与TNF-α比值逐渐升高。高剂量(100–200 μg) 的gp96免疫主要以活化Treg细胞为主,Treg的活化最终抵消了gp96诱导的效应T细胞活性的增强。进一步发现gp96通过与Treg细胞表面TLR2和TLR4作用活化NF-κB通路,诱导Foxp3以及IL-10、TGF-β1表达[67-69]。同时,Binder等研究发现高剂量的gp96通过上调CD91+pDC (Plasmacytoid dendritic cells) 细胞的Neuropilin-1 (Nrp-1),促进pDC与Treg细胞相互作用的稳定性,进而促进Treg细胞的活化[70](图2)。

图2 高剂量gp96免疫调节Treg的功能Fig.2 High-dose gp96 immunization regulates Treg function.High-dose gp6 activates Treg by two aspects.On the one hand,CD91 on the surface of plasma DC cells binds to gp96 to upregulate neuropilin-1 and stabilize the interaction between pDC and Treg.On the other hand,gp96 interacts with TLR2/ 4 to activate the NF-κB signaling pathway.Thus,high dose gp96 enhances immunosuppressive functions of Treg and can treat autoimmune diseases.
在上述研究基础上,笔者通过小鼠模型进一步验证高剂量gp96免疫活化Treg,可有效治疗因免疫过度活化引发的肝衰竭和自身免疫系统引发的Ⅰ型糖尿病[71],这些研究提示高剂量gp96在治疗自身免疫性疾病中具有广泛的应用潜力。
5 总结
大量研究表明TLRs激动剂可以通过直接或间接的方式调节Treg增殖和免疫抑制功能 (图3)。就直接调节作用而言,有研究表明TLR2、TLR4和TLR5等分子活化后可以直接增强Treg细胞的免疫抑制活性,另有研究则表明TLR2、TLR8和TLR9可以消除或逆转CD4+CD25+Treg细胞的免疫抑制功能,而TLR2、TLR4分子对Tregs功能的具体影响至今仍未有明确定论,需要进一步深入的研究。同时,除了影响Treg的功能外,TLR2、TLR4、TLR7和TLR9在配体的刺激下可促进CD4+CD25+Treg细胞的增殖,提高CD4+CD25+Treg数量。造成以上研究结果不一致的原因可能有:首先不同TLRs在Treg表面或细胞内的表达水平存在很大差异,细胞中其他分子如SOCS1、IRAK1、NFκB、TRAF3也会参与调节TLR下游通路[72],因此对这些TLRs定量分析、细胞内其他调节分子的检测将有助于分析激动剂对TLRs介导下游通路激活的强度和对Treg功能的影响;其次,Treg表型的不稳定性以及效应Tregs不同的亚型[73]都可能影响TLRs下游通路对其抑制功能的作用,因此有必要深入研究TLRs配体或激动剂对不同表型和不同亚型的Tregs的影响。就间接调节作用而言,APC上的TLR2、TLR7和TLR9活化后通过分泌促炎性细胞因子增强效应细胞的功能,进而间接减弱Treg的抑制功能,这说明应当在Tregs所处的具体免疫环境下了解TLR介导的通路对其功能的影响。此外,Foxp3作为Treg标志性转录因子对Treg的功能和分化具有重要影响,但是TLRs活化对Foxp3表达的调节机制还有待深入研究。热休克蛋白家族多个成员可以通过TLR2、TLR4等作用进而增强Treg的功能,可作为潜在新型TLRs激动剂治疗自身免疫性疾病,由于多个以热休克蛋白为基础的药物或疫苗已经用于临床治疗,其安全性有保障,因此具有良好的应用前景。综上所述,TLRs配体或激动剂在调节Treg的功能、增殖方面发挥重要作用,鉴于Treg在维持机体免疫稳态和引发免疫耐受中均发挥关键作用,因此深入探究不同TLRs分子活化和下游通路对于Treg免疫功能的影响及作用机制,开发新型TLRs激动剂用于自身免疫病、抗感染免疫和抗肿瘤免疫研究,不仅具有理论价值,而且具有实际应用潜力。
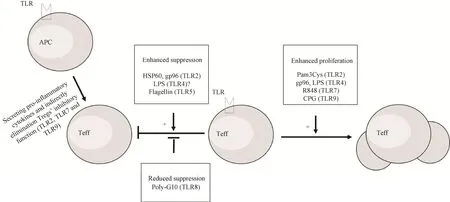
图3 TLR活化对Treg增殖与功能的调节Fig.3 Regulation of proliferation and function of Treg by TLR activation.The activation of TLR2,TLR4 and TLR5 on the surface of Treg directly enhances its immunosuppressive function,while TLR8 ligand reverses its immunosuppressive function.TLR not only affects Treg function,but also its proliferation.Activation of TLR2,TLR4,TLR7 and TLR9 directly promotes Treg proliferation.
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07 06:21:30
材料与冶金学报(2022年2期)2022-08-10 09:15:38
汽车维修与保养(2021年8期)2021-02-16 00:28:20
作文成功之路·小学版(2020年6期)2020-07-27 01:48:28
材料科学与工程学报(2016年4期)2017-01-15 13:35:48
工业设计(2016年4期)2016-05-04 04:00:15
合成化学(2015年4期)2016-01-17 09:01:11
无机化学学报(2014年10期)2014-02-28 17:33:13
无机化学学报(2014年6期)2014-02-28 17:32:06
无机化学学报(2014年5期)2014-02-28 17:31:42
