
5-氨基乙酰丙酸在18种园林植物上的应用效果
2020-03-23 10:35周志凯安玉艳汪良驹
淮阴工学院学报 2020年1期
李 旭,徐 丽, ,周志凯,安玉艳, 汪良驹*
(1. 泗洪县园林绿化管理所, 江苏 宿迁 223900;2.南京农业大学 园艺学院,南京 210095,3.淮阴工学院,江苏 淮安 223003)
园林植物生长离不开肥料施用,科学施肥是园林植物健康生长的基础。在常规生产管理过程中,园林植物施肥均以有机肥为主,同时配合一定量的化学肥料作为追肥,以便在改良土壤的同时,能够促进植物生长[1]。在现代园林生产过程中,由于可以促进植物对肥料的吸收,提高肥料利用效率,还可以肥水一体,节省人工成本,便于生产管理,液体肥料的施用越来越受重视。
5-氨基乙酰丙酸(ALA)是一种存在于动植物和微生物体内的天然氨基酸,但它不参与蛋白质合成,而是作为叶绿素、血红素等卟啉化合物生物合成的关键前体,参与植物的光合作用、呼吸作用、蒸腾作用、营养代谢和次生物质合成等,特别是能够提高植物的抗逆性[2-4]。现已证明,ALA可以提高植物抵抗高温、低温、干旱、水涝、盐渍、强光或弱光等胁迫能力,在农林生产上具有广阔用途。在观赏植物上,ALA可以提高红掌叶片光化学效率,增强抗逆性[5]。ALA或者含有ALA的肥料可以促进月季叶片的叶绿素含量提高,促进植株生长,增加开花数量,并且改善花的颜色[6]。用2~20 mg·L-1ALA处理可以显著改善红香芋试管苗耐冷性[7]。叶面喷施300 mg·L-1ALA溶液可以显著提高红掌叶片的叶绿素含量,促进干物质的积累,加快新叶抽生速度,有利于茎的横向加粗生长和伸长生长[8]。在牡丹现蕾期每10 d喷施一次ALA溶液,可以促进花芽形成和花朵开放[9]。用ALA处理菊花植株,可以延长鲜切花采后寿命,提高其观赏价值[10]。叶面喷施50 mg·L-1ALA能够提高切花菊抗寒性,促进光合作用和植株生长,促进低温下切花菊开放,缩短成花时间[11]。在温室百合上,喷施ALA溶液,可以促进植株生长,提高叶片叶绿素含量,有效延长瓶插切花寿命。此外,ALA处理还促进地下茎籽球的形成[12]。ALA处理可以提高盐胁迫下栀子植株抗氧化酶活性,促进脯氨酸积累,减少丙二醛含量和相对电导率,提高植物耐盐性[13]。最近,陈慧等[14]、王冬云等[15]报道了ALA提高人为高温或低温条件下几种园林植物的耐受性,证明这种新型植物生长调节剂可以增强园林植物抗逆性。但是,我国园林植物种类繁多,即使在某一特定地区,园林植物也有数十上百种。迄今尚未见到ALA在多种园林植物生产中应用报道。本试验以泗洪县世纪公园内主栽的18种园林植物为材料,通过根系浇灌或者叶面喷施的方式,将ALA液体供给植物,25 d后,观察植株叶片形态、测定叶片的叶绿素含量以及叶绿素快速荧光和820 nm反射荧光特性,旨在探讨绝大多数园林植物对ALA溶液的反应,从而为其在园林植物生产中应用提供支撑。
1 材料与方法
1.1 材料
本试验所用材料全部定植于宿迁市泗洪县世纪公园,植物种类有杜鹃(Rhododendronsimsii)、桂花(Osmanthusfragrans)、海桐(Pittosporumtobira)、樱花(Prunusserrulata)、银杏(Ginkgobiloba)、乌桕(Sapiumsebiferum)、栾树(Koelrenteriapaniculata)、香樟(Cinnamomumcamphora)、金森女贞(Ligustrumjaponicum)、杮树(Diosporuskaki)、南天竹(Nandinadomestica)、金边阔叶麦冬(Liriopemuscaricv.Variegata)、蔷薇(Rosamultiflora)、金山绣线菊(Spiraeajaponica)、绣线菊(Spiraeasalicifolia)、红叶石楠(Photiniafraseri)、大花金鸡菊(Coreopsisgrandiflora)等。其中,香樟树分为两类,一类是早春遇到倒春寒冻害严重受伤而重截枝重新萌发的树,一类是冻害不重基本正常生长的树。所用5-氨基乙酰丙酸(ALA)药剂由南京禾稼春生物科技有限公司提供,稀释倍数2000(折合ALA浓度50 mg·L-1)。
1.2 试验设计
试验于2017年4月21日进行。对于红叶石楠、大花金鸡菊、蔷薇、绣线菊、南天竹、金边阔叶麦冬、金森女贞、海桐、杜鹃等草本或灌木来说,叶面喷施ALA溶液,直至叶面液滴滴下为止;对于杮树、香樟、银杏、乌桕、栾树、樱花等大型乔木来说,每株浇灌5 kg ALA溶液。每处理乔木5~10株树,灌木或草本20 m2以上,设清水为对照,其它正常管理。25 d后,随机选取处理与对照新梢中部叶片,测量大小、厚度和叶绿素相对含量(SPAD),测定叶绿素快速荧光曲线和820 nm光反射荧光曲线,分析对照与处理之间光合系统I和光合系统II活性。通过外观形态和光合光化学效应,判定ALA在园林植物上应用效应。
1.3 测定项目与方法
叶片长度、宽度用卷尺测定,厚度用游标卡尺测定,叶绿素相对含量用SPAD502 Plus叶绿素含量测定仪测定。每个数值至少重复10次以上,取平均值,然后用处理比对照,得到相对数值,并以此做分析。当比值大于1,说明ALA有促进效应;反之,则有抑制效应。
叶绿素快速荧光和820 nm光反射荧光用M-PEA多功能叶绿素荧光仪测定[15]。所有叶片在黑暗下暗适应30 min以上,用M-PEA测定瞬时荧光值和820 nm反射荧光值,绘制OJIP曲线和MRo/MR曲线。根据表1提供的方法计算荧光参数值。每个处理至少测定10次,取平均值,然后用处理比对照,得到相对数值,做方差分析、Duncan氏测验和聚类分析(IBM SPSS20)。当P< 0.05时,认为差异达到显著水平;当P< 0.01时,认为差异达到极显著水平。

表1 OJIP荧光参数的含义及其计算方法
2 结果与分析
2.1 ALA处理对园林植物叶片形态与叶绿素相对含量的影响
从表2可知,ALA处理后25 d,绝大多数植物叶片变大、变厚,而且叶色变浓,叶绿素含量增加。从18种园林植物的总平均值来看,叶长和叶宽增加幅度为13%~16%,叶厚增加约5%,叶绿素相对含量增加20%。方差分析结果显示,ALA处理效应达到极显著水平。
从单一物种来说,叶长增幅达到P= 0.01显著水平的有杜鹃、桂花、海桐、栾树、受冻害香樟、金森女贞、蔷薇、金山绣线菊、绣线菊和大花金鸡菊,其增幅均大于13%;差异达P= 0.05水平的有乌桕、南天竹和红叶石楠,增幅为9%以上;其它植物如樱花、柿、未受冻害的香樟和金边阔叶麦冬叶长与对照间没有明显变化(P> 0.05)。从叶宽来看,杜鹃、桂花、海桐、银杏(以半径计算)、乌桕、香樟(受冻害后)、金森女贞、金边阔叶麦冬、金山绣线菊、绣线菊、红叶石楠和大花金鸡菊等增加量达12%以上,差异达到P= 0.05水平。蔷薇、樱花和南天竹的叶宽增量为8%,差异达到P= 0.05水平。香樟(未受冻的)、栾树和杮树叶片宽度与对照间没有明显差异。其中,柿叶片还有一定下降,只是没有达到显著水平(P> 0.05)。从叶片厚度上看,杜鹃、桂花、樱花、金森女贞、金边阔叶麦冬和蔷薇的增量均达到P= 0.01显著水平,幅度超过13%,而受冻害的香樟增幅为11%,达到P= 0.05水平。大花金鸡菊叶厚明显下降(P< 0.05)。海桐、银杏、乌桕、栾树、香樟(未受冻害)、柿、金山绣线菊、绣线菊、南天竹和红叶石楠叶片厚度与对照没有显著差异。从叶片SPAD上看,杜鹃、桂花、海桐、银杏、栾树、香樟(包括受冻和未受冻的)、金森女贞、金边阔叶麦冬和南天竹的叶绿素相对含量增加11%以上(P< 0.01);樱花、绣线菊增加9%以上(P< 0.05);只有乌桕、柿树、蔷薇和金山绣线菊叶片SPAD没有明显变化,而红叶石楠SPAD极显著降低(P< 0.01),说明叶片叶绿素含量因ALA处理而下降。这可能与其叶片变红以及花青素含量上升有关。
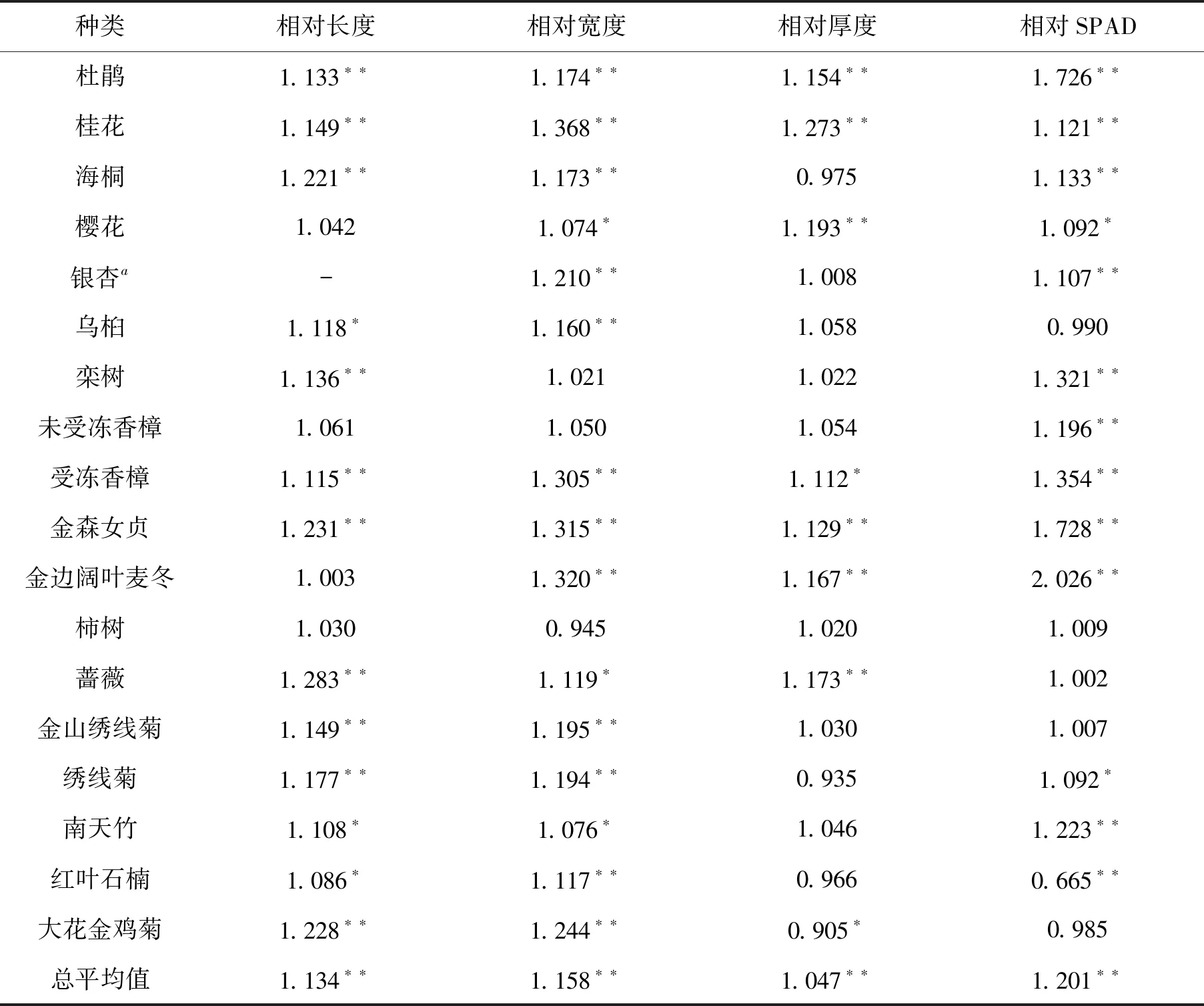
表2 ALA处理后园林植物叶片形态指标和叶绿素相对含量(与对照相对比)
注:a表示银杏叶片宽度指其半径;每种植物均为10张叶片平均值,经双向方差分析和显著性测验;*和**分别表示
P=0.05和P=0.01水平上差异显著;总平均值为F测验结果,**表示F > F0.01,下同。
2.2 ALA对园林植物叶绿素荧光参数的影响
2.2.1基本荧光参数
表3显示,ALA处理25 d后,尽管18种园林植物经ALA处理后叶片叶绿素初始荧光(Fo)总平均值极显著上升(F > F0.01),但绝大数物种Fo并没有明显变化,只有金边阔叶麦冬和南天竹Fo极显著上升,而红叶石楠极显著下降。这3种植物叶片都含有较多的花青苷,对ALA处理的响应与其它物种有所不同。正常条件下,绿叶植物叶片Fo几乎不受ALA处理影响。
从最大荧光值(Fm)相对值看,18种材料Fm总平均值上升11.8% (F > F0.01)。其中,杜鹃、栾树、金边阔叶麦冬和南天竹Fm增加20%以上(P< 0.01),海桐、受冻的香樟、金森女贞、柿和金山绣线菊增加7%以上(P< 0.05),而红叶石楠则极显著低于对照(P< 0.01)。这说明,红叶植物的Fm对ALA处理的响应可能具有特异性。
从可变荧光(Fv)相对值看,18种材料Fv总平均值上升13.5% (F > F0.01)。其中,杜鹃、海桐、栾树、受冻香樟、金森女贞、金边阔叶麦冬、杮、金山绣线菊和南天竹等经ALA处理后Fv显著或极显著上升,而红叶石楠则极显著下降。红叶石楠Fv下降与其Fm下降有关。
2.2.2 PSII反应中心最大光化学效率
表3显示,18种园林植物经ALA处理后叶片PSII最大光化学效率(ΦPo)整体上变化很小,除南天竹ΦPo小于1外,其它的都≥1,总平均值增加1.5%,说明ΦPo是一个比较稳定的叶绿素荧光指标,但ALA处理仍然可以提高叶片PSII最大光化学效应。其中,杜鹃、栾树和金边阔叶麦冬ΦPo上升达到P= 0.01差异水平,受冻香樟叶片ΦPo达到P= 0.05差异水平。其它物种ΦPo变化不大(P> 0.05)。
2.2.3 PSII反应中心供体侧活性
Wk代表PSII反应中心供体侧放氧复合体受抑程度。Wk升高,说明受抑程度越大。从表3看出,绝大多数绿色植物叶片经ALA处理后Wk< 1,说明PSII反应中心供体侧活性受到促进(F > F0.01)。其中,杜鹃、银杏、栾树、香樟(无论是否受冻)和金边阔叶麦冬Wk下降到P= 0.01水平,蔷薇下降达到P= 0.05水平。但是,南天竹、红叶石楠和大花金鸡菊三种植物叶片Wk极显著上升(P< 0.01)。ALA处理为何导致其Wk上升值得深入研究。
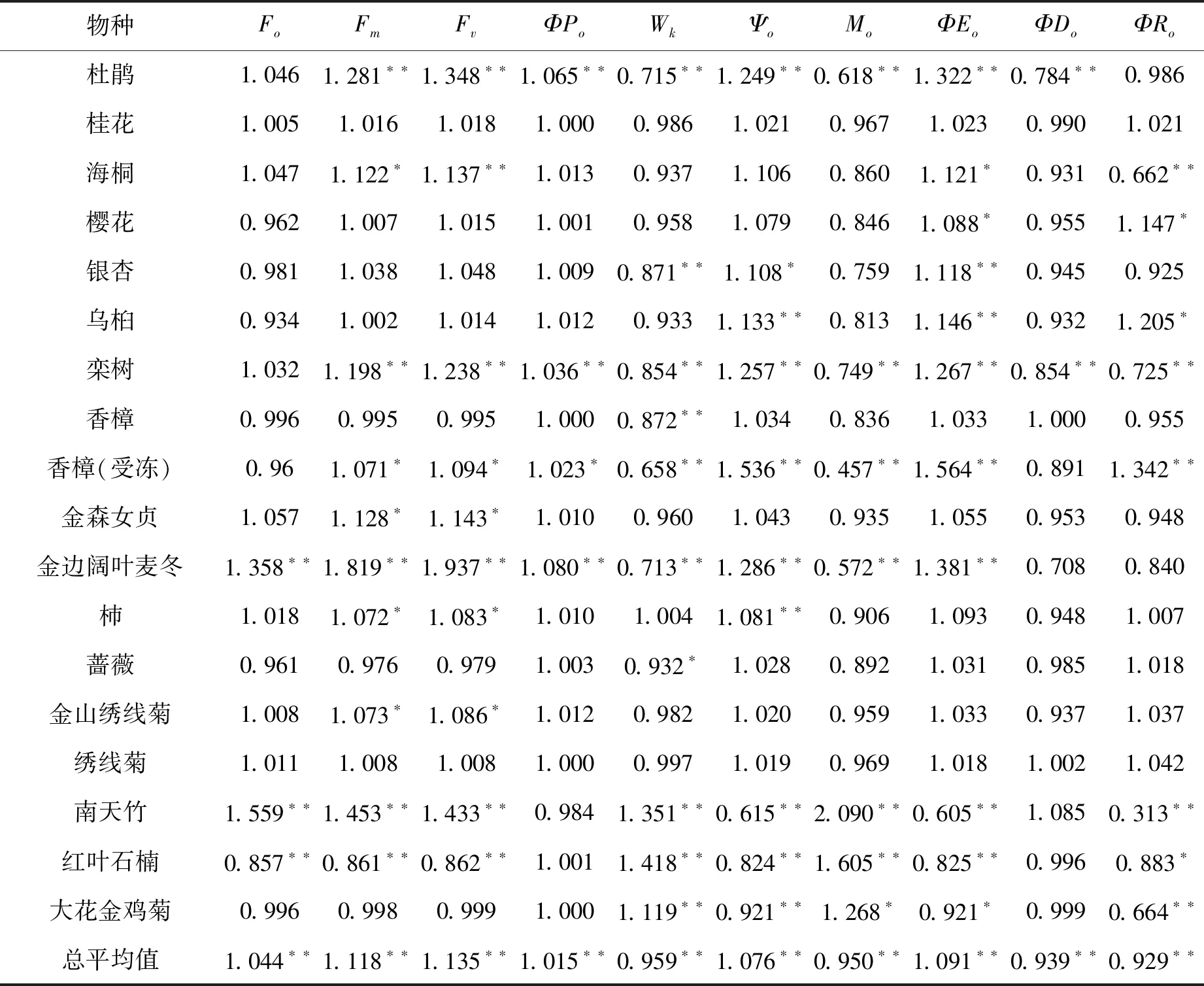
表3 ALA处理后不同物种叶片基础荧光参数和光合系统反应中心活性荧光参数相对值 (与对照比较)
2.2.4 PSII反应中心受体侧活性
Ψo代表着PSII反应中心受体侧捕获的激子将电子传递到电子传递链中QA以远其它电子受体的概率,而Mo表示PSII反应中心关闭的最大速率。Ψo越高,PSII反应中心活性越大;Mo越小,反应中心开放时间越久。表3显示,ALA处理后,杜鹃、乌桕、栾树、受冻香樟、金边阔叶麦冬和杮等Ψo极显著高于对照(P< 0.01),银杏Ψo显著高于对照(P< 0.05),而南天竹、红叶石楠和大花金鸡菊Ψo则极显著低于对照。18种材料Ψo总平均值比对照高7.6%(F > F0.01)。
从Mo看,ALA处理后,南天竹、红叶石楠和大花金鸡菊3种植物Mo极显著高于对照,其它绝大多数物种Mo下降或者变化不大。18种材料Mo总平均值比对照低5.0%,说明ALA对园林植物叶片PSII反应中心活性开放的促进效应具有普遍性。
2.2.5量子产额
ΦEo是用于电子传递的量子产额,ΦDo是热耗散的量子产额。ΦEo越高,光合能量转换效率越高;ΦDo越高,光合能量转换效率越低。从表3可以看出,除了南天竹、红叶石楠叶片ΦEo显著或极显著低于对照外,其它十多种植物叶片ΦEo均因ALA处理而上升,其中杜鹃、海桐、樱花、银杏、乌桕、栾树、受冻香樟、金边阔叶麦冬ΦEo显著或极显著高于对照。18种材料ΦEo比对照高9.1%(F > F0.01)。
另一方面,绝大多数物种叶片ΦDo因ALA处理而下降。南天竹ΦDo略高于对照,但P>0.05,未达到差异显著水平。红叶石楠、大花金鸡菊、绣线菊、桂花和正常生长香樟的ΦDo与对照间没有明显差异。18种材料ΦDo比对照低6.1%(F > F0.01),说明ALA处理后,植物叶片通过热耗散方式消耗的能量减少,更多的光能通过光合电子传递链转换为生物化学能。
ΦRo是用于还原PSI受体侧末端电子受体的量子产额,代表着PSI光化学效率。从表3可以看出,ALA处理后不同物种叶片ΦRo出现不同变化,但整体上是下降的。18种材料经ALA处理后叶片ΦRo比对照低7.1%(F > F0.01)。樱花、乌桕、受冻害香樟显著或极显著高于对照,而海桐、栾树、南天竹、红叶石楠和大花金鸡菊显著或极显著低于对照。ALA处理后不同物种存在着不同的反应,值得进一步讨论。
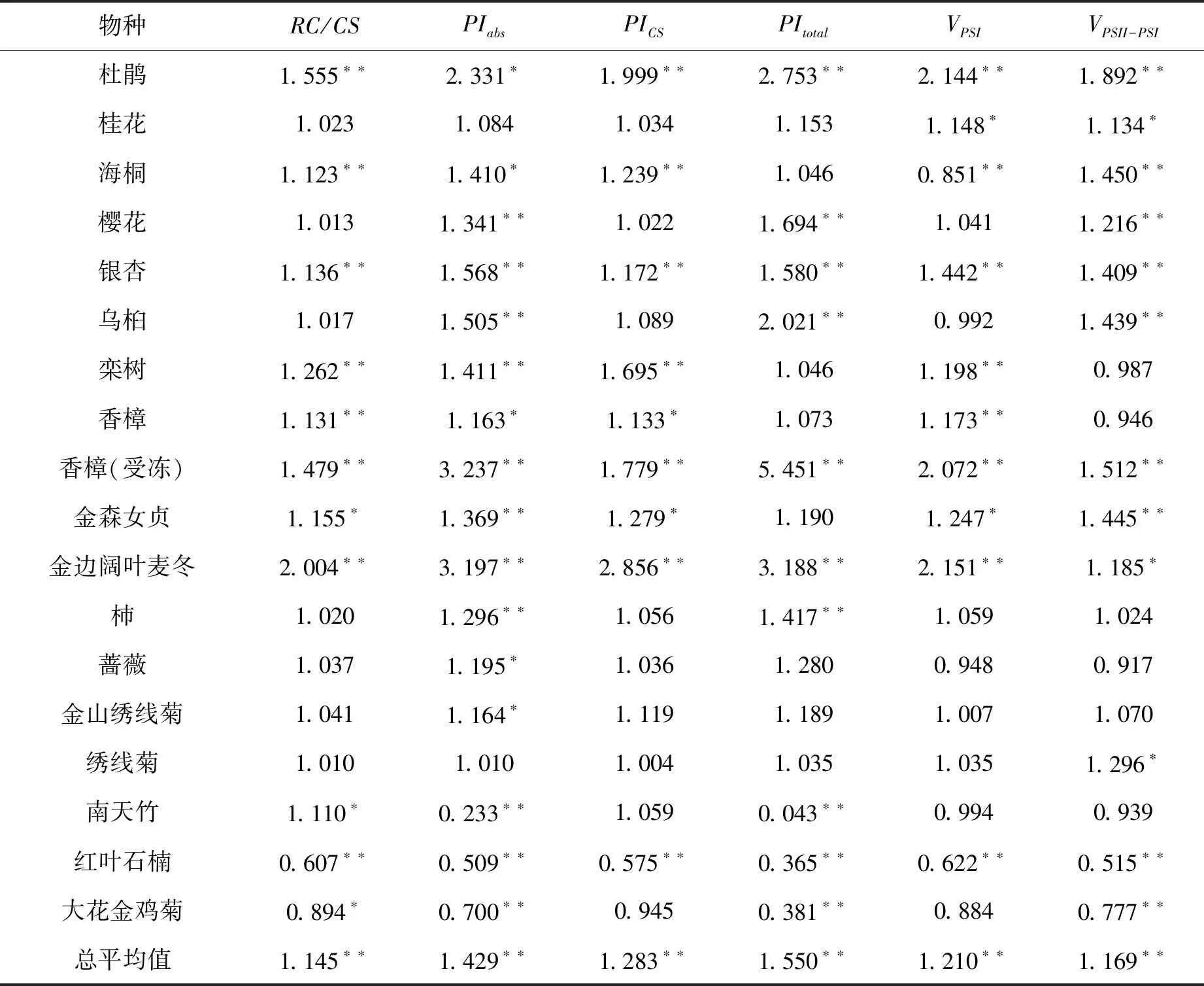
表4 ALA处理后有活性反应中心密度、光合性能指数和PSI氧还速率相对值 (与对照比较)
2.2.6单位面积有活性反应中心密度(RC/CS)和光合性能指数(PIabs、PIcs和PItotal)
表4显示了经ALA处理25d后不同园林植物叶片有活性的PSII反应中心密度(RC/CS)以及光合性能指数PI,包括以吸收为基础的性能指数PIabs、以面积为基础的性能指数PIcs和包括PSI和PSII在内的总的性能指数PItotal。从表4可知,ALA处理后,杜鹃、海桐、银杏、栾树、香樟(无论受冻与否)和金边阔叶麦冬的RC/CS均极显著上升,金森女贞和南天竹RC/CS显著上升,但红叶石楠和大花金鸡菊RC/CS却因ALA处理而显著或极显著下降。18种材料RC/CS总平均值比对照高出14.5%
(F > F0.01),说明ALA处理使得植物叶片PSII有活性反应中心的密度极显著上升。
从PIabs上看,ALA处理的18种园林植物叶片总平均值极显著上升(F > F0.01),其中杜鹃、樱花、银杏、乌桕、栾树、受冻香樟、金森女贞、金边阔叶麦冬和杮极显著高于对照(P< 0.01),海桐、未受冻香樟、蔷薇和金山绣线菊显著高于对照(P< 0.05)。但是,南天竹、红叶石楠和大花金鸡菊PIabs极显著低于对照(P< 0.01),暗示着这些物种光合性能指数对ALA的响应不同于一般的绿叶植物。
从PIcs上看,杜鹃、海桐、银杏、栾树、受冻香樟和金边阔叶麦冬经ALA处理后均极显著高于对照,未受冻香樟和金森女贞PIcs显著高于对照。其中,金边阔叶麦冬比对照高出186%,杜鹃高出99.9%,受冻害的香樟达78%,栾树达69.5%。但是,红叶石楠PIcs极显著低于对照。这也与物种有关。18种材料PIcs总平均值比对照高28.3%(表4)。
从PItotal上看,除了南天竹、红叶石楠和大花金鸡菊PItotal极显著低于对照外,其它物种PItotal均因ALA处理而有不同程度提高。18种材料PItotal总平均值比对照高55%(表4)。其中,杜鹃、樱花、银杏、乌桕、受冻害香樟和金边阔叶麦冬的差异达到P= 0.01显著水平。PItotal值大幅度增加,暗示着ALA处理对大多数园林植物叶片整个光合能力的促进效应。
2.2.7 PSI反应中心氧化活性(VPSI)和还原活性(VPSII-PSI)
从表4可以看出,ALA处理后,多数物种PSI反应中心氧化活性(VPSI)和还原活性(VPSII-PSI)都有明显增加。18种材料VPSI总平均值比对照高出21%,VPSII-PSI比对照高出16.9%(F > F0.01)。就VPSI而言,杜鹃、桂花、银杏、栾树、香樟(无论受冻与否)、金森女贞和金边阔叶麦冬经ALA处理后都显著或极显著高于对照,而海桐和红叶石楠VPSI则显著低于对照。类似地,杜鹃、桂花、海桐、樱花、银杏、受冻香樟、金森女贞、金边阔叶麦冬和绣线菊VPSII-PSI显著或极显著高于对照,而红叶石楠和大花金鸡菊VPSII-PSI极显著低于对照。
2.3 ALA处理后植物叶片形态学变量与叶绿素荧光参数变量间相关性分析
由Pearson分析可以看出(见表5),叶长相对变化与这些荧光参数间的相关性没有达到显著水平(P> 0.05),但叶宽却与VPSII-PSI呈显著正相关关系,而叶厚与δRo、VPSI和VPSII-PSI等呈显著正相关关系(P< 0.05),与PItotal呈极显著相关关系(P< 0.01),说明整个光合系统(包括PSI和PSII)的光合性能指数与大多数园林植物的叶片增厚存在着正相关关系,而与叶片增长、增宽关系不密切。
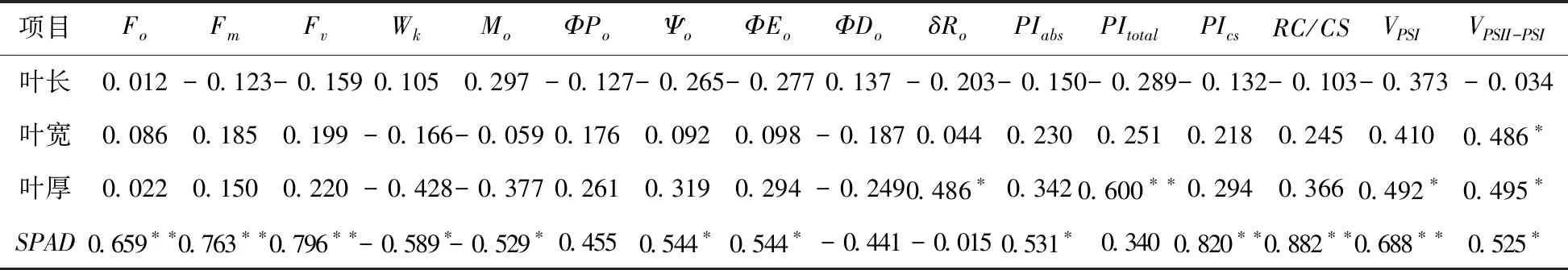
表5 园林植物叶片外观形态与叶绿素荧光参数间的相关分析
注:*和**分别表示在P=0.05和P=0.01水平上相关性显著。
与此不同的是,叶片叶绿素相对含量(SPAD)变化与多数叶绿素荧光参数变化关系密切。其中,呈显著或极显著正相关关系的有Fo、Fm、Fv、Ψo、ΦEo、PIabs、PICS、RC/CS、VPSI和VPSII-PSI,呈显著负相关关系的有Wk和Mo等参数。由于前组参数与光化学能量转化有关,后组参数与光合系统活性受抑有关,说明ALA诱导的叶片叶绿素含量增加一方面有助于提高叶片光合性能,另一方面可以减少光合抑制程度。
2.4 18种园林植物对ALA处理响应的模糊聚类
上文显示,不同园林植物的叶片形态和叶绿素荧光特性对ALA处理有着不同反应。据此,对这些植物做聚类分析(见图1)。如果从距离25处区分,18种植物可以分为两类,即受冻香樟为一类,其它17种物种为另一类。这说明,ALA对冻害后植物恢复具有特殊效应。对于后17种植物来说,如果从距离10处区分,杜鹃和金边阔叶麦冬为一类,其余15种植物为另一类。前2种植物对ALA处理非常敏感,后15种相对不敏感。对后15种植物来说,如果从距离7处区分,红叶石楠、大花金鸡菊和南天竹为一类,其他12种植物为另一类。至此,根据不同植物对ALA处理的响应特性可以将18种园林植物分为4类。第一类为受冻香樟,它的叶片生长和叶绿素荧光特性对ALA响应非常敏感。第二类有杜鹃和金边阔叶麦冬。它们对ALA的响应也非常敏感。第三类有红叶石楠、大花金鸡菊和南天竹。它们以红叶为主,对ALA处理的响应不同于其它绿叶植物。第四类为大多数绿叶植物,包括蔷薇、金山绣线菊、绣线菊、未受冻的香樟、桂花、樱花、柿、栾树、金森女贞、银杏、乌桕等。
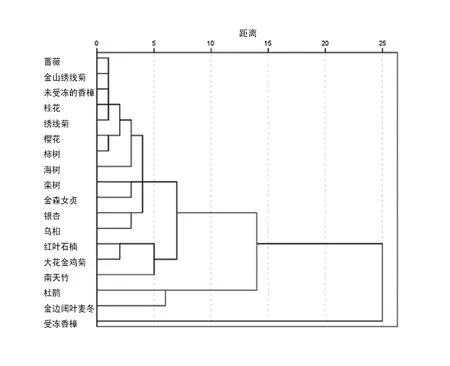
图1 18种园林植物的模糊聚类分析
3 讨论与结论
5-氨基乙酰丙酸(ALA)是一种广泛存在于动植物和微生物体内的天然氨基酸,具有独特的植物生长发育调节功能。日本学者Hotta等[16]首先提出外源ALA的生理调节作用。国内学者在甜瓜[17]、小白菜[17]、萝卜[19]、西瓜[20]、草莓[21-22]、黄瓜[23]、番茄[24]、菠菜[25]、水稻[26]、枣[27]、小麦[28]、芸豆[29]、大豆[30]、梨[31]、无花果[32]和苹果[33-34]等多种作物上先后报道了ALA的应用效应。但是,ALA在园林植物中应用研究的报道尚不算多。即使有,也是以某些单一植物为材料开展的专项研究[5-13]。系统研究ALA在园林植物上的应用尚鲜见报道。
本文以泗洪县世纪公园内种植的18种园林植物为材料,研究了ALA对其叶片生长和叶绿素荧光特性的影响,发现处理25 d后,绝大多数植物叶长、叶宽、叶厚以及叶绿素含量均有显著提高(表2),表明ALA具有促进植物生长效应。无论是叶面喷施还是根系浇灌,ALA对光合性能的促进效应是明显的。主要表现在:①对PSII反应中心供体侧放氧复合体活性的促进效应,即经ALA处理的叶片Wk明显下降(表3);②对PSII反应中心最大光化学效率的促进效应,即经ALA处理叶片ΦPo明显增加;③对PSII反应中心开放程度的促进效应,即经ALA处理后,Mo明显下降;④对PSII反应中心捕获的激子将电子传递到QA以远的其它电子受体的概率Ψo明显增加;⑤提高PSII有活性反应中心的密度(RC/CS)(表4);⑥降低叶片捕获的光能以热耗散的形式消耗的量子产额ΦDo;⑦提高PSI反应中心氧化活性VPSI;⑧提高PSII传递电子到PSI反应中心的活性VPSII-PSI;⑨提高以吸收为基础的光合性能指数PIabs、以面积为基础的光合性能指数PIcs和包括PSI和PSII在内的整个光合系统的性能指数PItotal。所有这些结果,均为ALA在园林植物上的应用提供了理论依据。但是,ALA处理降低部分植物PSI反应中心的光化学效率ΦRo。这可能与ALA的作用机理有关。现有理论认为,ALA之所以可以提高植物叶片光合能力,与其促进PSI附近的抗氧化酶活性有关[35]。这些抗氧化酶,包括SOD、POD、CAT和APX等,可以接受PSII传递过来的电子。虽然这样会降低PSI光化学效率ΦRo,但因为减少了PSI的光抑制,有利于提高两个光合系统之间的电子传递(VPSII-PSI),所以整个叶片光合能力(PItotal)因此能显著提高。因而,PSI光化学效率ΦRo下降是光抑制下降的反应,而不是光合速率下降的标志。在本试验所在的泗洪县世纪公园内,喷施过ALA的红叶石楠新梢迅速生长,形成大量的鲜红色嫩叶。这大大增加了红叶观赏价值,也说明ALA处理引起的ΦRo下降不是一种不良的反应。
ALA处理既促进植物叶片生长,也能提高光合性能,但并非所有指标之间都存在显著相关关系。比如,ALA处理促进红叶石楠、绣线菊和大花金鸡菊的叶片长度和宽度,但叶片厚度下降,其中大花金鸡菊的差异达到显著水平(表2)。另外,ALA处理的大花金鸡菊株高、花量也高于对照(数据未列出),但是光合性能指数(包括PIabs、PIcs和PItotal)却显著低于对照(表4)。这一点与南天竹和红叶石楠等红叶树种相似。在杮树上,ALA处理后叶长、叶宽、叶厚和叶绿素相对含量均无显著变化,但PItotal却增加了41%,说明叶片光合性能极显著上升。相关分析表明,ALA处理引起的植物叶片叶绿素荧光参数变化与叶片长度变化的相关性较低,与叶宽有一定关系,与叶厚关系更大,与叶片SPAD变化相关性最高(表5)。实际上,植物叶片相对厚度往往与其适应外界环境能力有关。叶片越厚,相对光合效率越高,适应环境能力越强。ALA之所以能够运用于园林植物,与其提高叶片相对厚度,增强光合能力和环境适应性有关。需要注意的是,ALA处理并没有提高红叶石楠叶片叶绿素含量,相反,它显著降低了叶片SPAD(表2),同时叶片红色素含量显著增加(资料未列出)。这与在苹果上的结果相似,外源ALA处理可以促进植物组织花青苷的生物合成[33]。事实上,ALA处理的红叶石楠新梢生长快,红色鲜艳,观赏价值高。
本文首次利用聚类分析法并根据植物对ALA处理的反应来区分植物种类,发现受冻香樟单独成一类。前人早已证明,ALA对逆境条件下植物光合速率的促进效应明显大于适境条件,因而ALA处理叶片的光合午休程度明显低于对照[29]。我们也曾经报道ALA处理可以提高香樟离体叶片耐寒性[14]。本文利用受冻害的香樟和未受冻的香樟作对比,观察到ALA对前者的影响远远大于后者,甚至单独于所有其它试验物种(图1),说明在入冬前或者倒春寒低温冻害来临前,浇灌一定的ALA,可以降低倒春寒冻害程度,提高香樟这种重要园林绿化树种的绿化效果。杜鹃和金边阔叶麦冬对ALA处理的响应成为一类。它们对ALA处理敏感。在试验当年,4~6月份处于长期阴雨天气,7~8月又处于长期高温干旱天气。9月份调查植株生长情况,发现对照植株伤害严重,长势不良,而ALA处理植株生长良好,证明植物抗逆性的确明显提高。另外,红叶石楠、南天竹和大花金鸡菊分成一类。这意味着大花金鸡菊与其它两种红叶植物类似,具有相似的响应机制。但具体响应机制尚不明确。
总之,ALA可以广泛应用于园林植物生产。尽管不同植物间的反应有所差异,但其提高植物抗逆性的效应以及增强园林绿化和美化的效果是明显。一些深层次的机制问题需要进一步研究阐明。
猜你喜欢
花卉(2022年11期)2022-06-09
故事会(2022年10期)2022-05-20
西安交通大学学报(社会科学版)(2021年2期)2021-04-06
知识窗(2019年7期)2019-08-12
北方音乐(2019年6期)2019-07-16
浙江农业科学(2019年4期)2019-04-19
作文成功之路·作文交响乐(2019年10期)2019-01-10
课外生活(小学1-3年级)(2018年1期)2018-02-10
小朋友·聪明学堂(2015年10期)2015-11-17
小小说月刊·下半月(2015年12期)2015-05-14
