
基于SSR技术对黄河三角洲二色补血草遗传多样性研究
2020-03-05 04:04:28吕冬雪张学杰张洛艳樊守金
植物研究 2020年6期
吕冬雪 张学杰 张洛艳 樊守金
(山东师范大学生命科学学院逆境植物重点实验室,济南 250014)
二色补血草(Limonium bicolor(Bag.)Kuntze)是越年生草本植物,隶属白花丹科(Plumbaginaceae)补血草属(Limonium)[1],典型的盐生植物[2],二色补血草分布在黄河流经省区,江苏北部等。它常生长在较低的斜坡、丘陵、平原和沿海地区,生于含盐的钙质土壤和砂地[2]。专家对其研究主要集中在生理抗盐机制、化学成分、栽培技术方面,但对其遗传多样性研究较少。为了探讨黄河三角洲盐渍生境下,二色补血草的遗传多样性水平及其成因,探究二色补血草居群遗传分化的程度与遗传结构的形成机制,为二色补血草种质资源保护提供科学依据。SSR 分子标记是由2~6 个核苷酸组成的串联重复序列,根据两端序列设计特异性引物,通过PCR 技术扩增微卫星DNA 序列,再经过聚丙烯酰胺凝胶电泳和毛细管电泳获得长度的多态性[3]。SSR 分子标记技术具有多种优势,共显性遗传、重复性好、位点丰富。结合荧光标记技术,智能化较高,真正实现大量数据快速收集处理,有效避免人工读带误差,数据准确,对于物种资源收集,保护改良[4]具有重要意义。随着NCBI中EST 数据库的丰富与完善,序列开发SSR 标记在众多植物中得到广泛应用[5]。这样避免了传统标记开发需要构建基因组DNA 文库等繁琐步骤,成为种质资源多样性、遗传作图、物种起源与进化等研究常用方法[6]。
1 材料及实验方法
1.1 材料采集
在黄河三角洲一定地理区域内,调查发现50个二色补血草居群,采用随机取样的方式对50 个地点均匀、分散、有间距的取样,选出12 个自然居群。根据自然居群现有分布规模以“株”为单位随机取样,个体间距10 m,采集新鲜叶片。共202 份样品。在这12 个地点,进行土壤样品采集。在每处耕层垂直挖掘30 cm 的深度,沿着切割表面从底部到顶部取适量土壤样品。每处采集3份。用电导率仪(DDS-307)测量土壤浸出液的电导率(见表1)。
1.2 DNA提取
琼脂糖电泳质量检测(Agarose gel electrophoresis,AE),采用浓度为1%的琼脂糖凝胶电泳,检测基因组DNA 的完整性和降解情况。将其置于凝胶成像系统中观察,DNA 具有清晰的主条带无断裂、拖尾、降解现象,拍照记录。浓度检测使用微量紫外分光光度计NanoDrop2000 测量DNA 提取物的浓度与纯度。
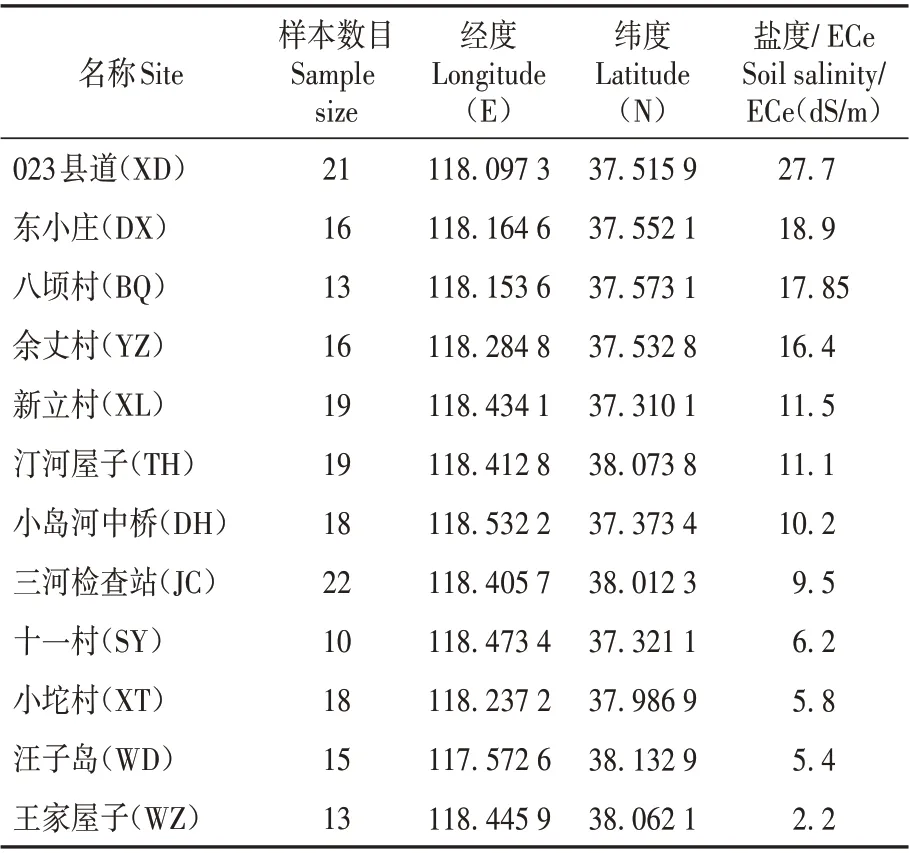
表1 采集地信息和土壤盐度Table 1 Geographic location and soil salinity at the study sites
1.3 引物设计及PCR体系流程
在NCBI 网站下载4 892 条EST 序列,通过SSRhunter 在线搜索得到340 条符合条件的序列,通过Primer premier5.0 软件,共设计得到92 对引物。华大基因有限公司合成上下游引物。实验建立阴性对照,并使用ddH2O 代替模板DNA 进行PCR 扩增。实验使用10 µL 体系。包括2 µL MIX,5 µL ddH2O,正反引物各1 µL,DNA 模板1 µL,反应流程为94℃预变性3 min,94℃变性30 s,退火时间设置30 s,72℃延伸80 s,35个循环,72℃延伸10 min,4℃保存。
1.4 SSR实验
进行PAGE 凝胶电泳及银染,利用荧光标记引物进行PCR,用3730XL 毛细管电泳检测STR 分子标记的方法对目的群体进行多态性微卫星位点的筛选。荧光PCR 产物信号检测的结果由Gene Marker v2.2.0软件解释,毛细管电泳图谱以PDF的形式呈现[7](见图1)。毛细管电泳实验由上海迈浦生物科技有限公司完成并得到18 对具有多态性的位点(见表2)。
1.5 数据分析

图1 二色补血草部分样品在SSRLB1引物处扩增的毛细管电泳检测Fig.1 Capillary electropherogram obtained by SSRLB1 primer of some samples from Limonium bicolor
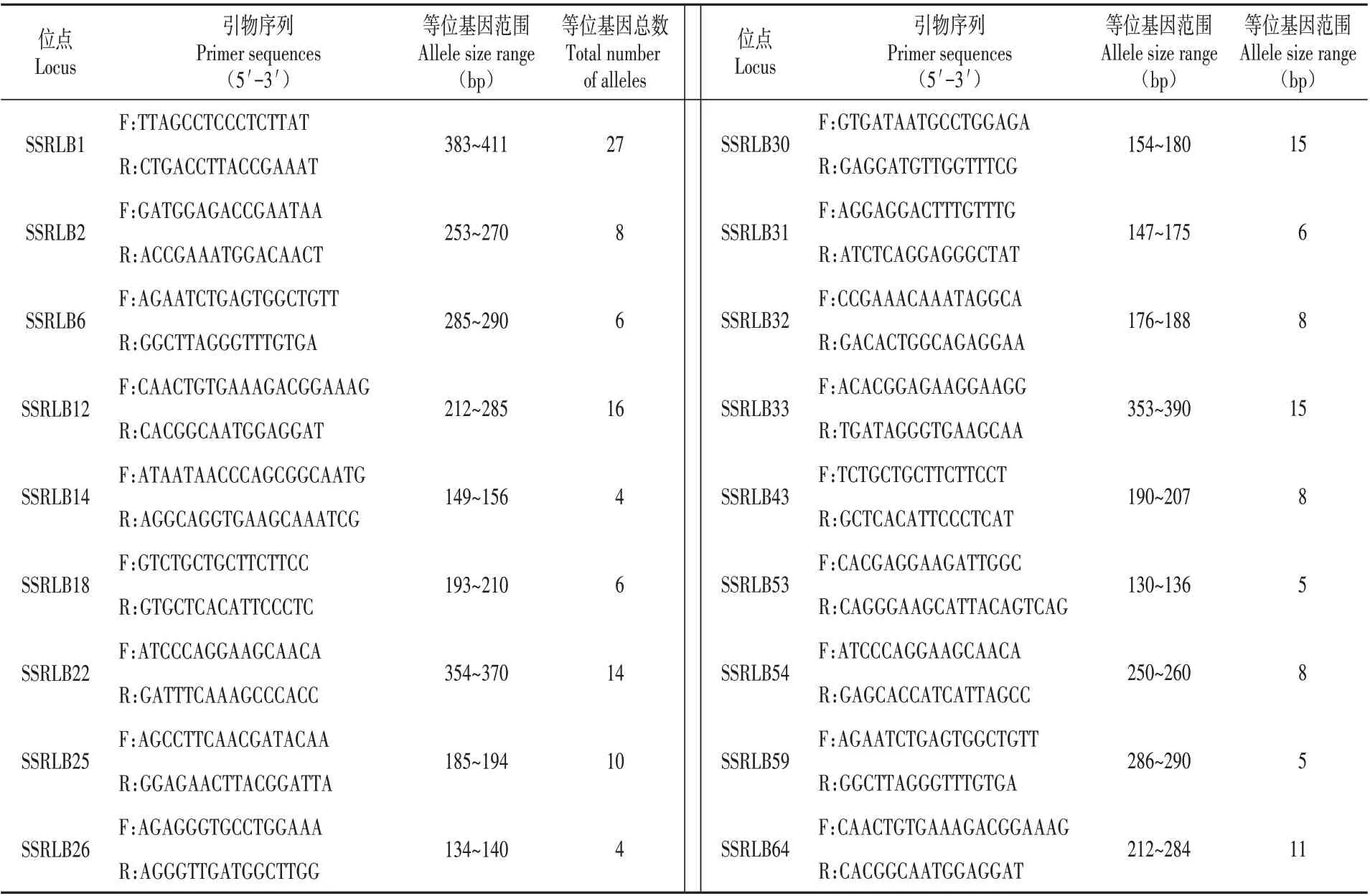
表2 18个微卫星位点的引物序列和等位基因多样性信息Table 2 Primer sequences and allelic diversity information for 18 microsatellite loci
运用遗传数据分析软件GENAIEX6.502 计算遗传多样性参数,可得出平均等位基因数目(Na)、有效等位基因数目(Ne)、平均观察杂合度(Ho)、平均期望杂合度(He)、香农信息指数(I)等遗传多样性参数,主成分分析(PCoA)结果;F-统计量和基因流(Nm)等遗传分化参数。利用ARLEQUIN version3.5.2.2软件进行分子遗传方差分析即AMOVA分析[8]。使用IBM SPSS statistics19.0 做出遗传多样性参数(He)、(I)与盐度的差异显著性分析。遗传结构使用STRUCTUREv2.3.4 马尔科夫链(Markov Chain Monie Corfo,MCMC)完成[9]。MEGAv6.0软件做出聚类分析图。
2 数据结果
2.1 平衡检测
12个种群的18个SSR 位点共获得216个种群位点统计数据,其中134 个与哈迪-温伯格平衡(HWE)一致,76 个显著偏离(P<0.01)HWE 平衡,而余下的6个则为单态。
2.2 遗传多样性结果分析
居群水平遗传多样性:得到二色补血草居群平均等位基因数目(Na)介于4.111~5.167,平均是4.644,最高群体是XD;有效等位基因数目(Ne)介于2.49~3.197,平均2.729,最高群体是XL。香农信息指数(I)介于0.967~1.14,平均1.037,最高群体是XL。平均观察杂合度(Ho)范围是0.305~0.540,平均0.430,最高群体是XL。平均期望杂合度(He)在0.486~0.556,平均为0.529,最高的群体是WZ。平均观察杂合度与平均预期杂合度之间的差异最小是种群XL,其次是XT,最大的是BQ,固定指数(F)是正的,其中居群XL最小,说明此居群相比于其他群体其杂合子较多(见表3)。
位点水平遗传多样性:通过使用具有显著多态性的18 对SSR 引物进行202 个二色补血草样品的扩增,得到176 条带。每对扩增出的多态性条带是4~27 条,平均每对是9.7 条。扩增得到平均等 位 基 因 数 目(Na)范 围 是2.083(SSRLB18)~11.667(SSRLB1),平均4.644;有效等位基因数目(Ne)范围是1.176(SSRLB18)~7.952(SSRLB1),平均2.729;香农信息指数(I)范围是0.255~2.221,平均1.037;平 均 观 察 杂 合 度(Ho)范 围 是0.055~0.829,平均0.430;平均期望杂合度(He)范围是0.137~0.869,平均0.529。固定指数(F)总体介于-0.545(SSRLB53)~0.836(SSRLB31),平 均 为0.198,其中4个位点为负值,负值出现位点为SSRLB2、SSRLB18、SSRLB53、SSRLB54,说明这四个位点杂合子过量;其余为正值,说明含有杂合子较少(见表4)。
2.3 遗传分化
二色补血草群体近交系数(FIS),范围为-0.546~0.835,SSRLB2、SSRLB18、SSRLB53、SSRLB54 四个位点为负值,平均为0.196。基因流(Nm)为2.188~13.741,平均4.106。遗传分化系数(FST)平均为0.067,范围是0.018(SSRLB53)~0.103(SSRLB14)(见表5)。在本研究中,使用ARLEQUINv.3.5 对所有居群在物种水平和分组水平进行AMOVA 分析。结果为96%的遗传变异来自居群内,4%的遗传变异来自于居群间(见表6)。
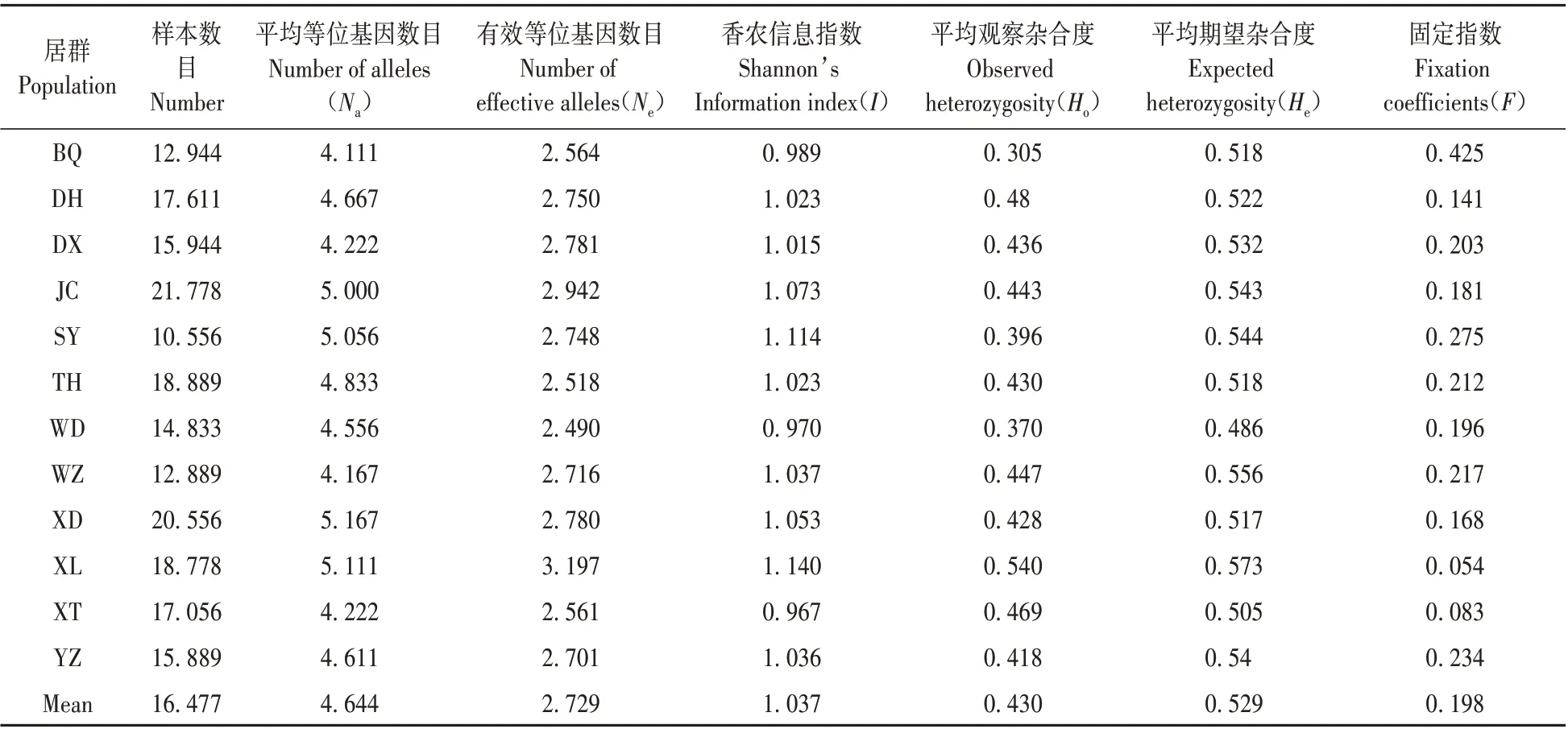
表3 二色补血草居群遗传多样性参数Table 3 The parameter of genetic diversity for L.bicolor populations
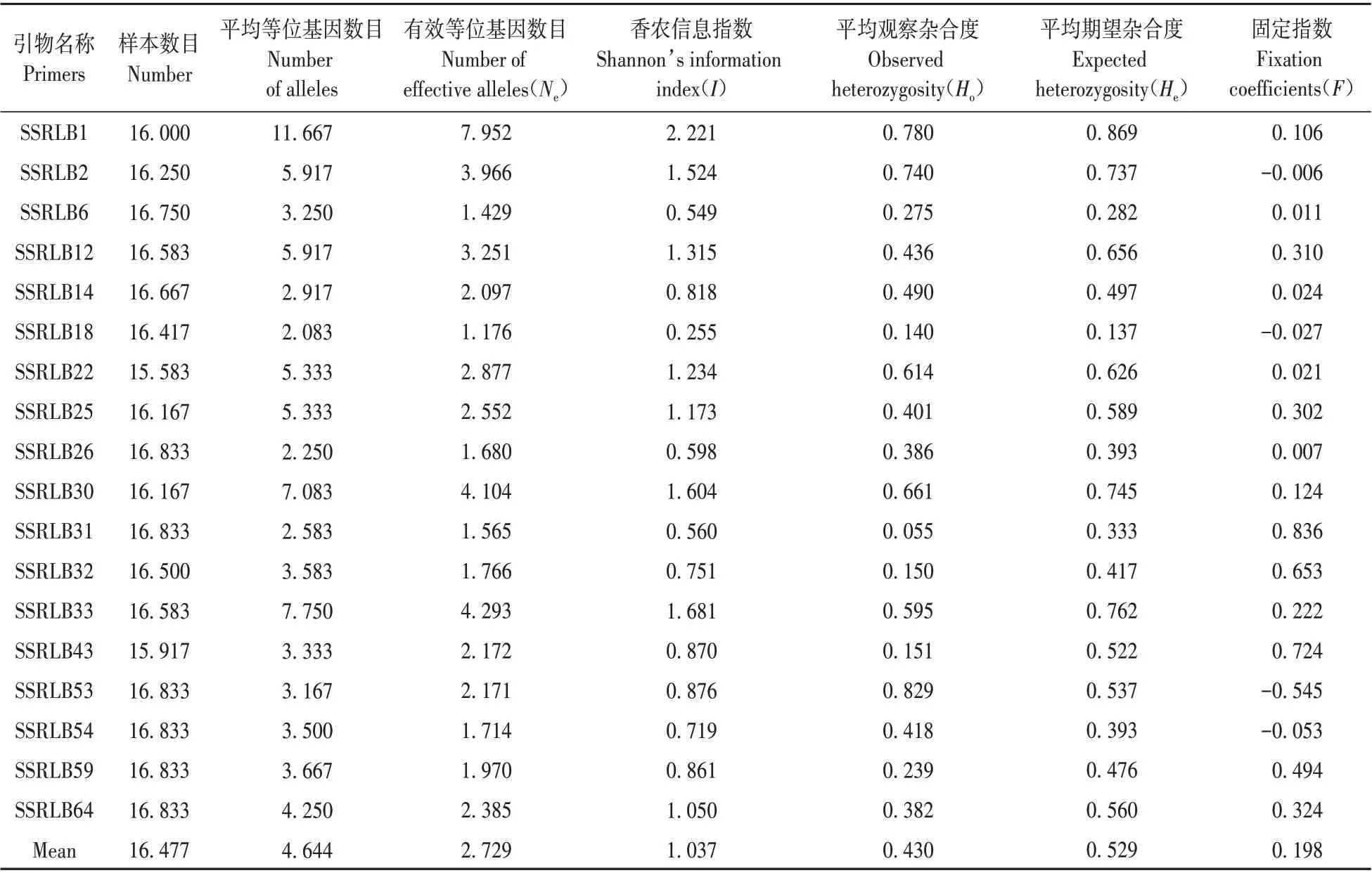
表4 18个位点遗传多样性参数Table 4 Diversity information parameters at 18 SSR loci
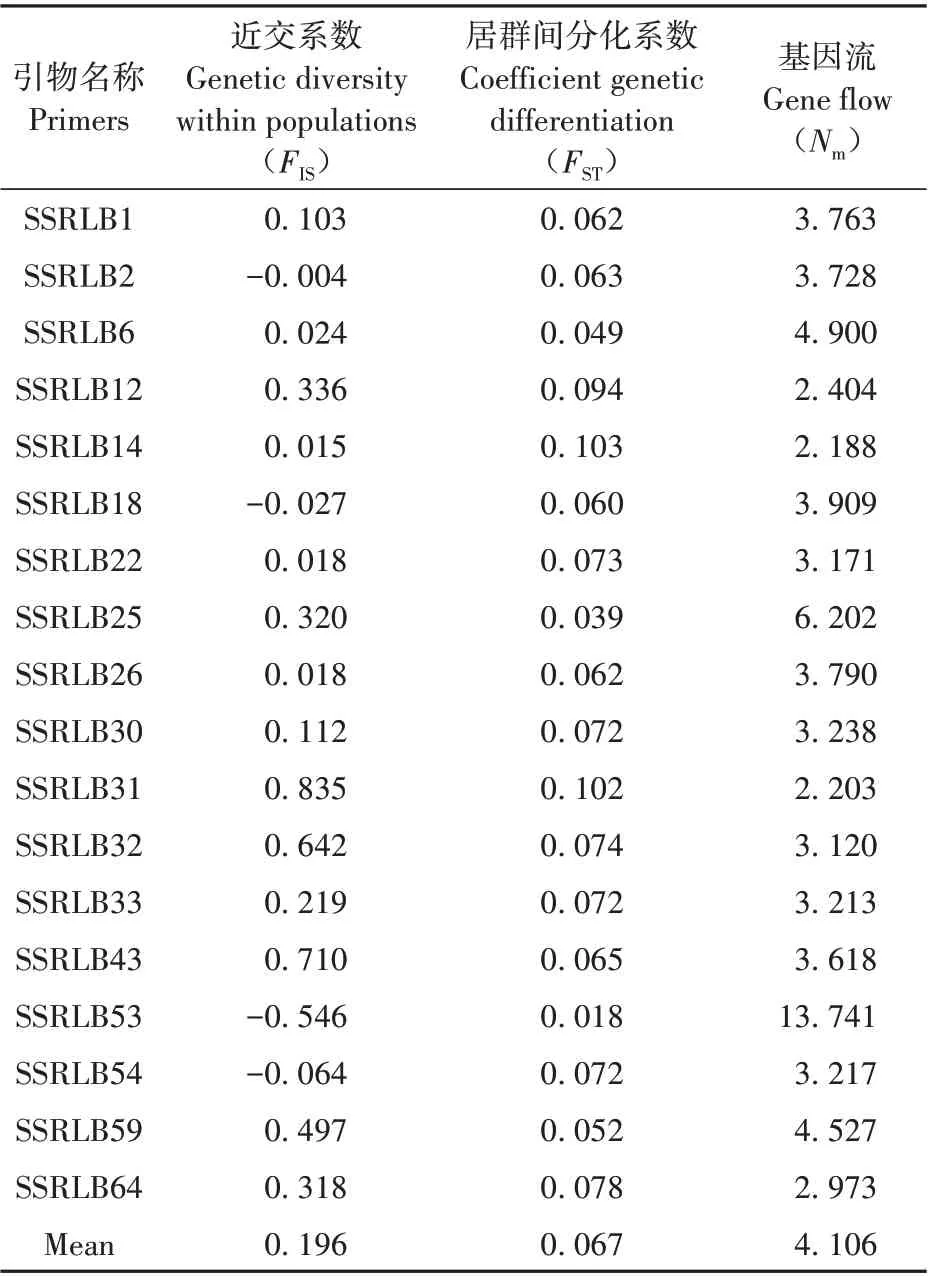
表5 18个位点遗传分化参数Table 5 Genetic differentiation parameters at 18 SSR loci
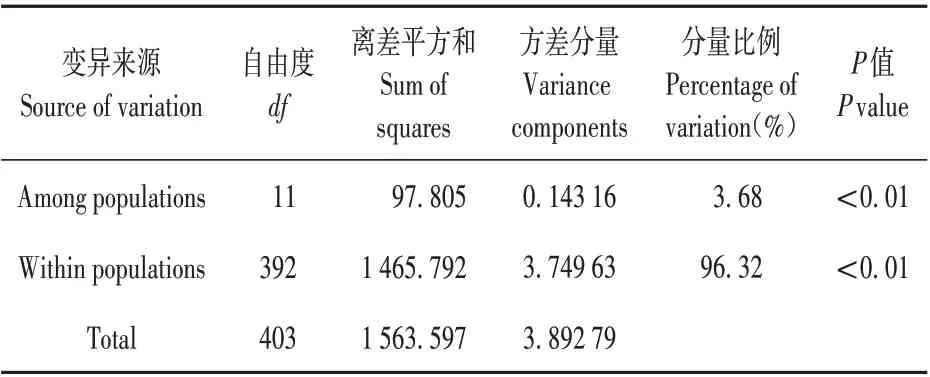
表6 18个位点分子方差分析结果Table 6 AMOVA results as a weighted average over eighteen microsatellite loci
2.4 遗传结构
使用GENAIEX6.502 软件对12 个居群进行基于遗传距离构建的主成分(PCoA)分析,可以反映亲缘关系的远近。图中两点之间的距离,代表两个居群间的遗传距离,两点之间的距离越近,说明两个居群间的遗传距离越近。图中距离相近的群体,实际地理距离并不接近,结果表明二色补血草的遗传距离与地理距离无相关性(见图2)。
通过Mantel 检验,结果显示:黄河三角洲二色补血草居群的遗传距离与地理距离之间无相关性(R2=0.001 2,P=0.374)(见图3);遗传距离与土壤盐度差异没有相关性(R2=0.005 8,P=0.4)(见图4);土壤盐度与遗传参数He、参数I关系结果显示:(R2=0.001,P=0.918),说明土壤盐度和遗传参数I没有相关性;(R2=0.005 9,P=0.821),说明土壤盐度和遗传参数He没有相关性(见图5)。
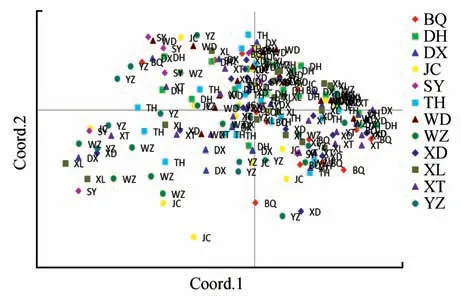
图2 基于SSR方法的主成分分析Fig.2 Principal component analysis based on SSR
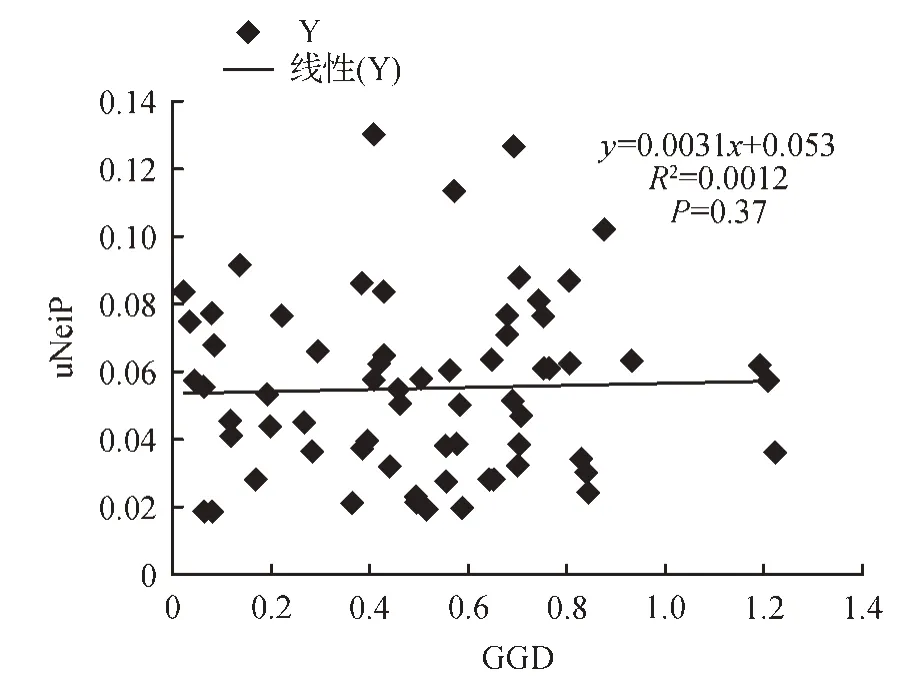
图3 SSR分析的遗传距离矩阵与地理位置矩阵的相关性Fig.3 The relevance between genetic distance matrix and geography distance matrix based on SSR
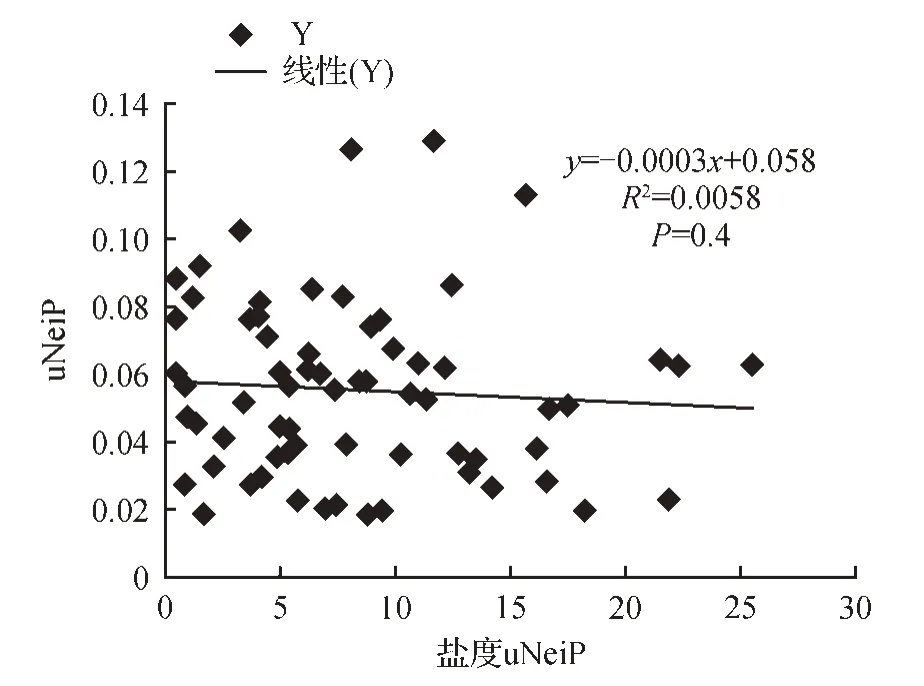
图4 SSR分析的遗传距离矩阵和土壤盐度矩阵的相关性Fig.4 The relevance between genetic distance matrix and salinity matrix based on SSR

图5 土壤盐度和居群遗传多样性参数(I)与(Ho)的相关性Fig.5 Correlation between soil salinity and population genetic diversity in YRD
通过运行软件并使用类平均聚类法(UPGMA)计算遗传距离。这种非加权算术平均聚类法灵敏度高,真实性强,具有单调性,应用广泛,聚类效果较好。XD 和XT 之间的遗传距离最小,遗传关系最接近,但地理距离不是最近的。说明遗传距离与地理距离无相关性,聚类结果见图6。
运行STRUCTUREv2.3.4 软件,设置Length of burn-in-period 为100 000,Number of MCMC replications after burn in 为1 000 000,K值定义为16,每个K 值运行10 次。点击Bar plot 出现彩图结果。将运行完的结果压缩(Result.zip),使用Structure Harvester 在线工具,判断群体的Clusters 即最可能的K 值。通过deltaK 确定最佳K 值[10],CLUSTERS重 复 抽 样 分 析,利 用CLUMPP1.1.2[11]软 件 和CLUMPAK 文件推断最佳K 值,运行次数10 次,将上步所得结果置于Distruct1.1[12]软件目录下,图形化显示结果。随着K 值增加,当K=5 时,△K 值达到峰值,当K 值继续增大,△K 值急剧下降。表明K=5 时,是比较适合的分组。不同的颜色代表不同的组,颜色的比例越大,二色补血草资源被分成相应组的可能性越大。属于相同颜色的群体显示出相对明显的区域特征[7](见图7~8)。

图6 二色补血草12个分布居群基于SSR分析的UPGMA聚类Fig.6 UPGMA dendrogram of twelve populations in L.bicolor base on SSR analysis using Nei’s unbiased genetic distances
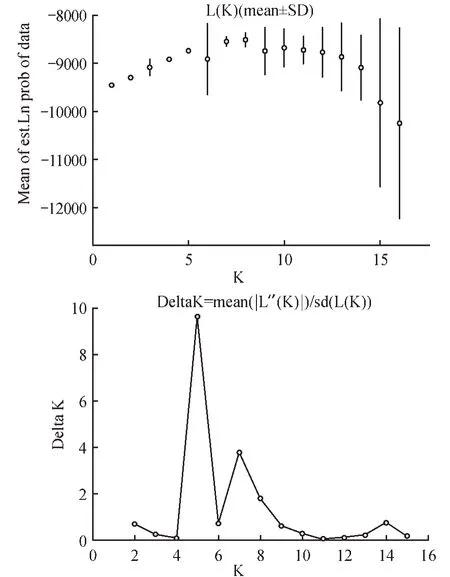
图7 K值与LnP(D)、△K值变化趋势图Fig.7 Line chart of K with LnP(D)and △K
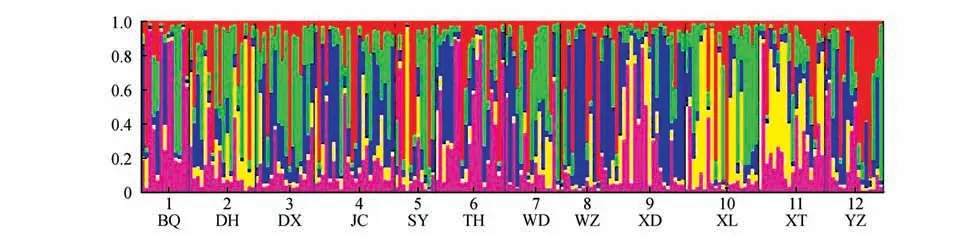
图8 基于EST-SSR标记的12份二色补血草种质群体遗传结构Fig.8 Population structure of 12 L. bicolor germplasms based on EST-SSR markers
3 讨论
3.1 遗传多样性讨论分析
SSR 分子标记揭示了分子水平上的遗传多样性[13]。平均等位基因数(Na)平均是4.644,说明二色补血草居群具有一定遗传多样性。采用香农多样性指数评价种质资源的遗传多样性,平均是1.037,也表明二色补血草居群具有一定的遗传多样性。一般认为杂合度高于0.5 的种群具有较高的遗传多样性[14],二色补血草种群平均期望杂合度平均是0.529,并且12个群体的平均期望杂合度只有WD 是0.486,其余最小0.505,说明其具有较高的遗传多样性。观察杂合度和期望杂合度的差值可用于测量居群的遗传多样性[14],在本研究中,期望杂合度与观察杂合度之间差值最小的是XL,表明遗传多样性最高,差值最大的是BQ,说明遗传多样性最低,居群间的遗传多样性比较可以指导二色补血草的种质资源收集。观察杂合度易受样本量的影响。在利用SSR 位点进行遗传分析时,要求居群个体在20~40,本研究每个居群为10~25 个个体,能够满足筛选多态性微卫星引物要求[15]。
遗传多样性的高低与物种寿命、地理分布、繁殖特性和传播媒介等有密切的关系[16]。二色补血草通常使用种子繁殖,但其遗传性非常特殊,具有大量不育分枝,具有纯合不育的特征[17]。二色补血草是两年生植物,花期较长,种子较小,花萼宿存,风力传粉,使得二色补血草具有较高的遗传多样性。保证二色补血草长期生存应减少人为干扰,合理利用资源[18],在此基础上,扩大居群数量,加快收集和育种工作[19]。研究二色补血草遗传多样性,可以为保育策略的制定、保持进化潜力措施的制定提供一定科学依据[20]。针对黄河三角洲应该完善种质资源收集的配置工作,恢复天然种群[21]。
3.2 遗传分化讨论分析
当测得FST在0.05~0.15时,表明居群间存在着中等程度的遗传分化。研究得到二色补血草FST为0.067,表明群体间存在6.7% 的遗传变异。93.3%的遗传变异存在于居群内。Nm>4,每代移动的个体数(Nm)>4,表明较大的基因流有效抵抗遗传漂变,居群间交流频繁。分子差异分析96%的遗传差异来自居群内,只有4%的遗传差异属于居群间。不同的居群间存在频繁的基因交流。基因流可以使遗传信息产生新的遗传组合,使得个体适应性更强[22]。黄河三角洲河流众多,黄河水域的传播作用,使二色补血草种子沿着黄河向不同地区延伸,不同居群的基因交流频繁,居群间遗传变异微弱[23]。
3.3 遗传结构讨论分析
主成分分析、Mantel-test检验、UPGMA 聚类分析、STRUCTURE 结构分析结果一致,均表明黄河三角洲二色补血草遗传距离和地理距离无相关性,居群间遗传信息交流频繁,遗传差异较小。12个二色补血草居群的遗传距离和土壤盐度无相关性,再次验证二色补血草是典型的盐生植物。虽然对二色补血草的培育进行了大量的研究,但未进行过大量的人工栽培和一些项目的推广[2],在此基础上,我们建议保持居群的完整性,建立保护点以保护自然种群,建立种质资源库、基因库等[24]。
4 结论
黄河三角洲二色补血草SSR 分析结果表明,二色补血草具有较高水平遗传多样性。遗传分化是中度的。遗传差异主要来自居群内部。并且遗传结构分析表明12 个居群间地理位置与土壤盐度均和遗传距离无相关性。下一步有待在其表型水平上进行多样性探索,以期获得更全面的二色补血草遗传多样性信息[7]。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
种子(2021年3期)2021-04-12 01:42:22
草地学报(2018年5期)2018-11-07 02:25:00
外语教学理论与实践(2016年1期)2016-06-11 05:51:48
广东海洋大学学报(2015年3期)2015-12-22 10:05:30
华南农业大学学报(2015年5期)2015-12-04 03:04:38
中国洗涤用品工业(2015年5期)2015-02-28 19:02:26
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:20:57
华东理工大学学报(自然科学版)(2014年1期)2014-02-27 13:48:29
