
板栗STP家族基因鉴定与分析
2020-02-26 01:04程运河程丽莉曹庆昌兰彦平
西南农业学报 2020年10期
程运河,程丽莉,曹庆昌,兰彦平*
(1.北京市林业果树科学研究院,北京 100093;2.国家林业局板栗工程技术研究中心,北京 100093)
【研究意义】板栗(Castaneamollissima)是原产于我国的重要阔叶经济林,雌雄同株异花,且雌、雄花比例严重失衡(1∶2400~4∶500)[1]。根据形态发生顺序,板栗花芽分化阶段可分为:生理分化期、花序原基分化期、苞片原基分化期、花簇原基分化期和雄(雌)蕊原基分化期。目前研究认为营养和激素是影响板栗花芽性别分化的重要原因[2]。徐娟[3]和陈在新等[4]的研究表明,结果母枝中的蔗糖含量高于雄花枝。生产中常通过修剪、施肥等方式改善树体营养,促进雌花分化,增加产量[5]。因此研究板栗的糖的分配对于调控板栗花芽性别分化、增加雌花比例具有重要意义。【前人研究进展】糖是主要的能量来源,也是重要的信号物质,广泛参与植物的生长发育和胁迫响应[6-8]。高等植物中,碳水化合物由源组织-叶片通过光合作用产生后,被运输到幼嫩叶片、根、花、果实等库组织,为正常生命-活动和生长发育提供能量[9-10]。糖主要以蔗糖形式通过韧皮部从源组织运输到库组织[11]。蔗糖运输到库组织时先在细胞外被分解成单糖,再由STP(SUGAR TRANSPORT PROTEIN)蛋白将单糖提取到库细胞中[12-13]。拟南芥(Arabidopsisthaliana)中的STP1(SUGAR TRANSPORT PROTEIN 1)基因是植物中第一个被鉴定到的单糖转运蛋白基因[14]。目前已在拟南芥基因组中鉴定到14个STP基因(STP1-14),且这些基因均属于MONOSACCHARIDE TRANSPORTER (MST)-like superfamily[15-16]。近年来,高粱(Sorghumbicolor)[17]、梨(PyrusbretschneideriRehd)[18]等植物中的STP基因也相继被鉴定。STP基因的表达具有时空特异性。拟南芥中AtSTP8在配子体中均高表达。AtSTP12与内珠皮的发育和侧根的发生有关。AtSTP1和AtSTP5则在种皮中表达[13, 19]。在拟南芥中,AtSTP3和AtSTP14主要在成熟叶片中表达[20-21]。此外,STPs的表达受其转运底物、生物胁迫和非生物胁迫的影响[8, 13, 22]。不同的STP转运单糖的种类和能力各异[16, 20-21]。STP蛋白转运的单糖主要包括葡萄糖、果糖、半乳糖、木糖和甘露糖等[11, 23]。大多STP家族成员能够转运多种单糖。少数STP蛋白对单糖的转运具有特异性。如木薯中的MeSTP4和MeSTP10只能转运麦芽糖[11]。STP蛋白对糖的转运是通过其跨膜结构域和疏水片段实现的[12, 24]。典型的STP蛋白均存在12个跨膜结构域[11, 13, 25]。跨膜结构域能将STP蛋白定位于细胞质膜上[24-25]。STP蛋白细胞质膜外侧的均疏水片段能够形成一个Lid区域,将糖和质子与质外体隔离,形成特殊的微环境,以实现糖的跨膜转运[12]。随着高通量测序技术的进步和成本的降低,基因组测序已在植物研究中广泛应用。北京农学院秦岭研究团队基于二代和三代测序,完成了中国板栗基因组测序、组装和注释。该研究组装的板栗基因组大小为785.53 Mb,Contig N50为944 Kb,能够覆盖96.7 %的基因组,注释了36 479个基因。该基因组图谱的绘制为板栗的进化起源、分子生物学研究和遗传改良提供了重要的参考依据。【本研究切入点】本研究利用生物信息学方法,在板栗基因组中鉴定STP家族成员,并对其基因结构、蛋白保守结构域、表达模式等进行分析。【拟解决的关键问题】旨在提供板栗STP基本信息,解析STP的功能,为进一步研究STP家族成员在板栗花芽性别分化过程中的作用提供参考。
1 材料与方法
1.1 板栗基因组STP家族成员的鉴定
首先在gigaScience网站下载板栗基因组数据。从Pfam网站下载STP蛋白保守domain的隐马尔科夫(HMM)模型文件,标号为PF00083。利用HMM3.0软件,基于隐马尔科夫模型,在板栗基因组数据中搜索含有STP结构域的蛋白序列,阈值为E-value < e-10。将搜索到的蛋白序列放到NCBI网站,在CDD数据库中寻找保守的STP结构域,阈值设为E-value < 0.01。
1.2 板栗STP家族基因结构、保守基序分析和进化分析
TBtools软件用于CmSTP家族基因结构分析。利用MEME(http://meme-suite.org/tools/meme)在线网站,输入CmSTP家族的氨基酸序列,搜索保守基序,参数设置为:基序的重复数“any”。基序长度6~200 aa,预测基序数量10个。将得到的保守基序在Pfam(http://pfam.xfam.org/)网站进行功能注释。从Phytozome数据库下载拟南芥(Arabidopsisthaliana)、木薯(Manihotesculenta)的STP氨基酸序列,并将其与筛选到的板栗CmSTP氨基酸序列用Clustal W进行多重比对后导入MEGAX,用临接法(NEIGHBOR-joining)构建进化树,校验参数为bootsrap=1000。
1.3 板栗STP家族蛋白理化特性分析
利用在线工具ExPASy(http://web.expasy.org/protparam)分析板栗STP家族成员的等电点、长度和分子量等属性。利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测CmSTP家族蛋白质二级结构。用Plant-mPLoc软件(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)进行亚细胞定位预测。使用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)预测跨膜结构域。
1.4 板栗STP家族基因表达模式分析
混合芽是板栗形成产量的基础。检测STP在板栗混合芽早期发育中的表达量,对于研究糖代谢与板栗花发育和产量形成具有重要意义。本研究利用北京市林业果树科学研究院板栗研究室的前期研究中板栗不同时期混合花芽转录组数据,对板栗STP家族基因表达量进行分析。其取样方法如下:以北京市密云县板栗标准化栽培示范园10年生板栗优系‘YH-1’为材料,在春季萌动后,分别于2017年3月9日,2017年3月16日,2017年3月30日,2017年4月7日和2017年4月13日,取混合芽,进行转录组测序。设置3个生物学重复,每重复1株板栗。利用百迈客云平台(http://www.biocloud.net/)从5个发育时期的RNASeq数据中提取CmSTP家族成员的表达量,并利用HemI1.0软件绘制表达模式热图。
2 结果与分析
2.1 板栗STP基因鉴定
利用HMM3.0软件,在板栗基因组数据库中共搜索到17个STP家族成员(表1)。依据各基因在染色体上的位置信息,依次命名为CmSTP1~CmSTP17。CmSTP开放阅读框长度范围为843~1584 bp,其中CmSTP14开放阅读框最短,为843 bp,CmSTP6开放阅读框最长,为1584 bp。CmSTP蛋白分子量相对较大,介于32 122.5~57 788.3 kDa,等电点介于5.46(CmSTP3)~10.46(CmSTP17),其中只有CmSTP3蛋白偏酸性,其余CmSTP成员蛋白均呈碱性或弱碱性。
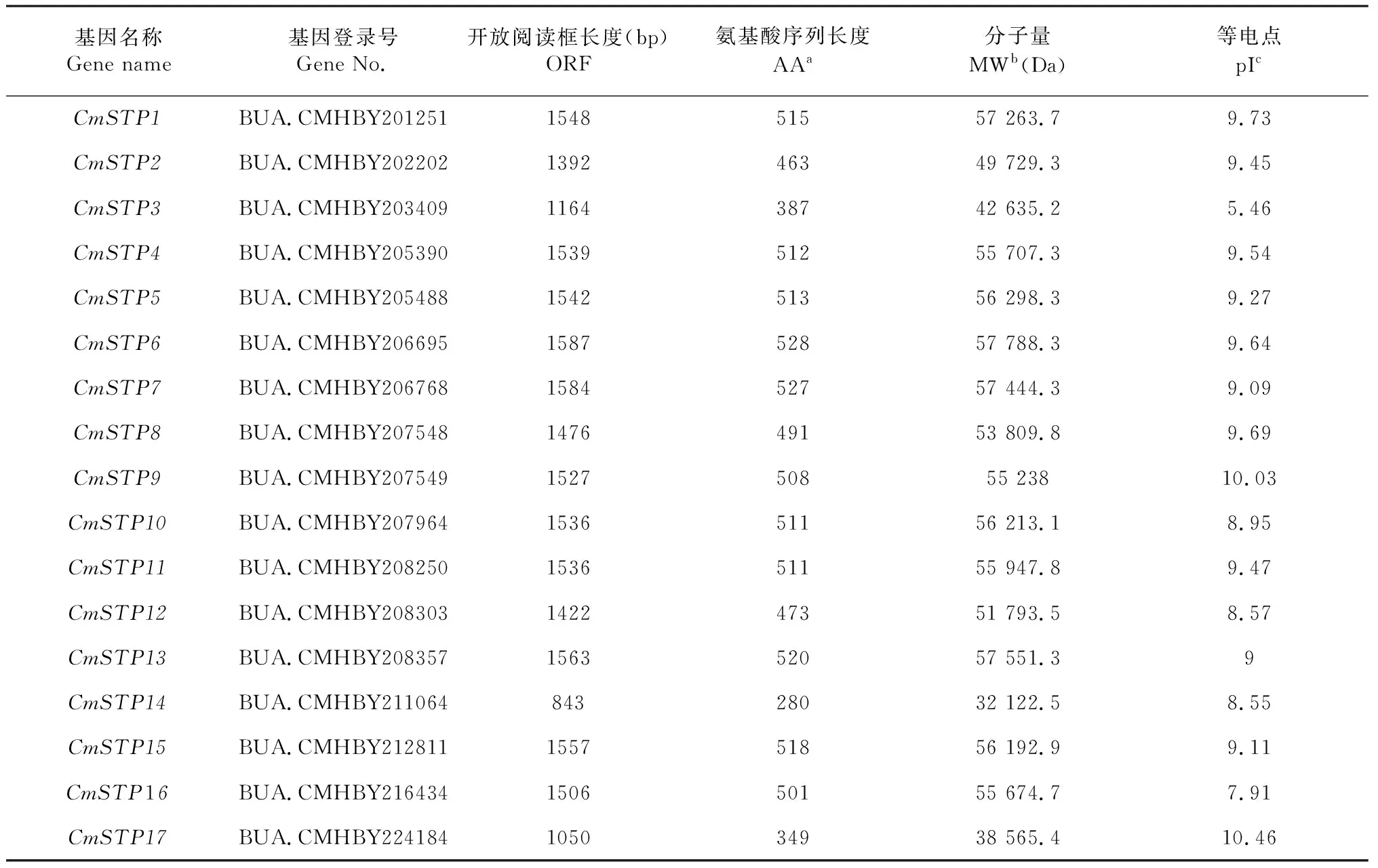
表1 板栗CmSTP基因家族信息
2.2 板栗STP结构特征
2.2.1 板栗STP基因结构分析 在鉴定得到的17个CmSTP基因中,只有CmSTP17没有内含子(图1)。其余CmSTP基因中内含子数量介于1~4个。其中CmSTP14有1个内含子,CmSTP8、CmSTP9、CmSTP12均有2个内含子,CmSTP3内含子数量最多,为4个。CmSTP13、CmSTP6、CmSTP15和CmSTP8内含子的长度明显长于其它CmSTP基因。

图1 板栗STP家族基因结构Fig.1 The structure of STP gene family in Chinese chestnut
2.2.2 板栗STP蛋白保守基序分析 使用MEME软件分析了CmSTP蛋白序列,共鉴定到10个保守基序:Motif1~Motif10(图2)。将得到的Motif序列在Pfam网站注释,结果显示除Motif5没有得到注释信息外,其余Motif均为Sugar_tr家族(PF00083)结构域(表2)。17个CmSTP蛋白保守基序数量和类型不同。CmSTP14、CmSTP3和CmSTP17保守基序数量较少,分别只有5、6、7个Motif。CmSTP2、CmSTP8和CmSTP12均有9个Motif。在所有保守基序中,只有Motif1、Motif3和Motif5同时存在于17个CmSTP蛋白中。

表2 板栗STP蛋白保守基序注释

图2 板栗STP蛋白保守基序分布Fig.2 The prediction of conserved motifs in Chinese chestnut STP proteins
2.2.3 板栗STP蛋白二级结构分析 利用在线二级结构预测软件SOPMA对板栗STP家族蛋白的二级结构进行分析(表3)。17个板栗STP蛋白二级结构均包含α-螺旋、延伸主链、无规则卷曲和β-转角。其中α-螺旋比例最高,介于46.00 %~52.95 %,其次为无规则卷曲26.68 %~31.52 %,再者为延伸主链14.59 %~19.38 %,最低的是β-转角3.44 %~5.86 %。

表3 板栗STP家族蛋白二级结构基本特征
2.2.4 板栗STP蛋白跨膜结构域及亚细胞定位预测 使用TMHMM在线工具预测板栗STP蛋白跨膜结构域(表4)。CmSTP1, CmSTP4-7,CmSTP9, CmSTP13跨膜结构域数量最多,为12个;CmSTP3, CmSTP14, CmSTP17的跨膜结构域较少,分别为5,6和8个。进一步对CmSTP蛋白N端和C端跨膜结构域进行统计,发现C端跨膜蛋白的数量较N端稳定。在C端,CmSTP3和CmSTP14的有2个跨膜结构域,CmSTP10和CmSTP16有5个跨膜结构域,其余13个CmSTP均有6个跨膜结构域。亚细胞定位预测结果显示,所有CmSTP均定位于细胞膜(表4)。

表4 板栗STP蛋白跨膜结构域及亚细胞定位信息
2.3 板栗STP系统进化分析
利用17个板栗(Castaneamollissima)STP家族成员、20个木薯(Manihotesculenta)STP家族成员和14个拟南芥(Arabidopsisthaliana)STP家族成员的蛋白序列构建系统发育树(表5)。52个STP蛋白聚为4个类群(Group I~Group IV,图3)。类群I和类群II中以木薯STP蛋白为主,且均只包含2个CmSTP蛋白(Group I: CmSTP5和CmSTP6;Group II: CmSTP12和CmSTP15)。Group III的STP成员最多(19个),以板栗STP成员为主(8个),其次为拟南芥(6个),木薯最少(5个)。类群IV中STP数量较少(9个),其中板栗STP成员为5个,木薯STP成员为3个,拟南芥STP成员为2个。从整体来看,不同物种STP成员在类群中的分布具有差别。板栗STP家族成员主要分布在类群III和IV中,木薯STP家族成员主要分布在类群I,II和III中,而拟南芥STP家族成员主要分布在类群III和IV中。

表5 木薯和拟南芥CmSTP蛋白名称及对应的基因登录号

图3 板栗(★)、木薯(●)和拟南芥(▲)STP蛋白系统进化树Fig.3 The phylogenetic tree of STP proteins from Chinese chestnut (★), Manihot esculenta (●) and Arabidopsis thaliana(▲)
2.4 板栗STP基因表达模式分析
对不同时期(S1:2017年3月9日,S2:2017年3月16日,S3:2017年3月30日,S4:2017年4月7日,S5:2017年4月13日)板栗混合芽转录组数据中CmSTP基因的表达量进行分析(图4),未检测到CmSTP1,CmSTP3,CmSTP9,CmSTP11,CmSTP14和CmSTP17表达数据,说明该6个基因可能在混合芽中不表达。其余CmSTP基因中CmSTP6,CmSTP12和CmSTP15的表达量相对较高,并且高表达主要发生在混合芽发育的早期(S1,S2)。在S3期三者的表达量急剧降低,随后维持在较低水平。CmSTP6的高表达由S1期持续到S2期,并且略有上升。而CmSTP12和CmSTP15的高表达主要发生在S1期,随后其表达量逐渐降低。虽然CmSTP2,CmST8和CmSTP13的表达量相对较低,但与CmSTP6,CmSTP12和CmSTP15的表达量变化趋势一致。
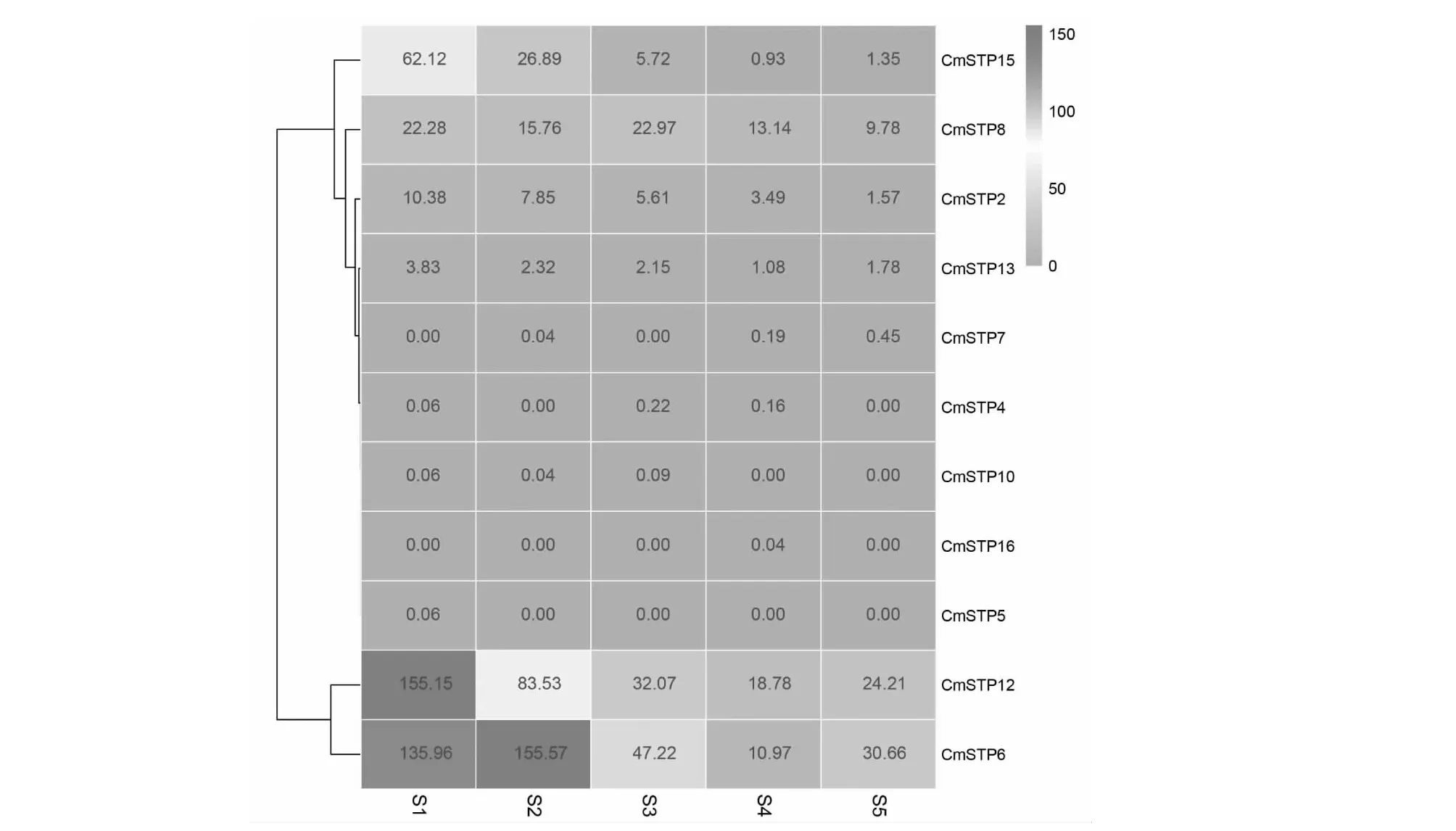
S1,2017年3月9日;S2,2017年3月16日;S3,2017年3月30日;S4,2017年4月7日;S5,2017年4月13日;S1, March 9, 2017; S2, March 16, 2017; S3, March 30, 2017; S4, April 7, 2017; S5, April 13, 2017
3 讨 论
在板栗基因组数据中共鉴定到17个STP家族成员,与拟南芥(Arabidopsisthaliana, 14个)[16]、番茄(Solanumlycopersicum, 18个)[26]、木薯(Manihotesculenta, 20个)[11]中STP的数量相近。板栗中14个CmSTP蛋白氨基酸长度介于463~528(表1),与其它物种的STP蛋白相近[11, 16, 26]。CmSTP3、CmSTP14和CmSTP17的氨基酸序列明显短于其它CmSTP(表1),但其C端均有保守的Sugar_tr家族(PF00083)结构域基序Motif1和Motif3(图2)。注释到的其它6个Sugar_tr家族结构域基序在N端和C端均有分布,但在CmSTP成员中存在不同程度的丢失,这与番茄[26]、甘蓝[25]等物种上的研究结果一致。
17个CmSTP的亚细胞定位预测均位于细胞膜,符合STP蛋白从细胞外将单糖转运到库细胞的功能[27-28]。但各CmSTP蛋白跨膜结构域的数量不尽相同。板栗中含12个跨膜结构域的STP蛋白最多,有7个,占总CmSTP数量的41.2 %。其余CmSTP蛋白均有不同程度缺失,其中N端跨膜结构域的缺失较为严重与木薯中的研究一致[11]。在其它物种上的研究推测,N端保守结构域的丢失可能是STP进化的标志[24-25, 29]。这可能是CmSTP中最保守的Motif1和Motif3均位于C端的原因(图2)。最近对拟南芥AtSTP13的研究证明,C端基序是STP蛋白细胞膜定位的核心结构域[24]。上述结果暗示,Motif1和Motif3可能是STP家族的核心结构域,在STP的细胞膜定位中发挥着关键作用。根据进化关系,板栗(Castaneamollissima)、木薯(Manihotesculenta)和拟南芥(Arabidopsisthaliana)的STP家族成员均可分成4组(图3)。禾本科植物中STP也有相似的分类[29],说明不同物种的STP在进化上具有相似性。
板栗转录组数据分析结果显示,混合芽中CmSTP6、CmSTP12和CmSTP15表达较高,CmSTP2和CmSTP8次之,其他CmSTP表达量较低或不表达(图4)。并且CmSTP6、CmSTP12、CmSTP15、CmSTP2和CmSTP8均在混合芽春季萌动期高表达,随后逐渐降低,具有时空特异性(图4)。STP基因的表达能够调控碳水化合物代谢[6, 30-31]。在植物中,通常可将组织的发育过程分为生理分化期和形态分化期。在早春,混合芽虽然未进行形态分化,但其内部的生理生化进程已经比较活跃,此时的混合芽处于生理分化期。CmSTP6、CmSTP12和CmSTP15在早春的高表达,暗示三者可能负责将树体中贮藏的糖运输到芽中,为新生组织的分化和发育提供能量和结构物质。值得注意的是CmSTP6的高表达持续到S2期,该时期是本研究材料 ‘YH-1’响应修剪、促进结果枝形成的临界时期。在拟南芥中,与CmSTP6同源关系较近的AtSTP13与碳、氮代谢有关。AtSTP13能与AtCWIN1蛋白协同作用,将质外体的蔗糖分解成葡萄糖和果糖,并以sugar/H+共运输的形式跨膜运输到细胞内[31]。在拟南芥中过表达AtSTP13可提高植株氮素利用率,促进植株生长[19]。而蒿柳(Salixviminalis)中的研究表明,碳水化合物的代谢有利于雌花的分化[32]。苹果中,扭捎可以增加尾梢的碳氮比,促进花芽形成,提高花芽比例[33]。上述结果暗示CmSTP6可能通过调控混合芽的碳氮比,参与板栗混合芽的早期发育,但其具体机制仍需进一步研究。STP的生物学功能具有多样性,本研究提供了板栗STP家族基本信息,为后续CmSTP家族成员的功能验证提供了参考。
4 结 论
本研究在板栗基因组数据库中共筛选到17个STP家族成员。保守结构域分析表明,板栗STP家族成员中共有10个保守motif,其中Motif1,Motif3和Motif5保守程度最高,存在于所有板栗STP蛋白中,其余motif在板栗STP家族进化过程中存在不同程度的丢失。进化分析结果显示,板栗与拟南芥、木薯的STP家族均可分为4类,板栗STP家族成员主要分布在Group III和Group IV中。板栗STP基因表达分析结果显示,CmSTP6、CmSTP12和CmSTP15在混合芽萌动早期的表达量较高,而其它成员表达量较低或不表达,说明STP家族基因的表达可能具有组织特异性,参与不同的生物学进程。
猜你喜欢
中华实验眼科杂志(2022年8期)2022-11-15
福建农林大学学报(自然科学版)(2022年4期)2022-11-01
南方医科大学学报(2022年3期)2022-04-13
园艺与种苗(2020年12期)2021-01-08
医药前沿(2020年28期)2020-12-02
三农资讯半月报(2020年15期)2020-08-25
课程教育研究·学法教法研究(2016年26期)2016-12-15
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
