
不同培养条件对嗜热蓝细菌生长及活性物质的影响研究
2020-02-25 11:44雒义凡李俐珩李玫锦梁园梅MaurycyDaroch
可再生能源 2020年1期
雒义凡,李俐珩,李玫锦,梁园梅,Maurycy Daroch
(北京大学 深圳研究生院 环境与能源学院,广东 深圳 518055)
0 引言
随着工业发展和人口数量的急剧增长,人为因素(如化石燃料的燃烧)成为温室气体(CO2)排放量上升的最主要因素。其中,工业排放的烟道气含热高,会释放大量热能,而设置专门的冷却装置的成本和耗能较高。因此,须寻找一种低能耗、高效率且能循环利用CO2的方法。目前的CO2固定技术主要分为物理、化学和生物3 大类,其中,物理和化学方法均存在一定局限性,且耗时长、效率低;而生物碳减排(BCM)具有效率高、能耗低的优点。BCM 是自养生物通过光合作用将大气中的CO2转化为有机碳的过程。将烟道气通入生物培养液中培养微生物,不仅冷却了烟道气,同时产生的生物质可用于生产动物饲料、医药保健品、食品色素等[1]。
蓝细菌(又称蓝藻)是能够产氧的光合自养微生物,形态结构较为简单。蓝细菌的细胞中含有多糖、类胡萝卜素、脂肪酸和藻蓝蛋白等多种具有生物活性的物质,具有良好的研究价值。蓝细菌分布范围广泛,不仅存在于淡水和海洋环境中,在温泉、南极冰川、沙漠等极端环境下也有分布[2]。其中,嗜热蓝细菌可在温泉等高温地带生长繁殖,生长温度可达45 ℃甚至更高[3];在生物碳减排技术方面,嗜热蓝细菌具有易培养、可利用高温烟道气中的CO2、抗病菌污染等优点。因此,利用嗜热蓝细菌进行工业高温烟道气的碳源减排具有良好的发展前景。
目前,针对蓝细菌适宜生长温度的研究,温度通常为常温15~35 ℃。本实验采用的蓝细菌菌株来自四川甘孜地区的温泉带[4],该温泉带的温度为30~98 ℃,两菌株采集于温度约为50 ℃的温泉中。目前,国内外针对嗜热蓝细菌的研究多集中在新种属的鉴定和筛选阶段,关于嗜热蓝细菌生理特性及工业生产应用价值的研究还较少。因此,本文通过探究采自温泉生态系统的嗜热蓝细菌分离株在不同温度和NaHCO3浓度培养条件下的生长情况,初步探索嗜热蓝细菌应用于固定工业烟道气中的CO2的可行性,为生物碳减排奠定菌种基础。
1 材料与方法
1.1 藻种
实验所用的菌株 Thermosynechococcus elongatus PKUAC -SCTE111 ( 记 为 E111) 和Thermosynechococcus elongatus PKUAC-SCT E732(记为E732)采集自甘孜州地区的温泉地带[4],现保存于北京大学环境与能源学院生物能源实验室藻种库。
1.2 培养基
嗜热蓝细菌的温度实验统一采用BG-11 培养基。配方如下 (蒸馏水配制,g/L):NaNO31.5,K2HPO40.04,MgSO·47H2O 0.075,CaCl·22H2O 0.036,Na2CO30.02,C6H8O·7H2O(柠檬酸)0.006,FeC6H5O7·NH4OH(柠檬酸铁铵)0.006,EDTA-Na2(乙二胺四乙酸二钠)0.001,H3BO42.86,MnCl2·4H2O 1.81,ZnSO40.222,Na2MoO40.39,CuSO4·5H2O 0.079,Co(NO3)2·6H2O 0.0494。配制完成后,于 121 ℃,1×105Pa 下灭菌 30 min。
嗜热蓝细菌的碳源培养实验统一采用BG-11-NaHCO3培养基。配制 1 mol/L 的 NaHCO3溶液,并用0.22 μm 微孔滤膜过滤,滤液于4 ℃储存备用;与高压高温灭菌后的BG-11 培养基混合配制不同浓度的BG-11-NaHCO3培养基。
1.3 主要仪器
SCIENTZ-II D 型超声波细胞破碎机(宁波科技生物技术有限公司)、Startorius PB-10 型pH 计(德国赛多利斯股份公司)、BSA 系列BS124S-CW型电子天平(德国赛多利斯股份公司)、LIV-3200型紫外分光光度计 (上海美谱达仪器有限公司)、Centrifuge5810R/5418R 型高速离心机 (德国Eppendorf 公司)、MGC-250P 型光照培养箱(上海一恒科技有限公司)、Epoch 型微孔板分光光度计(美国 Biotech 公司)。
1.4 实验方法
1.4.1 培养方法
种子液培养条件:培养温度为45 ℃,连续光照,转速为80 r/min,白炽灯光照强度约为 40 μmol/(m2·s);待菌种进入生长对数期后,按 2%(体积比)的接种量,接种至100 mL 新鲜BG-11培养基(250 mL 锥形瓶)
温度筛选实验: 设置 4 个温度梯度(45,50,55,60 ℃),其余条件与种子液培养条件一致,放置于摇床式光照培养箱中连续培养15 d,测定相关生长指标。各梯度分别设置3 个平行组。
NaHCO3浓度筛选实验: 设置5 个浓度梯度(0,0.1,0.3,0.5,1 mol/L),培养温度为 50 ℃,其余条件与种子液培养条件一致,放置在于摇床式光照培养箱中连续培养15 d,测定相关生长指标。各梯度分别设置3 个平行组。
1.4.2 分析方法
①测定生物量
嗜热蓝细菌生长期间,每隔24 h 进行取样,测定生长液的OD685nm,以其反映嗜热蓝细菌的生长状况。
培养15 d 后收集培养液,4 000 r/min 离心10 min 后,倒掉上清液,-80 ℃的条件下预冷 1 h,再置于冷冻干燥机中干燥24 h,测定嗜热蓝细菌的生物质干重。
②测定藻胆蛋白
取 25 mL 培养 15 d 后的 E111 和 E732 的藻液,4 000 r/min 离心 10 min,去上清,使用 15 mL的0.05 mol/L 磷酸盐缓冲液(pH 值为 6.8)重悬,置于-20 ℃的条件下冷冻4 h,在9±1 ℃的水浴中解冻,用超声波均化器将样品超声处理90 s(输出9 s,占空比为 65%)。将粗提物以 10 000 r/min(4℃,10 min)的转速离心除去细胞碎片,收集上清液。实验过程中将样品保存在冰上和黑暗中以避免提取过程中的色素降解。通过分光光度法分别测量 615 nm 处的藻蓝蛋白(Phycocyanin,C-PC),652 nm 处的别藻蓝蛋白 (也称异藻蓝蛋白,Allophycocyanin,A-PC) 和 562 nm处的藻红蛋白(Phycoerythrin,C-PE)的吸光度,并根据文献[5]中的公式计算C-PC,A-PC 和C-PE 的含量(单位为mg/mL)。

式中:A615,A652 和 A562 分别为藻蓝蛋白、别藻蓝蛋白和藻红蛋白的吸光度。
③测定脂肪酸
取5~10 mg 冷冻干燥 24 h 后的藻粉,加入200 μL 的氯仿,再加入 300 μL 含 0.6 mol/L 盐酸的甲醇溶液(盐酸和甲醇的体积比为1∶2)。将其放置于85 ℃的水浴中1 h,取出样品冷却后加入1 mL 正己烷,充分溶解后静置 1 h;取 500 μL 正己烷相加入 25 μL 的标准液(10 mg/mL),用气相色谱仪进行分析。
2 结果与分析
2.1 嗜热蓝细菌在不同温度下的生长状况
温度是影响蓝细菌生理活性和细胞内酶促反应速率的重要因素。考虑到工业烟道气温度高达120 ℃,直接通入后会引起培养系统的整体温度提升,而实验藻株的原生生存环境的温度约为50℃。因此,设置培养温度从45 ℃开始。
嗜热蓝细菌在不同温度下的生长曲线、生物量、生长速率和倍增时间分别如图1 和表1 所示。
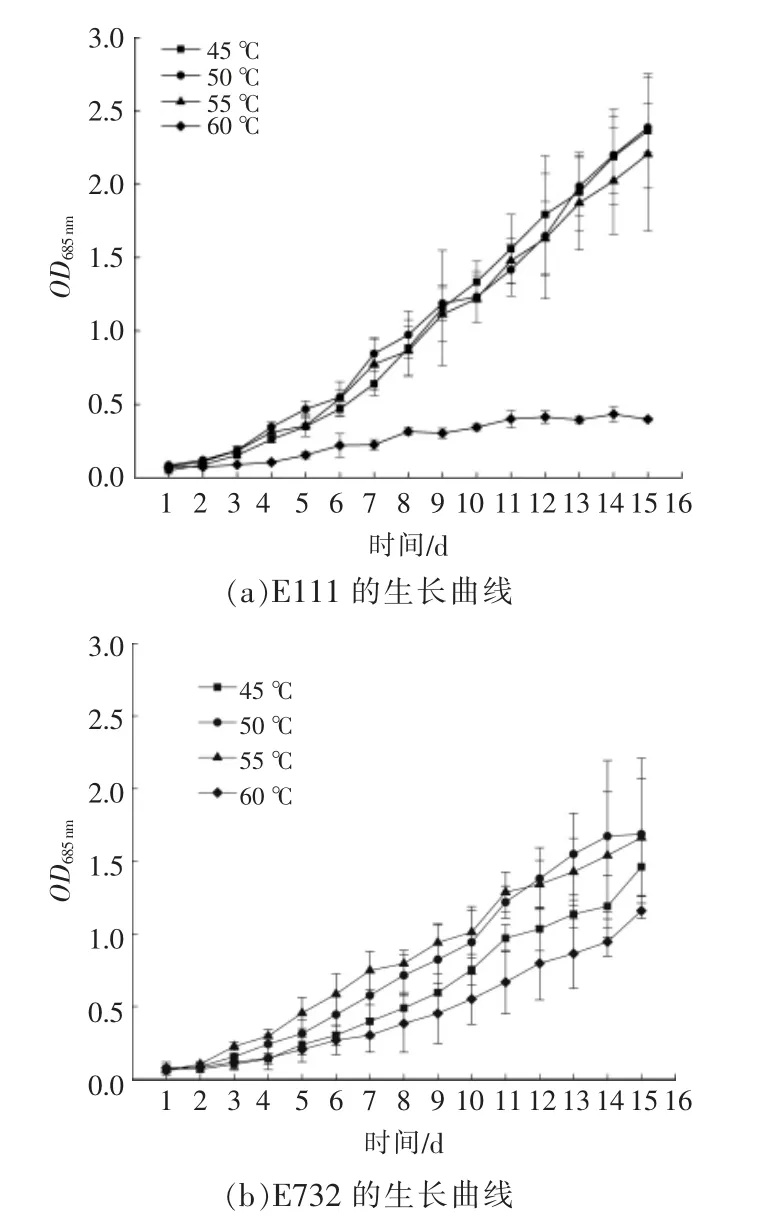
图1 嗜热蓝细菌在不同温度下的生长曲线Fig.1 Growth curves of thermophilic cyanobacteria at different temperatures

表1 E111 和E732 在不同温度下的生长指数Table 1 Growth parameters of E111 and E732 under different temperatures
从图1 可以看出,在相同温度下,E111 和E732 的生长曲线的变化趋势大致相似。多数藻种的适宜生长温度为25~35 ℃,并且藻种的细胞密度通常在30 ℃左右时达到最大值[6]。本文中,E111 和 E732 的生长温度为 45~60 ℃,高于大多数蓝细菌的生长温度。由表1 可知,在50 ℃的培养条件下,E111 和E732 的生长速率均达到最大值,生物干重高于其他温度下的培养组,倍增时间也最短。因此,E111 和E732 的最适生长温度约为50 ℃。结合图1(a)和表1可以看出:当培养温度分别为 45,50,55 ℃时,E111 均能正常生长,且生长速率相近;当培养温度为60 ℃时,E111 生长缓慢,并在第8 d 进入停滞增长期,生长速率只有0.081。从图1(b)可以看出,在 60 ℃的培养温度下,E732 可以正常生长,且第 15 d 的 OD685nm高达1.159,说明E732 体内可能存在更优异的耐热机制,能生长在60 ℃及以上的高温环境中。与夏金兰[7]诱变筛选得到的耐高温小球藻(生长温度为55 ℃) 相比,E732 是非常有潜力的耐热型光合微生物菌株。
2.2 嗜热蓝细菌在不同NaHCO3浓度下的生长状况
与CO2相比,HCO3-在水体中具有溶解度高、停留时间长等优点。嗜热蓝细菌能够利用空气中的CO2进行生长,还能够将培养液中的HCO3-作为碳源进行生长。研究嗜热蓝细菌吸收或转化HCO3-的能力,能够为转化CO2生成HCO3-实现碳减排并循环利用的工业化链条奠定基础。嗜热蓝细菌在不同NaHCO3浓度下的生长曲线、生物量、生长速率和倍增时间分别如图2 和表2 所示。结合图2 和表2 可以看出: 在培养基中添加适量的NaHCO3可以促进嗜热蓝细菌的生长,当NaHCO3的浓度分别为 0.1,0.3 mol/L 时,E111 的生长速率分别为 0.239,0.195;当 NaHCO3的 浓度为 0.5 mol/L 时,E111 菌株生长缓慢,生长速率为0.143。康丽娟[8]通过研究发现,NaHCO3浓度的提升会抑制光合色素中叶绿素a 的含量,过量的NaHCO3会伤害嗜热蓝细菌细胞的内部结构,导致类囊体减少,生长速率降低。在NaHCO3浓度相同的培养条件下,E732 生长速率的变化趋势与E111 相似;当 NaHCO3的浓度提升至 1 mol/L 时,E111 和E732 在第3 d 后均进入衰亡期。

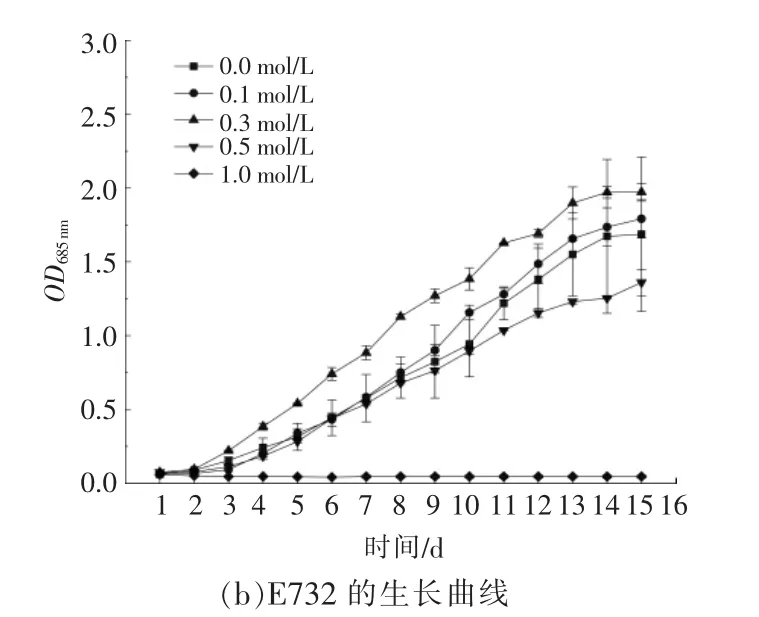
图2 E111 和E732 在不同NaHCO3 浓度下的生长曲线Fig.2 Growth curves of E111 and E732 at different concentrations of NaHCO3
目前,许多常见光合微生物如亚心型扁藻、发状念珠蓝细菌、集胞蓝细菌PCC6803 和鱼腥蓝细菌PCC7120 对于NaHCO3吸收利用能力普遍较低,最高不超过0.05 mol/L[9]~[12]。本实验所选取的两株嗜热蓝细菌吸收利用NaHCO3的浓度范围显著高于目前报道的光合微生物。蒋礼玲在对普通小球藻的不同培养模式的研究中发现,添加碳源会促进小球藻的生物量增加[13]。当NaHCO3的浓度为 0.1 mol/L 时,E111,E732 的生物干重分别可达1.663,1.160 g/L,相比空白对照组,生物干重分别约增加了84.8%,43.2%。这说明NaHCO3的添加在一定程度上提高了蓝细菌的生长速率,促进了生物质的积累。当NaHCO3浓度为0~0.5 mol/L时,其浓度越高,菌株进入稳定期的时间越快;但是,当NaHCO3浓度超过0.5~1 mol/L 时,会抑制菌株的正常生长甚至导致菌株衰亡。
2.3 藻胆蛋白
藻胆蛋白是一种高附加值蛋白,主要包括藻蓝蛋白、别藻蓝蛋白和藻红蛋白,藻胆蛋白对于研究光合作用的原初反应机理有重要意义,其作为天然色素蛋白具有广阔的应用前景。E111 和E732 在不同温度下的藻胆蛋白合成量见表3。由表3 可知: 培养温度的变化对于E111 的藻胆蛋白合成量影响甚微,藻胆蛋白合成量基本稳定在80 mg/g,约占生物总干重的8%;当培养温度为55℃时,E732 的藻胆蛋白合成量可达121.38 mg/g,是 50 ℃时的 3 倍。
E111 和E732 在不同NaHCO3浓度下的藻胆蛋白合成量见表4。从表4可以看出: 添加NaHCO3能够促进藻胆蛋白的合成,E111 和E732的藻胆蛋白合成量均高于未添加NaHCO3的空白对照组;当 NaHCO3浓度为 0.1 mol/L 时,E111 的藻胆蛋白合成量高达137.97 mg/g,约为空白对照组藻胆蛋白合成量的2 倍;当NaHCO3浓度为0.3 mol/L 时,E732 的藻胆蛋白合成量高达 176.81 mg/g。当 NaHCO3浓度超过 0.5 mol/L 时,E111 和E732 的藻胆蛋白合成量会降低,这与Schubert[14]的研究结论相一致。
在合成藻胆蛋白的过程中,各类蛋白含量的变化呈现同一趋势,即藻蓝蛋白含量>别藻蓝蛋白含量>藻红蛋白含量,其中,藻蓝蛋白是藻胆蛋白中的主要成分。由表4 可知,NaHCO3浓度的变化对藻红蛋白合成量的影响不显著,对藻蓝蛋白合成量的影响显著。在刘畅[15]的研究中,蓝细菌的藻蓝蛋白合成量最高为44.77 mg/g,而本实验中,当NaHCO3的浓度 0.3 mol/L 时,E732 的藻蓝蛋白合成量高达101.85 mg/g。这是因为HCO3-能够增加细胞中的叶绿素含量,加快卡尔文循环,从而提升光合效率。蓝细菌在固碳的同时,产生了大量的生物质,其中包含大量的藻蓝蛋白。因此,E732 是活性物质积累量较高的优势菌株。
2.4 脂肪酸
蓝细菌油脂的主要成分是甘油和脂肪酸。蓝细菌能够通过光合作用将空气中的CO2通过自身合成途径转化成脂肪酸。目前,在蓝细菌中发现的多是含 15~19 个碳原子的脂肪烃。E111 和E732 在不同条件下脂肪酸含量见表5。
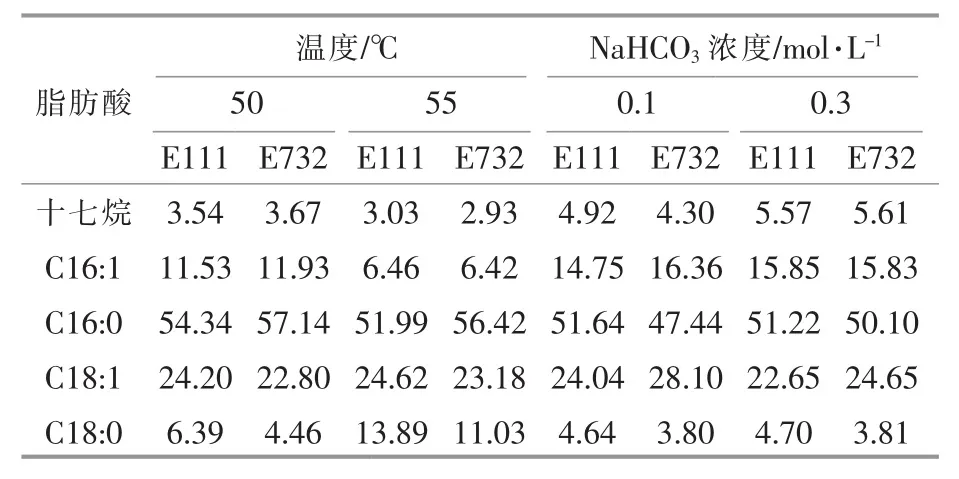
表5 E111 和E732 在不同条件下脂肪酸含量Table 5 Fatty acid composition of E111 and E732 under different conditions μg/mg
由表5 可知,构成E111 和E732 的脂肪酸的成分包括十七烷,C16∶0,C16∶1,C18∶0,C18∶1,以碳原子数为 16 和 18 的脂肪酸为主,其中,C16∶0 的含量最高。碳原子数为16 的脂肪酸占总脂肪酸的60%~70%,碳原子数为18 的脂肪酸占总脂肪酸的30%~40%。培养温度对嗜热蓝细菌的总脂含量无明显影响,但随着培养温度的降低,饱和脂肪酸和不饱和脂肪酸的比值在减小,这可能是因为微藻通过合成油脂来维持细胞膜的流动性。本实验的结果表明,随着培养温度的变化,构成E111 和E732 的脂肪酸的种类不变,但饱和脂肪酸与不饱和脂肪酸的比值随培养温度的升高而增加。脂肪酸的不饱和度随培养温度变化也与藻种有关,目前尚未有一致性规律。添加NaHCO3对于菌株合成脂肪酸无显著影响。
3 结论与展望
本文以两株采自温泉的嗜热蓝细菌为研究对象,通过设定高温(最高温度为60 ℃)、高浓度碳酸氢盐(NaHCO3的最高浓度为1 mol/L)的培养条件,筛选到能够在该条件下保持高生长速率,并积累高含量活性物质的菌株,为固定工业烟道气中的CO2奠定基础。
嗜热蓝细菌E111 和E732 的最适宜生长温度均为 50 ℃,其中,E732 可在 60 ℃的高温环境下生长,高于其他常见菌株。在适宜的温度条件下,E111 和E732 的生长速率随着培养温度的升高而加快。添加NaHCO3会促进嗜热蓝细菌的生长,当 NaHCO3的浓度为 0.1~0.5 mol/L 时,E111和E732 可以正常生长;当NaHCO3的浓度提升至1 mol/L 时,细胞生长速率降低,E111 和 E732 均无法存活。不同的培养条件会影响嗜热蓝细菌中活性物质的积累。添加NaHCO3会促进嗜热蓝细菌的生长和藻胆蛋白合成,当NaHCO3的浓度为0.1~0.5 mol/L 时,随着 NaHCO3浓度的提高,藻胆蛋白的合成量显著提高;培养温度会改变菌株细胞内不同脂肪酸类型的比例,培养温度越高,饱和脂肪酸与不饱和脂肪酸的比例增大。
综上所述,嗜热蓝细菌E111 和E732 均可在高温、 高NaHCO3浓度的培养条件下表现出良好的生长稳定性,即二者都有耐受高温和高CO2浓度的能力,且E732 的耐热能力更优异。此外,培养条件改变了嗜热蓝细菌活性物质的积累情况,还可产出有广泛应用价值的藻胆蛋白等附加产物。因此,利用蓝细菌处理高温工业烟道气固定CO2的同时降低了冷却能耗,嗜热蓝细菌E111 和E732 作为潜在的优势菌株,在工业应用和科研领域都具有良好的研究和发展前景。但是本研究筛选得到的菌株在生长速率、 生物量的积累等方面还有待提高,固碳效率和生物质副产物的合成效率也有待提升,可以利用生物分子技术对特定基因进行改造来进一步提升效率,从而将嗜热蓝细菌固碳早日应用于工业领域,实现资源利用最大化。
猜你喜欢
化工管理(2022年13期)2022-12-02
草地学报(2022年3期)2022-03-28
今日农业(2021年11期)2021-11-27
中学生数理化·高一版(2021年3期)2021-06-09
中国食用菌(2020年11期)2021-01-18
中国生殖健康(2020年5期)2021-01-18
健康博览(2020年11期)2020-11-23
中学生数理化·高一版(2020年6期)2020-07-25
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年23期)2014-03-11
