
烤烟成熟期烟叶GS同工酶活性与氮素运转的关系
2019-12-25 10:55:36周健飞武云杰张安乾彭玉富杨铁钊
作物学报 2019年1期
周健飞 武云杰 薛 刚 张安乾 田 培 彭玉富 杨铁钊,*
烤烟成熟期烟叶GS同工酶活性与氮素运转的关系
周健飞1武云杰1薛 刚1张安乾1田 培1彭玉富2杨铁钊1,*
1河南农业大学烟草学院, 河南郑州 450002;2河南中烟工业有限责任公司技术中心, 河南郑州 450000
为研究不同氮效率烤烟品种成熟期叶片谷氨酰胺合成酶同工酶活性和相关生理指标的动态变化, 深入理解烤烟叶片氮素代谢的生理生化机制。以氮效率不同的3个烤烟品种中烟100、K326和NC89为材料进行盆栽试验。在不同叶龄时期取第12片叶(自下向上数), 采用Western blot方法, 测定叶片的叶肉和主脉谷氨酰胺合成酶同工酶蛋白亚基含量, 同时对叶片NH4+浓度、总氮、质外体NH4+浓度以及氨气挥发量进行测定。结果表明, 烤烟叶片叶肉中的GS主要以GS2同工酶为主, 其蛋白亚基含量随着叶龄的增长逐渐下降, 而叶脉中GS1同工酶占主导地位, 其蛋白亚基含量呈先升高后下降的趋势。在叶龄45~65 d, 叶肉和主脉的GS同工酶活性均表现为NC89>K326>中烟100, 且品种间差异达显著水平。叶肉和主脉中GS1同工酶活性与总氮和叶片铵浓度呈负相关, 与质外体铵浓度和氨气挥发量呈正相关。而叶肉中GS2同工酶活性与总氮和叶片铵浓度呈正相关, 与质外体铵浓度和氨气挥发量呈负相关。叶脉GS2活性仅与总氮和氨气挥发量有显著相关性。氮低效烤烟品种成熟期叶片中两种谷氨酰胺合成酶同工酶活性均较低, 氮素转移和再利用能力差, 导致植株吸收的氮素以氨气形式挥发损失量大, 叶片衰老速度较快。而氮高效品种氮素同化和再利用能力较强, 氨气挥发量小, 易发生贪青晚熟。
烤烟; 谷氨酰胺合成酶同工酶; 氮素代谢; 氨气挥发
成熟期是烟叶品质形成的关键时期, 氮代谢只有适时向碳代谢转化才能形成符合工业要求的优质烟叶[1]。而氮代谢特性在不同基因型间存在差异[2]。我国植烟地区目前氮肥用量普遍偏多, 如遭遇生育后期较多雨水, 土壤中缓效氮和矿化氮释放量增加, 则背离“烟株长成肥退尽”的需肥规律, 不耐氮肥品种易形成难以烘烤的“黑暴”烟叶, 品质下降。因此, 深入研究氮素利用生理生化机制, 对于筛选和培育氮素适应范围广的烟草品种具有重要指导意义。
目前对谷氨酰胺合成酶(GS)同工酶的研究逐渐成为改良作物氮效率的热点。分子生物学研究结果表明, GS同工酶在植物的氮素转运和分配中具有重要作用[3-5]。高等植物的GS基于亚细胞定位不同可以分为两类, 一类为定位于胞液中的GS1; 另一类为存在于叶绿体中的GS2。GS1的功能是对叶片衰老时的氮源进行转移及再利用, 而GS2主要对光呼吸、硝酸还原产生的氨进行同化[6]。大量研究结果集中于粮食作物中GS同工酶的变化动态以及不同氮效率品种间的差异。但不同烤烟品种叶片的GS同工酶活性及其与相关生理指标的联系, 以及对氮效率的影响尚未见报道。本试验选用成熟期不同氮效率烤烟品种为试验材料, 采用Western-blot技术和质外体提取等方法, 探索GS同工酶活性与相关生理指标的关系, 进一步研究不同氮效率品种成熟期氮素代谢机制和分子机理, 以期为筛选氮素适应范围广的烤烟品种提供理论依据。
1 材料与方法
1.1 试验材料
于2017年在河南农业大学毛庄农场进行盆栽试验。参试烤烟品种为中烟100 (ZY100)、K326和NC89 (成熟期氮效率为NC89>K326>ZY100[7-9])。供试土壤为黄棕壤, pH值为7.79, 含有机质9.73 g kg–1、全氮0.96 g kg–1、速效氮68.71 mg kg–1、速效磷25.29 mg kg–1、速效钾95.87 mg kg–1。盆栽采用聚乙烯塑料盆, 其内径为35 cm, 高度30 cm, 装土18 kg。每盆植烟1株, 每个品种60盆, 重复3次。每盆施用纯氮4 g, N∶P2O5∶K2O为1∶2∶3, 常规水分管理。在叶龄35、45、55、65、75 d (以幼叶长1 cm, 宽0.5 cm时作为叶龄第1天)取第12片叶(自下向上数)测定烟叶叶肉和主脉GS同工酶蛋白亚基含量, 同时测定叶片及质外体相关生理指标。
1.2 测定项目与方法
1.2.1 烟叶两种GS同工酶聚合方式比较 参考文献[10]中Native-PAGE的试验方法, 以叶龄55 d时的3个参试品种的烟叶样品为试验对象。烟草叶片GS同工酶分离和酶活性检测使用3%浓缩胶(pH 6.7)和5%分离胶(pH 8.7), 浓缩胶稳定电压为80 V, 分离胶稳定电压为120 V。上样缓冲液pH为7.6, 由25 mmol L–1Tris-HCl、5% β-巯基乙醇、0.05%溴酚蓝和50%甘油组成, 取5倍的上样缓冲液和粗酶液混匀。电极缓冲液由25 mmol L–1Tris和192 mmol L–1甘氨酸组成, 电泳稳定于4℃条件下进行。
1.2.2 烟叶GS同工酶蛋白亚基含量的测定 参照Wang等[11]的试验方法。首先制备鲜叶片样品粗酶液, 随后将粗酶液与等体积的上样缓冲液混合, 沸水浴5 min变性处理。室温条件下进行SDS-PAGE电泳分离(浓缩胶5%, 分离胶15%), 凝胶的制备和电泳方法参考文献[10]。电泳结束后将蛋白转移至PVDF膜上进行Western-blot检测, 所用抗体由贵州烟草研究院馈赠。以蛋白Marker为标准, 用Quantity One (Bio-Rad,Version4.6.2)进行照相及光密度分析。
1.2.3 烟叶相关生理指标的测定 分别采用凯氏定氮法和水合茚三酮法[12]测定叶片总氮和NH4+浓度。提取质外体和测定质外体NH4+浓度参考文献[13]的方法。采用武云杰等[14]的方法测定氨气实际挥发量。
1.3 数据处理分析
用Microsoft Excel 2016求各测定指标的平均值、标准差及绘图, 用IBM SPSS Statistics 22.0进行方差分析, 对各处理差异进行LSD多重比较。
2 结果与分析
2.1 烟叶GS同工酶聚合方式比较
将烟叶的叶肉和主脉分离, 分别进行其蛋白提取液Native-PAGE并结合活性染色, 叶肉和叶脉中均只观察到1条条带(图1和图2)。由于Native-PAGE是根据GS同工酶全酶相对分子量进行分离, 说明叶片两种GS同工酶分子量比较接近, 聚体方式一致。因此可以用Western-blot测定出的GS同工酶蛋白亚基含量来间接表征GS同工酶的活性。
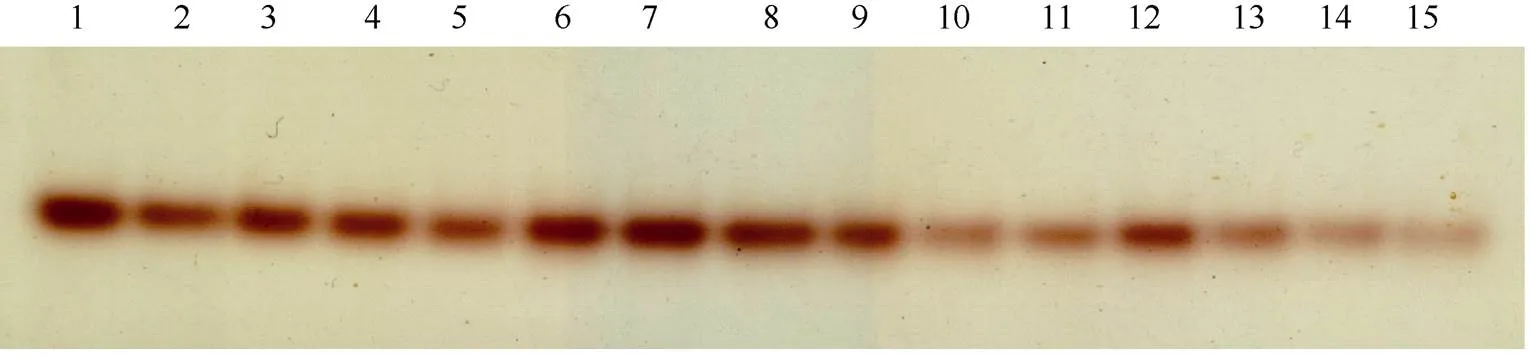
图1 叶肉GS同工酶Native-PAGE结合活性染色
1~15分别代表叶龄55 d时K326、NC89和ZY100第12片叶叶肉样品, 每个品种5次重复。
1-15 represents the 12th leaf mesophyll sample of K326, NC89, and ZY100 at leaf age 55 days with five replicates for each species.

图2 叶脉GS同工酶Native-PAGE结合活性染色
16~30分别代表叶龄55 d时K326、NC89和ZY100第12片叶主脉样品, 每个品种5次重复。
16-30 represents the 12th leaf main vein sample of K326, NC89, and ZY100 at leaf age 55 days with five replicates for each species.
2.2 烟叶GS同工酶活性动态变化
2.2.1 叶肉GS同工酶活性动态变化 由图3可知, 烤烟叶片叶肉中的GS主要以GS2同工酶为主,其蛋白亚基含量随着叶龄的增长逐渐下降, 而GS1蛋白亚基含量呈先升高后降低的趋势。由图4的灰度分析结果可知, 叶龄35 d时GS1蛋白亚基含量均较低, 叶龄45 d时GS1蛋白亚基含量大幅增加, NC89、K326和ZY100分别增加了3.78、4.52和2.54倍; 随后GS1蛋白亚基含量持续增加, 至叶龄65 d左右含量开始减少。3个参试品种在叶龄45 d至65 d的叶肉GS1蛋白亚基含量差异均达显著水平, 差异为NC89>K326>ZY100。对于叶肉GS2蛋白亚基含量来说, 35 d至45 d降幅较缓, 45 d后大幅减少, 随后一直呈缓慢下降趋势。45~65 d始终表现为NC89>K326>ZY100, 且差异达到显著水平。
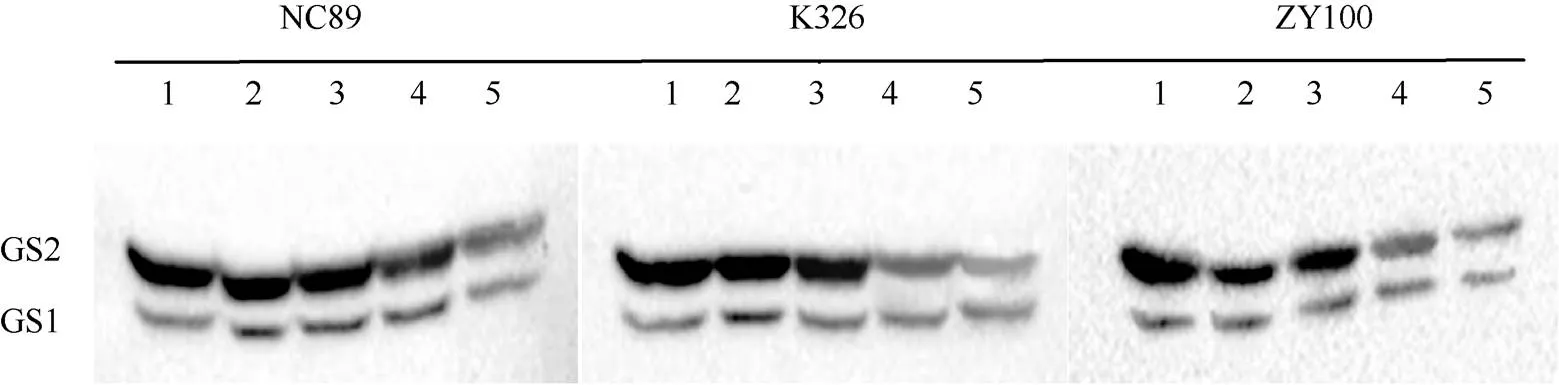
图3 叶肉GS同工酶活性动态变化谱带
1: 叶龄35 d; 2: 叶龄45 d; 3: 叶龄55 d; 4: 叶龄65 d; 5: 叶龄75 d。
1: leaf age 35 days; 2: leaf age 45 days; 3: leaf age 55 days; 4: leaf age 65 days; 5: leaf age 75 days.
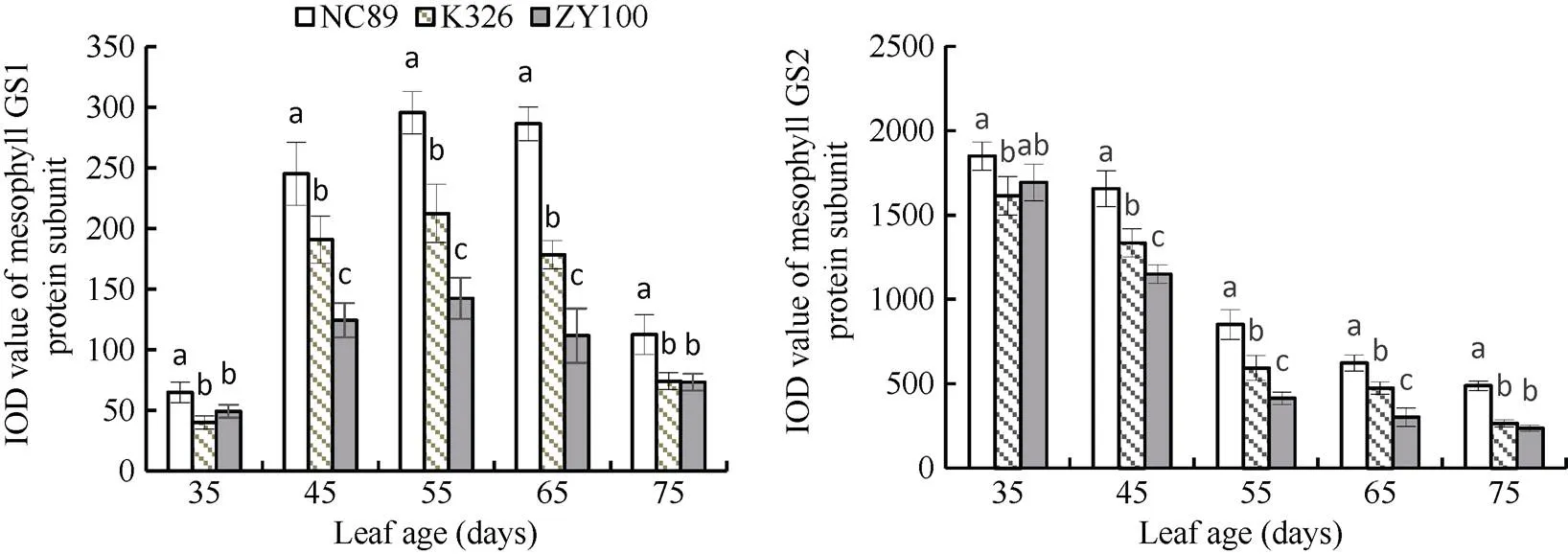
图4 叶肉GS同工酶蛋白亚基含量动态变化
图柱上同一时期不同字母表示在< 0.05水平上差异显著。
Bars superscripted by different letters are significantly different at0.05 within the same period.
2.2.2 叶脉GS同工酶活性动态变化 由图5可知, 与叶肉不同的是, 叶脉中的GS主要以GS1同工酶为主, 其蛋白亚基含量自叶龄45 d始大幅增加, 随后呈先升高后降低的趋势, 而GS2蛋白亚基含量一直呈逐渐降低趋势。由图6的灰度分析结果可知, 叶脉GS1在叶龄35 d时蛋白亚基含量较低, 45 d时含量以较大幅度增加, NC89和K326的GS1蛋白亚基含量至叶龄55 d时达到峰值, NC89分别是K326和ZY100的1.27倍和2.42倍。而ZY100在叶龄45 d时含量已最高, 随后均呈缓慢下降趋势。与叶肉GS1不同的是, 各品种在叶龄75 d时仍具有相对较高的叶脉GS1蛋白亚基含量。3个参试品种在叶龄45 d后叶脉GS1蛋白亚基含量差异均达显著水平, 且NC89>K326> ZY100。对于叶脉GS2蛋白亚基含量来说, 叶龄45 d后含量大幅减少, 叶龄35~65 d始终表现为NC89> K326>ZY100, 且差异达到显著水平。
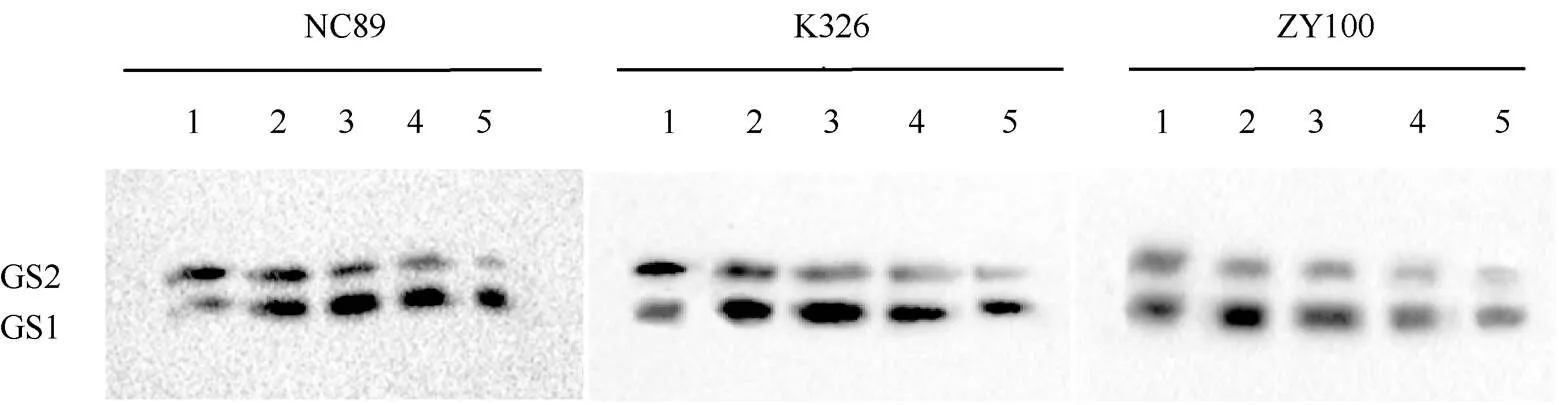
图5 叶脉GS同工酶活性动态变化谱带
1: 叶龄35 d; 2: 叶龄45 d; 3: 叶龄55 d; 4: 叶龄65 d; 5: 叶龄75 d。
1: leaf age 35 days; 2: leaf age 45 days; 3: leaf age 55 days; 4: leaf age 65 days; 5: leaf age 75 days.

图6 叶脉GS同工酶蛋白亚基含量动态变化
图柱上同一时期不同字母表示在< 0.05水平上差异显著。
Bars superscripted by different letters are significantly different at0.05 within the same period.
2.3 叶片总氮含量和NH4+浓度的动态变化
总氮含量可以反映出烟叶总体的氮素水平。由图7可知, 自35 d开始叶片总氮含量表现出持续下降趋势。45~65 d均表现为NC89>K326>ZY100, 差异达到显著水平。叶龄65 d时NC89总氮含量为1.89%, 分别是K326和ZY100的1.27倍和2.33倍。总氮降解百分数(1-最终剩余量/最大积累量)在一定程度上可以反映出烟叶氮素降解速率, NC89、K326和ZY100的总氮降解百分数分别为47.95%、63.65%和71.13%。叶片NH4+浓度在叶龄45 d时达到最大值, 随后浓度持续下降。55~75 d各品种间差异达到显著水平, 且均表现为NC89>K326>ZY100。
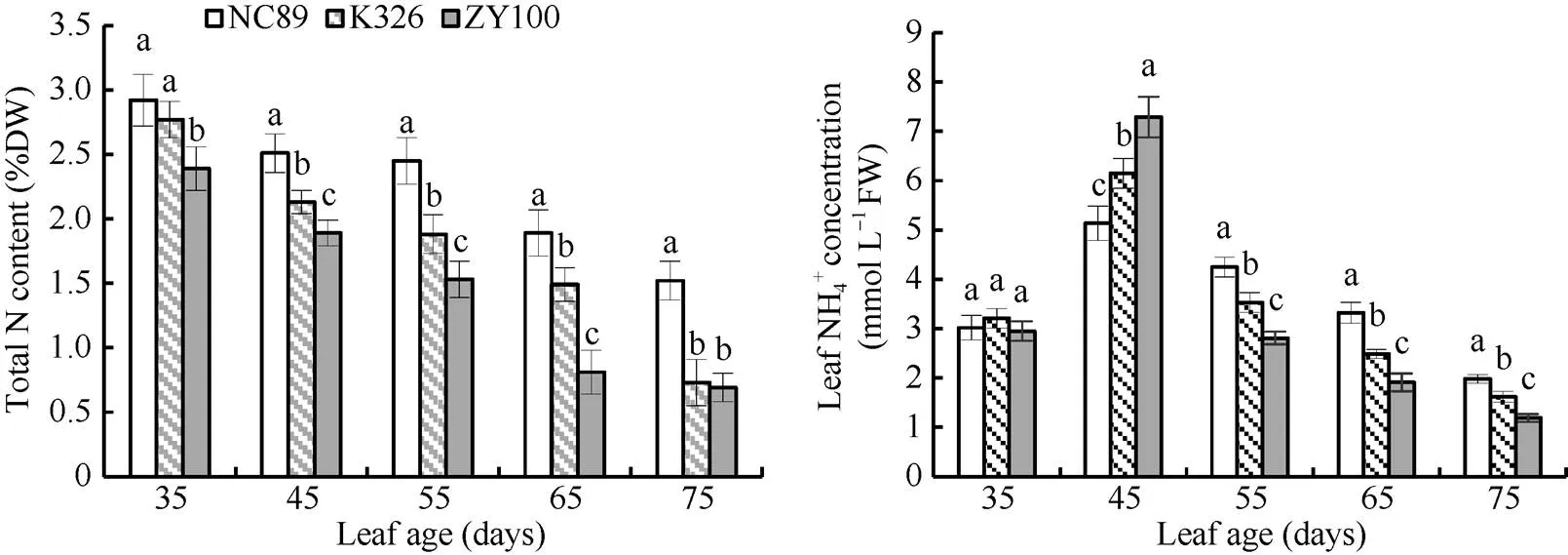
图7 叶片总氮含量和NH4+浓度的动态变化
图柱上同一时期不同字母表示在< 0.05水平上差异显著。
Bars superscripted by different letters are significantly different at0.05 within the same period.
2.4 质外体NH4+浓度和氨气挥发量的动态变化
质外体被称为NH4+的动力池, 维持着叶片和大气的气体交换。若NH4+过量积累将以氨气的形式挥发出去, 从而达到质外体中水溶态氨和气态氨的平衡[15]。由图8可以看出, 质外体NH4+浓度于55 d达到最高点, 随后大幅下降, 在45~65 d间始终表现为ZY100>K326>NC89, 其中ZY100增长幅度非常大。氨气挥发量在叶龄35 d时较低, 随后大幅升高并于55 d达到最大值, ZY100、K326和NC89分别为28.73、17.46和10.11 μg m–2h–1, 差异达到显著水平。随后尽管ZY100有较大的下降幅度, 但依然显著高于K326和NC89。
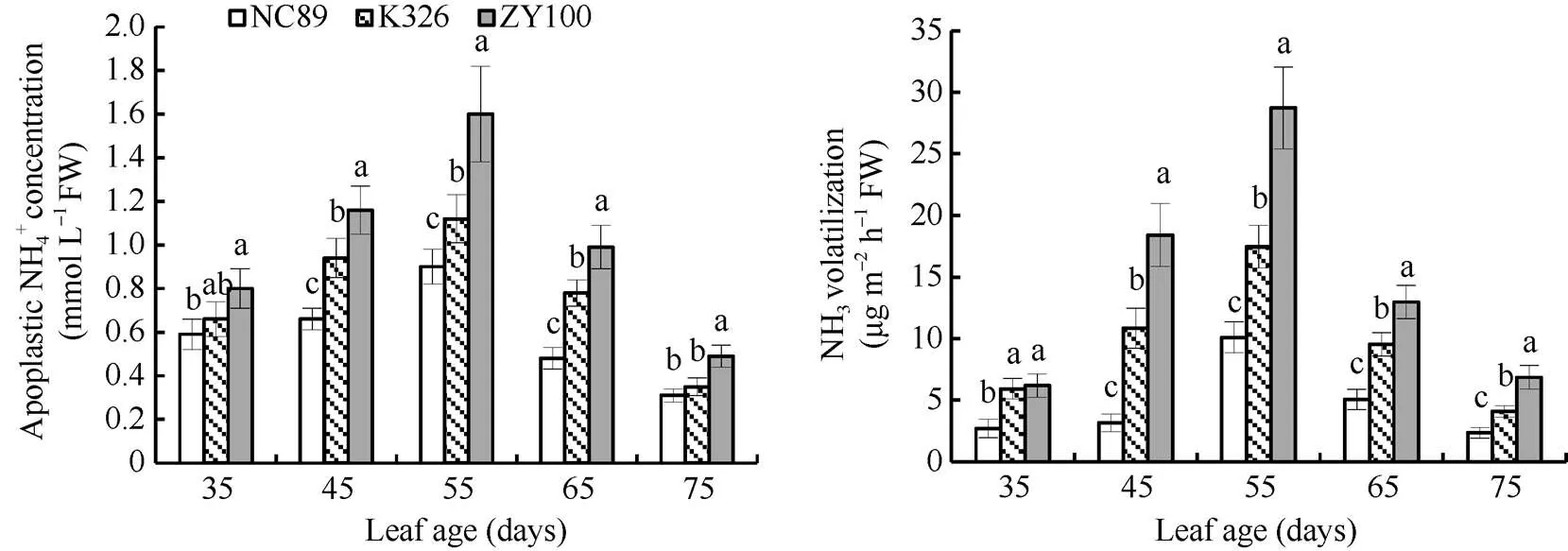
图8 叶片质外体NH4+浓度和氨气挥发量的动态变化
图柱上同一时期不同字母表示在< 0.05水平上差异显著。
Bars superscripted by different letters are significantly different at0.05 within the same period.
2.5 烟叶成熟期GS同工酶的蛋白亚基含量与生理指标的相关性分析
通过对叶龄35~75 d每隔10 d的叶肉和主脉的GS同工酶蛋白亚基含量与相关生理指标作相关性分析, 由表1可知, 叶肉GS1蛋白亚基含量与质外体铵浓度和氨气挥发量呈正相关, 与总氮和叶片铵浓度呈负相关; 叶肉GS2蛋白亚基含量与叶片总氮呈极显著正相关, 与叶片铵浓度呈显著正相关, 与氨气挥发量呈极显著负相关, 与质外体铵浓度呈显著负相关; 叶脉GS1蛋白亚基含量与叶片氨气挥发量呈极显著正相关, 与质外体铵浓度呈显著正相关, 与叶片铵浓度呈极显著负相关, 与总氮呈显著负相关; 叶脉GS2蛋白亚基含量仅与总氮和氨气挥发量有显著相关性。
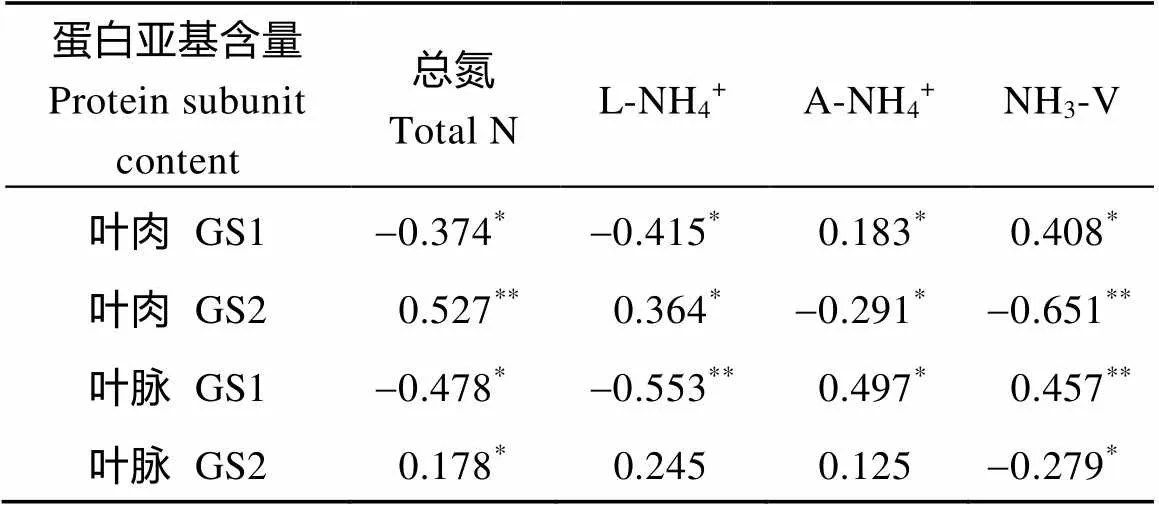
表1 GS同工酶的蛋白亚基含量与生理指标的相关系数
L-NH4+: 叶片铵离子浓度; A-NH4+: 质外体铵浓度; NH3-V: 氨气挥发量。**和*分别表示在0.01和0.05水平上显著相关。
L-NH4+: leaf NH4+concentration; A-NH4+: apoplastic NH4+concentration; NH3-V: NH3volatilization amount.**and*denote significances of correlation at the0.01 and 0.05 probability levels, respectively.
3 讨论
现蕾打顶导致烤烟的源库关系发生改变, 叶位偏下、叶龄较大的烟叶仍然是“源”, 而叶位偏上或未完全成熟的烟叶成为新的“库”, 即生长重心转移到保留的叶片上来[16]。烤烟成熟期若具有较低的氮素同化利用效率则不易贪青晚熟。衰老叶片降解的氮素既可以转移到其他叶片中再利用, 亦可以通过质外体以氨气的形式挥发出去[14], 不同氮效率烤烟品种具有不同的氮素运筹方式。近年来大量研究表明, 不同源库间GS同工酶的相对含量和时空表达特性可能是造成这种差异的重要原因之一[17]。
烟叶叶肉和叶脉中两种谷氨酰胺合成酶同工酶相对分子量较为接近, 聚体方式一致, 因此烟草GS同工酶活性可以由其蛋白亚基含量来表征。本研究中, 对于两种GS同工酶蛋白亚基含量在叶肉和主脉的变化趋势而言, 品种间表现基本一致, 说明烤烟中部叶在叶龄45 d时已完成生长发育而进入成熟时期, 氮运转进入分配再利用阶段, GS1活性升高开始行使其氮素再运转功能, 而GS2随着叶绿体的降解活性不断下降。GS2在叶肉中处于主导地位, 其活性在各时期均明显高于GS1, 而叶脉中则是GS1处于主导位置, 这可能与叶绿体在叶肉中大量分布, 而叶脉中富含输导组织、叶绿体含量较少有关。
成熟期氮低效烤烟品种ZY100和中等氮效率烤烟品种K326, 相对于氮高效品种NC89而言, 叶肉和叶脉中起同化作用的GS2同工酶在衰老启动时活性就大幅降低。ZY100的GS2活性更是低于NC89, 最终导致叶片降解的氮素再同化量较少, 表现为叶片具有较低的总氮含量和NH4+浓度, 以及较高的总氮降解百分数。同时氮低效品种ZY100叶肉和叶脉中负责氮素再运转的GS1活性均较低, 可能会导致降解氮素向“库”转移的能力相对较弱。NH4+在植物体内过量积累将产生毒害作用, 而本研究中叶片NH4+浓度在叶龄45 d之后持续下降, 质外体NH4+浓度积累高峰比叶片晚10 d左右, 且氨气挥发量在成熟期大量增加, 说明NH4+向质外体转移可能是一种缓解氨害积累的生理机制, 且有利于实现自身的氮素调亏。氮高效品种NC89在成熟期叶肉和叶脉均具有相对较高的GS1和GS2活性, 结合GS同工酶的生理功能和“源库流”理论, 较高的氮素同化效率造成了氮同化物在“源”的富集, 为成熟期烟株的代谢提供了丰富的底物, “源”压力较大, 同时由于其GS1活性较高, 转运效率高, 衰老降解的氮素被大量再利用, 致使NH4+向质外体转移量较少, 易导致贪青晚熟。源库之间的压力差和质外体生理上的差异综合导致了不同基因型烤烟成熟期氮素利用效率的差异。相关性分析也表明, 成熟期不同氮效率烤烟品种叶片质外体铵浓度及氨气挥发量与GS同工酶活性具显著或极显著相关性, 说明GS同工酶活性的差异是不同基因型间具有不同成熟期氮效率的主要原因之一。
最新研究表明, GS同工酶活性的调控在RNA转录、蛋白质翻译及翻译后修饰等不同水平上实现[18-19], 且某些转录因子也可以对GS的活性造成影响[20]。氮素利用效率是作物的一个复杂的数量性状, 还需要其他调控的参与, 如外界环境条件、激素调控等, 这些都是下一步研究工作的方向和重点。
4 结论
氮低效烤烟品种成熟期GS同工酶活性在叶肉和叶脉中均较低, 氮素营养物质降解速度快且转移再利用能力弱, NH4+向质外体转移能力强, 从而以氨气大量挥发的形式实现氮素调亏。而氮高效品种与其恰好相反, 生育后期在高氮条件下容易贪青晚熟。叶片GS同工酶活性最终决定了不同烤烟品种的氮素代谢能力和衰老特性。本研究对于解释烤烟品种氮代谢差异以及成熟期氮低效品种的选育具有一定的指导作用。
[1] 史宏志, 韩锦峰. 烤烟碳氮代谢几个问题的探讨. 烟草科技, 1998, 129(2): 34–36. Shi H Z, Han J F. Discussion on several problems of carbon and nitrogen metabolism in flue-cured tobacco., 1998, 129(2): 34–36 (in Chinese).
[2] 严小龙, 张福锁. 植物营养遗传. 北京: 中国农业出版社, 1997. pp 78–106. Yan X L, Zhang F S. Genetics of Plant Nutrition. Beijing: China Agriculture Press, 1997. pp 78–106 (in Chinese).
[3] Hirel B, Andrieu B, Renard S, Quillere I, Chelle M, Pommel B, Fournier C. Physiology of maize II: Identification of physiological markers representative of the nitrogen status of maize () leaves during grain filling., 2005, 124: 178–188.
[4] Fei H, Chaillou S, Hirel B, Polowick P, Mahon J, Vessey J. Effects of the overexpression of a soybean cytosolic glutamine synthetase gene () linked to organ-specific promoters on growth and nitrogen accumulation of pea plants supplied with ammonium., 2006, 44: 543–550.
[5] Li X P, Zhao X Q, He X, Zhao G Y, Li B, Liu D C, Zhang A M, Zhang X Y, Tong Y P, Li Z S. Haplotype analysis of the genes encoding glutamine synthetase plastic isoforms and their association with nitrogen-use and yield-related traits in bread wheat., 2011, 189: 449–458.
[6] 邓扬悟. 甜瓜谷氨酰胺合成酶基因的克隆和功能研究. 上海交通大学博士学位论文, 上海, 2010. Deng Y W. Cloning and Characterization of Glutamine Synthetase Genes in Melon. PhD Dissertation of Shanghai JiaoTong University, Shanghai, China, 2010 (in Chinese with English abstract).
[7] 卢素萍, 李玉娥, 刘鹏飞, 赵铭钦, 张渝婕, 李志鹏. 氮用量对南阳不同基因型烤烟烟叶品质的影响. 江西农业大学学报, 2016, 38: 67–73. Lu S P, Li Y E, Liu P F, Zhao M Q, Zhang Y J, Li Z P. Effects of nitrogen fertilizer on quality of different genotypes of flue-cured tobacco in Nanyang county., 2016, 38: 67–73 (in Chinese with English abstract).
[8] 崔昌范, 王书凯, 宋立君. 烤烟新品种K326和NC89特征特性及配套栽培技术规范. 延边大学农学学报, 1999, 21(1): 70–75. Cui C F, Wang S K, Song L J. Characteristics of flue-cured tobacco new varieties K326 and NC89 and their cultivation techniques., 1999, 21(1): 70–75 (in Chinese).
[9] 武云杰, 李飞, 杨铁钊, 张小全. 氮素营养水平对衰老期烟叶氮代谢的影响及品种间差异. 中国烟草学报, 2014, 20(4): 41–47. Wu Y J, Li F, Yang T Z, Zhang X Q. Effects of nitrogen nutrition on nitrogen metabolism in flue-cured tobacco leaves and differences among varieties., 2014, 20(4): 41–47 (in Chinese with English abstract).
[10] 黄培堂. 分子克隆实验指南. 北京: 化学工业出版社, 2007. pp 155–157. Huang P T. The Condensed Protocols from Molecular Cloning: a Laboratory Manual. Beijing: Chemical Industry Press, 2007. pp 155–157 (in Chinese).
[11] Wang X, Wei Y, Shi L, Ma X, Theg S. New isoforms and assembly of glutamine synthetase in the leaf of wheat (L.)., 2015, 66: 6827–6834.
[12] 邹琦. 植物生理学实验指导. 北京: 中国农业出版社, 2000. pp 56–89. Zou Q. The Guidance of Plant Physiology Experiment. Beijing: China Agriculture Press, 2000. pp 56–89 (in Chinese).
[13] 段旺军, 杨铁钊, 刘化冰, 张小全, 戴亚, 李东亮. 烟叶氨气补偿点的品种间差异及其与氮素代谢的关系研究. 植物营养与肥料学报, 2011, 17: 419–424. Duan W J, Yang T Z, Liu H B, Zhang X Q, Dai Y, Li D L. Differences in NH3compensation point among tobacco (L.) cultivars and its relationship with nitrogen metabolism., 2011, 17: 419–424 (in Chinese with English abstract).
[14] 武云杰, 杨铁钊, 张小全, 李飞, 李丽华. 不同烤烟品种烟叶衰老期氨气挥发及其与氮素代谢的相关性. 中国农业科学, 2013, 46: 4027–4034. Wu Y J, Yang T Z, Zhang X Q, Li F, Li L H. Study on correlation between ammonia volatilization and nitrogen metabolism during the tobacco leaves senescence., 2013, 46: 4027–4034 (in Chinese with English abstract).
[15] Nielsen K H, Schjoerring J K. Regulation of apoplastic NH4+concentration in leaves of oilseed rape., 1998, 118: 1361–1368.
[16] 范江, 杨继洪, 王守旗, 肖志新, 梁梅, 李海平, 邱尧. 打顶对烤烟生长和各器官烟碱积累分配的影响. 作物研究, 2016, 30(1): 36–40. Fan J, Yang J H, Wang S Q, Xiao Z X, Liang M, Li H P, Qiu Y. Effect of topping on growth of flue-cured tobacco and nicotine accumulation and distribution in different organs., 2016, 30(1): 36–40 (in Chinese with English abstract).
[17] 王云华, 欧吉权, 王志强, 周忠新, 欧阳敏, 张楚富. 黄瓜(L.)子叶发育过程中谷氨酰胺合成酶同工酶的变化. 武汉大学学报(理学版), 2005, 51: 517–520. Wang Y H, Ou J Q, Wang Z Q, Zhou Z X, Ou Y M, Zhang C F. Changes of glutamine synthetase isozymes during the development of cucumber (L.) cotyledons.(Nat Sci Edn), 2005, 51: 517–520(in Chinese with English abstract).
[18] Rueda-López M, Crespillo R, Cánovas F M, Avila C. Differential regulation of two glutamine synthetase genes by a single Dof transcription factor., 2008, 56: 73–85.
[19] Stanulović V S, Garcia R M, Labruyère W T, Ruijter J M, Hakvoort T B M, Lamers W H. The 3'-UTR of the glutamine-synthetase gene interacts specifically with upstream regulatory elements, contains mRNA-instbility elements and is involved in glutamine sensing., 2006, 88: 1255–1264.
[20] Kumar R, Taware R, Gaur V S, Guru S K, Kumar A. Influence of nitrogen on the expression of TaDof1 transcription factor in wheat and its relationship with photo synthetic and ammonium assimilating efficiency., 2009, 36: 2209–2216.
Relationship between GS isoenzyme activity and nitrogen transportation in flue-cured tobacco leaves
ZHOU Jian-Fei1, WU Yun-Jie1, XUE Gang1, ZHANG An-Qian1, TIAN Pei1, PENG Yu-Fu2, and YANG Tie-Zhao1,*
1College of Tobacco Science, Henan Agricultural University, Zhengzhou 450002, Henan, China;2Technical Center, Henan Tobacco Industry Co., Ltd., Zhengzhou 450000, Henan, China
A pot experiment was conducted to study the dynamic changes of glutamine synthetase isoenzyme activity and the related physiological indexes in leaves of flue-cured tobacco with different nitrogen efficiencies at maturity stage, and understand the physiological and biochemical mechanism of nitrogen metabolism in flue-cured tobacco leaves. Three different flue-cured tobacco varieties Zhongyan 100, K326, and NC89 with different nitrogen efficiencies were used as materials, and the 12th leaves were taken at different leaf ages. Western blot method was used to determine the content of glutamine synthetase isoenzyme isoforms in leaf mesophyll and main veins. At the same time, the concentrations of NH4+, total nitrogen, apoplastic NH4+and ammonia volatilization were determined. The results show, GS2 was the main form of GS isoenzyme in mesophyll of flue-cured tobacco leaves, the protein subunit content gradually decreased with the age of the leaf. The GS1 isoenzyme dominated in the veins, and its protein subunits showed a trend of increasing first and then decreasing. From 45 to 65 days of leaf age, the GS isoenzyme activities of leaf mesophyll and main vein were all expressed as NC89 > K326 > Zhongyan 100, and the difference among varieties was significant. The activities of GS1 in mesophyll and main veins were negatively correlated with total nitrogen and leaf ammonium concentrations, and positively correlated with apoplastic ammonium concentration and ammonia volatilization. The GS2 activity in mesophyll was positively correlated with total nitrogen and leaf ammonium, and negatively correlated with apoplastic ammonium concentration and ammonia volatilization. The GS2 activity of leaf veins was only significantly correlated with the total nitrogen and ammonia volatilization. The activity of two glutamine synthetase isoenzymes in leaves of nitrogen-inefficient flue-cured tobacco varieties at the mature period was lower, and the ability of nitrogen transfer and reuse was poor, resulting in most of the nitrogen absorbed by plants evaporated in the form of ammonia, and the leaves had a faster rate of aging. However, nitrogen efficient varieties had strong nitrogen reutilization capacity, so the ammonia volatilization was small and the aging was postponed.
flue-cured tobacco;glutamine synthetase isozyme; nitrogen metabolism; ammonia volatilization
2018-04-12;
2018-08-20;
2018-09-14.
10.3724/SP.J.1006.2019.84054
通信作者(Corresponding author): 杨铁钊, E-mail: yangtiezhao@126.com
E-mail: ndzhoujianfei@163.com
本研究由河南中烟工业有限责任公司科技项目(HNZY102015004)和河南省烟草公司科技项目(2018410000270035)资助。
This study was supported by the Henan Tobacco Industry Co., Ltd.Science and Technology Project (HNZY102015004) and Henan Tobacco Company Science and Technology Project (2018410000270035).
URL:http://kns.cnki.net/kcms/detail/11.1809.S.20180912.1609.004.html
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
遗传(2023年8期)2023-08-25 06:59:00
蔬菜(2022年12期)2022-01-01 11:32:10
宁夏医学杂志(2020年3期)2021-01-21 08:23:38
种业导刊(2019年9期)2019-10-21 08:36:38
农村百事通(2019年1期)2019-01-30 13:36:34
农民致富之友(2017年7期)2017-04-27 19:17:28
农民致富之友(2017年2期)2017-03-23 23:45:57
农民致富之友(2015年16期)2015-05-30 21:11:00
农家顾问(2014年11期)2014-12-01 13:29:34
