
转mapk双链RNA 表达载体黄瓜对土壤线虫多样性的影响
2019-12-13 10:48赵晓曼赵松宇孙婷婷田雪亮弭宝彬
中国蔬菜 2019年12期
赵晓曼 赵松宇 孙婷婷 田雪亮* 弭宝彬
(1 河南科技学院资源与环境学院,河南新乡 453003;2 湖南省农业科学院蔬菜研究所,湖南长沙 410125)
据统计,1996 年全球转基因农作物种植面积为170 万hm2,到2000 年增至4 420 万hm2,2016年全球转基因作物种植面积就达1.851 亿hm2,比1996 年增加了110 倍。2016 年耐除草剂作物种植面积为8 650 万hm2,约占全球转基因作物种植面积的47%,抗虫转基因作物约占12%(Clive,2016)。随着转基因作物大面积种植,人们对转基因作物的农业生态环境和人类健康的潜在安全性问题日益关注(刘迎哲,2017;黄锦华 等,2018),开展了众多研究。目前,大量研究集中在转Bt 蛋白基因作物的环境安全性,也就是转基因作物对天敌昆虫和土壤微生物的影响,如细菌、真菌、土壤线虫等(陈彦君,2017;梁晋刚和张秀杰,2017;邱良妙 等,2018;周霞 等,2018)。土壤线虫种类多样,数量丰富,广泛分布于各类土壤中,是土壤生物中十分重要的类群。据估计目前已知土壤线虫数量占地球线虫总数量的35%(Andrassy,1992),它们在土壤生态系统中占有多个营养级,与其他土壤生物构成食物网,对土壤物质循环和能量流动起着重要的作用,因此土壤线虫在指示土壤食物网总体情况及土壤生态功能方面具备较强的优势(Bongers,1990),在评价转基因作物环境安全性方面也被广泛应用。Griffiths 等(2005)研究发现,转Bt基因玉米土壤线虫数量与亲本玉米相比表现出显著的短暂降低。李修强等(2012)研究发现,转Bt基因水稻土壤线虫数量与亲本相比无明显差异。风春等(2015)研究发现,转Bt基因棉花对土壤线虫群落结构和种类组成的影响不显著。上述研究主要针对转Bt基因作物对土壤线虫的安全性进行评价,而对其他类型的转基因作物安全性的研究较少。
促分裂原活化蛋白激酶(mitogen-activated protein,MAPK)广泛存在于真核生物中,是一类高度保守的信号转导路径,在病原物诱导的抗病反应中发挥着重要的作用(马曼莉,2018)。根结线虫是严重危害番茄、黄瓜等蔬菜作物的土传性病害,其防治方法包括抗性品种利用、化学防控、物理防控及生物防控等。转mapk双链RNA表达载体黄瓜能够抑制根结线虫对黄瓜根系的侵染,对土壤真菌、古菌、细菌未产生显著影响(陈国华 等,2013;弭宝彬 等,2013;弭宝彬,2013)。但转mapk双链RNA 表达载体作用靶标为根结线虫的MAPK 激酶,而土壤线虫也存在类似基因,转mapk基因是否对土壤线虫具有影响有待研究。
鉴于此,本试验以转mapk双链RNA 表达载体黄瓜为对象,分析转基因黄瓜对土壤线虫多样性的影响,以期为评价转基因黄瓜的生态安全性奠定基础。
1 材料与方法
1.1 试验地点及供试材料
试验田位于北京顺义温室基地。转基因黄瓜材料来自中国农业科学院蔬菜花卉研究所,含有Mi mpk1基因片段的双链RNA 表达载体。该基因片段来自南方根结线虫,GenBank 登录号为DQ923592。以非转基因黄瓜(Cucumis sativusL.)材料(品种代号9930)为对照。转基因黄瓜和非转基因黄瓜材料在同一温室连续种植3 a(2015~2018年),每年种植两茬,春茬黄瓜生长期为4~6 月;秋茬黄瓜生长期为8~10 月。转基因黄瓜和非转基因黄瓜材料按垄间隔种植,各材料种植50 垄,每垄40 株,两种材料的种植面积分别为200 m2。每种材料3 次重复。黄瓜生长期间按正常农事操作管理。为了防止转基因材料扩散,温室设置防虫网,专人管理。
1.2 土壤取样方法
于第3 年6 月中旬取黄瓜根际土壤样品,转基因黄瓜和非转基因黄瓜材料分别选5 垄作为5 个取样点。每个取样点选择10 株黄瓜,用土壤采样器采集距离根部5 cm 处土壤,取样深度 0~15 cm。10 株黄瓜根际土壤混入一个密封袋,作为一个样点的土壤样品。每个材料共计取5 个样点,即5 次重复,带回实验室,分离土壤线虫。
1.3 土壤线虫收集
土壤线虫收集采用浅盘法:将塑料筐放入平底托盘,单层面巾纸平铺于塑料筐底。称取200 g 土壤样品置于面巾纸上,在平底托盘中加入自来水,确保水面浸没土壤,静置过夜16 h 以上,然后收集平底托盘水中的土壤线虫,离心弃上清液,富集土壤线虫后置于4 ℃冰箱保存。
1.4 土壤线虫DNA 的提取
为了排除土壤腐殖酸对下游试验的影响,采用土壤微生物DNA 提取试剂盒(MP Biomedicals FastDNA kit,American)提取土壤线虫DNA,具体流程参考说明书。提取的土壤线虫DNA 经检测合格后,保存于-20 ℃ 冰箱。
1.5 线虫18S rDNA 基因克隆文库构建
以18S rDNA 基因片段作为土壤线虫的marker 基因,引物为MN18F(5′-CGCG AATRGCTCATTACAACAGC-3′)和22R(5′-GCCT GCTGCCTTCCTTGGA-3′)(Waite et al.,2003),目的片段约900 bp。PCR 反应体系25 μL:1.5 μL DNA 模板,1 μmol·L-1上下游引物,0.5 mmol·L-1dNTPs,2 μL EasyTaqDNA 聚合酶、1×反应缓冲液,20 μL ddH2O。PCR 程序:95 ℃预变性4 min;95 ℃ 变性30 s,58 ℃退火30 s,72 ℃延伸1 min,30 个循环;72 ℃ 延伸5 min。PCR 产物经电泳检测,纯化,连接入pEASY-T5 载体(北京百迈克生物技术有限公司),转化Trans5α 感受态细胞(北京百迈克生物技术有限公司),涂布LB 平板(氨苄青霉素50 μg·mL-1),过夜后随机挑取200 个阳性克隆培养于LB 液体培养基(氨苄青霉素50 μg·mL-1)中振荡培养8 h,将克隆送北京六合华大基因科技有限公司测序。转基因黄瓜和非转基因黄瓜根际土壤线虫各构建3 个文库。
1.6 数据分析
采用Chimera Check 软件对土壤线虫18S rDNA 基因序列进行过滤去杂,去除嵌合体序列。用DOTUR 软件包对土壤线虫18S rDNA 序列划分操作分类单元(operational taxonomic unit,OTU),设定阈值为97%。将每个OTU 代表序列在NCBI进行BLAST 比对,鉴定土壤线虫的物种信息。依据各线虫比例,计算Shannon 和Simpson 指数(海棠 等,2008)。
Shannon 指数H=-∑PilnPi
Simpson 指数D=1-∑Pi2
Pi是样本中第i个线虫分类单元个体数占土壤线虫分类单元总体数量的比例。土壤线虫种类数目是描述土壤线虫群落的重要指标,文库OTU 数目(S)代表土壤线虫种类数目。
为了分析转基因黄瓜和非转基因黄瓜根际土壤线虫群落的差异,采用SPSS 软件对Shannon、Simpson 指数和S 进行方差分析。
1.7 土壤线虫群落指数分析
根据土壤线虫物种信息划分营养类群:食细菌线虫(bacterivores,Ba)、植食性线虫(plant parasitic nematodes,PP)、食真菌线虫(fungivores,Fu)、杂食-捕食性线虫(omnivores-predatores,Om),将分离得到的每个属线虫赋予c-p 值(colonize-persister),并计算各线虫营养类群相对丰度和群落指数(Shannon et al.,1950;Ferris et al.,2001)。
富集指数EI=100×〔e/(e+b)〕
式中,b包括Ba2 和Fu2 类群,e包括Ba1 和Fu2 类群。
结构指数SI=100×〔s/(s+b)〕
式中,s包括 Ba3-Ba5(c-p 值为3~5 的食细菌线虫比例)、Fu3-Fu5(c-p 值为3~5 的食真菌线虫比例)、Om3-Om5(c-p 值为3~5 的杂食-捕食线虫比例)类群。
线虫通路指数 NCR=NBa/(NBa+NFu)
式中NBa指食细菌性线虫的数量,NFu指食真菌性线虫的数量。
为了分析转基因黄瓜和非转基因黄瓜根际土壤线虫群落的差异,采用SPSS 软件对EI、SI 和NCR 进行方差分析。
2 结果与分析
2.1 土壤线虫类群分析
非转基因黄瓜根际土壤线虫文库分别获得73、75 和78 个有效克隆子。转基因黄瓜根际土壤线虫文库分别获得74、75 和76 个有效克隆子。以97%阈值划分OTU,非转基因黄瓜根际土壤线虫文库分别获得22、23、23 个OTU;转基因黄瓜土壤线虫文库获得23、25、22 个OTU。
在线虫属分类水平上,转基因黄瓜根际土壤获得13 个线虫属,分别为:小杆线虫属(Rhabditis)、头叶线虫属(Cephalobus)、拟丽突线虫属(Acrobelodies)、根结线虫属(Meloidogyne)、原杆线虫属(Protorhabditis)、孢囊线虫属(Heteroder)、矮化线虫属(Tylenchorhycbus)、短体线虫属(Pratylenchus)、茎线虫属(Ditylenehus)、滑刃线虫属(Aphelenchoides)、胼眂拟毛刺线虫属(Paratrichodorus)、棱咽线虫属(Prismatolaimus)、遍宫线虫属(Ecumenicus)。非转基因黄瓜根际土壤获得除原杆线虫属、棱咽线虫属、胼眂拟毛刺线虫属以外的10 个线虫属。转基因黄瓜和非转基因黄瓜根际土壤线虫优势属都为:小杆线虫属、头叶线虫属、拟丽突线虫属。小杆线虫属线虫在转基因黄瓜和非转基因黄瓜根际土壤中所占比例较高且相近,分别为44.0%和36.0%。头叶线虫属在转基因黄瓜和非转基因黄瓜根际土壤中占第2 位,分别为22.9%和23.3%。拟丽突线虫属在转基因黄瓜和非转基因黄瓜根际土壤中所占比例分别为8.6%和9.6%(图1-A 和1-B)。
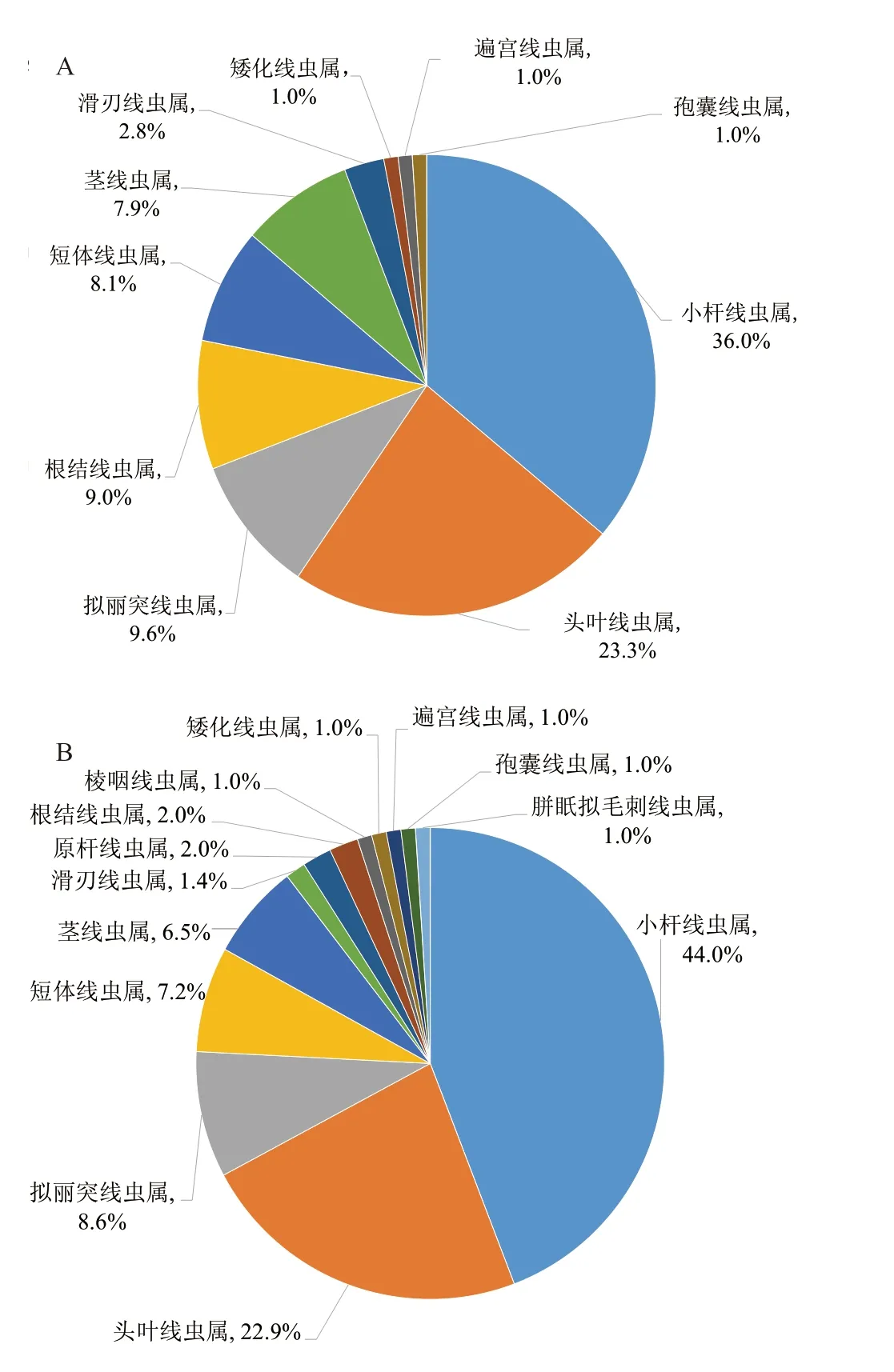
图1 转基因黄瓜和非转基因黄瓜根际土壤线虫各属比例
2.2 土壤线虫营养类群分析
转基因黄瓜和非转基因黄瓜根际土壤中食细菌线虫为优势营养类群,所占比例分别为77.1%和73.2%,无显著差别(图2)。植食性线虫居第二位,所占比例分别为20.0%和25.4%差异显著(P=0.003),其中非转基因黄瓜根际土壤中植食性线虫比例高于转基因黄瓜,主要源于非转基因黄瓜根际土壤中根结线虫数量较多,这也证实转基因黄瓜对根结线虫具有一定的抑制作用。食真菌线虫和捕食性线虫在转基因黄瓜和非转基因黄瓜根际土壤中均为稀有类群,所占比例较小。

图2 转基因黄瓜和非转基因黄瓜根际土壤线虫营养类群
2.3 土壤线虫多样性指数分析
从多样性指数来看(表1),转基因黄瓜和非转基因黄瓜根际土壤线虫的Shannon 指数、Simpson 指数以及土壤线虫的物种数目(S)均与非转基因黄瓜无显著差异。总体来看,转基因黄瓜土壤线虫的多样指数与非转基因黄瓜无显著差异。
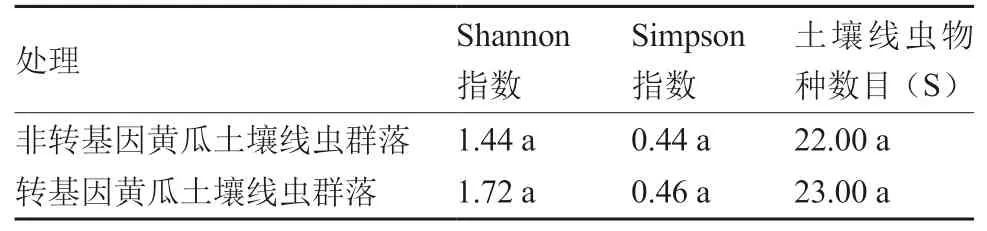
表1 转基因黄瓜和非转基因黄瓜根际土壤线虫多样性指数
2.4 土壤线虫群落指数分析
从土壤线虫群落指数来看(表2),转基因黄瓜根际土壤线虫的富集指数(EI)、结构指数(SI)以及土壤线虫的线虫通路指数(NCR)与转非基因黄瓜均无显著差异。
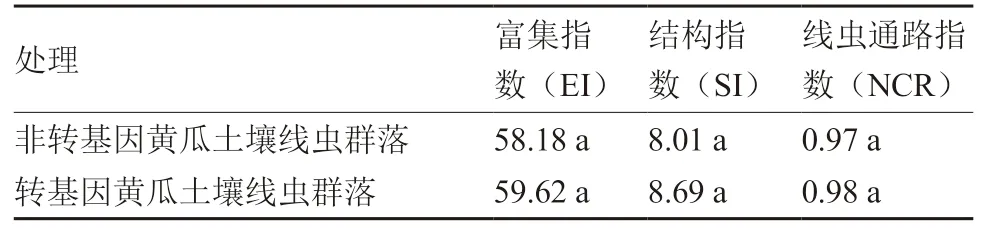
表2 转基因黄瓜和非转基因黄瓜根际土壤线虫生态指数
3 结论与讨论
本试验发现,转mapk基因黄瓜根际土壤线虫优势类群与非转基因黄瓜基本一致,均为小杆线虫属、头叶线虫属和拟丽突线虫属。研究表明,小杆线虫属线虫对土壤环境适应性强,生活史短,世代交替快,繁殖迅速,因此种群数量多,在各类型土壤中均为优势类群,这与本试验结果一致(曾四满,2016;赵婉婷 等,2019)。孔云等(2018)研究发现玉米田土壤中头叶线虫属线虫为优势类群,也表明该属线虫具有较强的适应能力,与本试验结果较为一致。拟丽突线虫属线虫具有很宽的生态幅(12~35 ℃),对温度要求较低,环境适应能力较强(Anderson &Coleman,1982;Bakonyi &Nagy,2000;郭佳惠 等,2018)。本试验中拟丽突线虫属线虫的比例相对较高,也表明该线虫适应大棚土壤温度环境。转基因黄瓜和非转基因黄瓜根际土壤线虫的线虫通路指数均较高,表明温室土壤有机质分解过程以细菌通道为主,细菌丰度高,为食细菌线虫提供丰富食物来源,这也是食细菌线虫成为优势类群的主要原因。
从土壤线虫多样性指数和生态指数来看,转基因黄瓜和非转基因黄瓜根际土壤线虫群落的Shannon、Simpson、EI、SI 和NCR 指数无显著差异,表明转mapk基因黄瓜并未对根际土壤线虫产生显著的影响,其原因可能有两方面:一方面,土壤线虫包括食细菌线虫、食真菌线虫和杂食-捕食性线虫,主要取食土壤细菌、真菌或其他线虫,不取食植物。因此,转基因黄瓜表达的mapk双链RNA 不会对这几类线虫产生直接作用(闫小梅,2015)。另一方面,转基因黄瓜表达的mapk双链RNA 随根系分泌物进入土壤,可能被土壤中的RNA 酶所降解,失去生物活性(王艳艳 等,2015),不能发挥对土壤线虫的抑制作用。此外,本试验发现土壤线虫中也有一定数量的根结线虫,并且根结线虫在转基因黄瓜根际土壤线虫中所占比例较低,表明转基因黄瓜对根结线虫有一定的抑制作用。本试验中选用的转基因黄瓜植株能够表达的mapk双链 RNA,对侵入黄瓜根内的根结线虫进行RNA 干扰,从而杀死根结线虫(陈国华,2008)。
总体来看,转mapk双链RNA 表达载体黄瓜并未对土壤线虫多样性有显著影响,但后续需继续观察长期的累积效应,其对根际土壤真核生物群落的影响还需深入研讨。
猜你喜欢
今日农业(2022年14期)2022-09-15
浙江农业科学(2022年7期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
四川农业科技(2021年7期)2021-12-01
农业科技与信息(2020年14期)2020-12-18
中国果业信息(2020年10期)2020-12-15
西北农业学报(2020年12期)2020-12-14
绿色科技(2019年14期)2019-11-19
中国森林病虫(2019年5期)2019-10-11
江苏农业科学(2019年5期)2019-09-02
