
基于高通量测序技术研究老年人肠道菌群结构变化*
2019-11-22 03:26王思平
胃肠病学 2019年9期
余 莉 李 红 王思平
中国人民解放军总医院健康管理研究院国宾二科(100853)
背景:随着我国人口的老龄化,老年人的健康保健备受关注。目的:探讨增龄对肠道菌群结构的影响,以期通过调节肠道微生态提升老年人健康水平。方法:收集30例健康老年人(60~80岁)和10例健康青年人(18~40岁)的粪便样本,采用高通量测序技术测定所有粪便细菌的16S rRNA基因V3、V4区序列,使用MiSeq 16S宏基因组APP和Qimme软件分析菌群物种分类、物种多样性指数、样本组间显著性差异和各组生物学标志物。结果:老年健康组与青年健康组肠道菌群OTU均数接近,物种多样性相似(P>0.05)。Metastats分析显示,在菌门水平上,两组间优势菌门拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)差异均无统计学意义(P>0.05)。在菌属水平上,两组间共有20余种菌属差异有统计学意义(P<0.05)。老年健康组八叠球菌属(Sarcina)、厌氧支原体属(Anaeroplasma)、甲基杆菌属(Methylobacterium)、丛毛单胞菌属(Comamonas)、Olsenella、Rheinheimera、Terrisporobacter、食酸菌属(Acidovorax)、红蝽菌科(Coriobacteriaceae_UCG-003)、粪芽孢菌属(Coprobacillus)、Gelria、Aquabacterium、氨基酸球菌属(Acidaminococcus)、Caldicoprobacter、肠杆菌属(Enterorhabdus)、Turicibacter丰度显著增高,青年健康组厌氧乙酸属(Acetanaerobacterium)、漫游球菌属(Vagococcus)、瘤胃菌科(Ruminococcaceae_UCG-009)、草酸杆菌属(Oxalobacter)、粪球菌属_2(Coprococcus_2)丰度显著增高。LEfSe分析显示,老年健康组生物学标志物富集于食酸菌属、缠结优杆菌(Eubacterium_nodatum_group)、梭菌科_1(Clostridiaceae_1)、丛毛单胞菌科(Comamonadaceae),青年健康组则富集于普氏菌属_2(Prevotella_2)、罗氏菌属(Roseburia)、厌氧乙酸属。结论:随着年龄的增长,肠道菌群结构和功能发生一定程度的改变。老年人产丁酸盐细菌明显减少,同时机会致病菌增多。
近年来,随着分子生物学的发展,被誉为人类“第二基因组”的肠道菌群成为研究热点。越来越多的研究表明,肠道菌群结构和功能的改变与代谢性疾病、自身免疫性疾病、过敏性疾病、精神疾病、神经退行性疾病、消化道肿瘤等的发生、发展之间存在关联。影响肠道菌群结构的因素主要有性别、年龄、宿主遗传背景、环境、食物、抗菌药物等[1]。我国是世界上60岁以上老年人最多的国家[2],据国家统计局年度调查(2011)和《中国老龄事业发展报告(2013)》,我国已全面进入人口老龄化社会,老年人的健康保健和医疗成为全社会关注的焦点,预防衰老相关疾病,改善老年人健康是亟待解决的社会问题。本研究主要探讨增龄对肠道菌群结构的影响,以期通过调节肠道微生态为提升老年人健康水平提供新思路。
对象与方法
一、研究对象
连续收集2016年1月—2018年1月在中国人民解放军总医院健康管理研究院进行健康体检、检查项目包括常规结肠镜检查的个体。纳入标准:①年龄18~40岁或60~80岁;②结肠镜检查结果提示未见异常;③体力状况ECOG评分0~2,依从性良好;④签署知情同意书。排除标准:①肿瘤和放化疗患者;②近1个月服用过微生态制剂(益生菌、益生元、合生元)、深海鱼油制剂(omega-3)、抗氧化剂(维生素A、C、E);③肝肾功能异常和胰腺疾病;④自身免疫性疾病,近3个月服用过非甾体消炎药、免疫抑制剂、糖皮质激素;⑤严重心脑血管疾病、糖尿病、高脂血症;⑥肥胖(体质指数>28 kg/m2);⑦活动性炎症性肠病、结直肠息肉;⑧近1个月内感染、使用抗菌药物;⑨近1个月服用过聚乙二醇等导泻剂;⑩慢性便秘、急慢性腹泻。入组患者根据年龄分为老年健康组(60~80岁)和青年健康组(18~40岁)。
二、方法
受检者于结肠镜检查前留取粪便标本,通过筛选纳入研究者以高通量测序技术(FloraCheckTM,苏州艾达康医疗科技有限公司)测定所有粪便细菌的16S rRNA基因V3、V4高可变区序列,使用MiSeq 16S宏基因组APP和Qimme软件分析菌群物种分类、物种多样性指数、样本组间显著性差异和各组生物学标志物,获得老年健康组与青年健康组肠道菌群组成及其活性信息。
1. OTU(operational taxonomic units, 运算分类单元)分析和物种注释:OTU是一个人为设置的分类单元,根据测序结果,将相似性>97%者归为一个OTU。以韦恩图表示两组特有或共有的OTU个数。将总OTU与数据库比对,对物种在不同分类水平上进行注释,以profiling柱状图表示两组物种的相对丰度。
2. α多样性分析:基于OTU分析结果,可反映菌群丰度和多样性,包括chao1、observed-species、PD-whole-tree和shannon四个指数,其中chao1常用于评估物种总数,shannon用于估算样本微生物多样性,本研究应用这两个指数进行统计学分析,P<0.05为差异有统计学意义。
3. β多样性分析:用于比较样本在物种多样性方面的差异。本研究采用主坐标分析(principal coordinates analysis, PCoA)比较样本间菌群组成的差异,以二维坐标图呈现。坐标图上两点距离越近,样本组成越相似;两点距离越远,样本组成相似度越低。
4. 差异性分析:采用Metastats检验对物种组成差异有统计学意义的组别从菌门、菌属水平进行菌群分析,P<0.05为差异有统计学意义。
5. LEfSe分析:LEfSe采用线性判别分析(linear discriminant analysis, LDA)估算每组物种丰度对差异效果影响的大小,找出显著差异性的群落或物种,即宏基因组生物学标志物(biomarker)。
三、统计学分析

结 果
一、一般情况
共40例健康成人纳入研究,其中老年健康组30例,青年健康组10例。老年健康组男性15例,女性15例,平均年龄(67.0±4.6)岁,体质指数(BMI)(22.72±1.69) kg/m2;青年健康组男性6例,女性4例,平均年龄(31.7±5.9)岁,BMI(22.44±2.14) kg/m2。两组间性别(χ2=0.033,P=0.855)和BMI(t=0.429,P=0.670)差异无统计学意义,年龄差异有统计学意义(t=19.519,P=0.001)。
二、生物信息学分析
1. OTU和丰度分析:老年健康组OTU个数为4 791条,青年健康组为3 472条(图1)。根据物种注释结果,各样本物种在不同分类等级中组成柱状图,可见各样本丰度较高的物种及其比例(图2)。
2. α多样性分析:chao1、shannon指数分析图中,老年健康组与青年健康组OTU均数接近,两组物种多样性相似,差异无统计学意义(P>0.05)(图3)。
3. β多样性分析:在属水平上对老年健康组和青年健康组进行PCoA分析,结果显示主成分1和2无法将两组有效区分,主成分1可解释两组差异的8.6%,主成分2可解释两组差异的6.51%(图4)。
4. 老年健康组和青年健康组注释、主成分分布:将总OTU与数据库比对,对物种在不同分类水平上进行注释,作物种profiling柱状图,纵坐标表示相对丰度,不同颜色对应不同物种名称,色块长度表示该物种的相对丰度比例。此步骤主要在门、属水平进行分析。
①门水平:两组样本共包含14个门,所有样本的优势菌门均为拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)。老年健康组丰度最高的4个门依次为:拟杆菌门53.8%,厚壁菌门34.0%,变形菌门9.1%,放线菌门(Actinobacteria) 2.0%;而青年健康组分别为:拟杆菌门63.1%,厚壁菌门32.0%,变形菌门3.4%,放线菌门0.7%(图5)。Metastats分析显示两组间四个菌门差异均无统计学意义(P>0.05)。
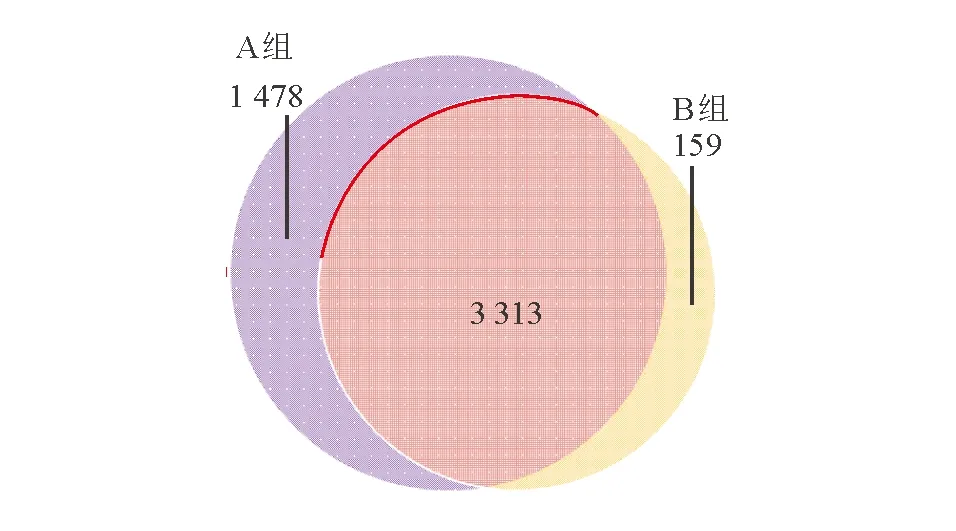
图1 老年健康组(A)和青年健康组(B)基于OTU的韦恩图
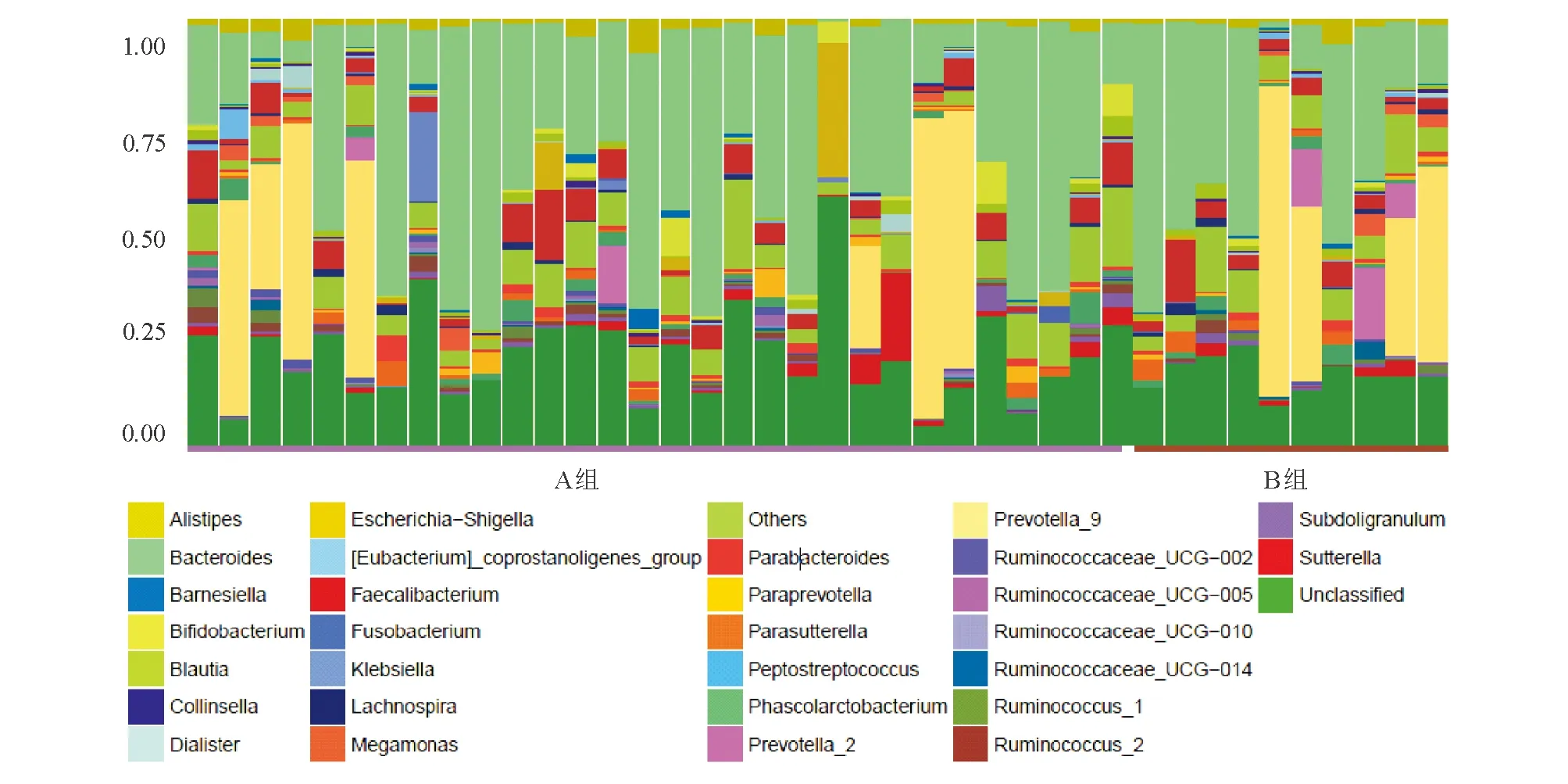
图2 老年健康组(A)和青年健康组(B)各样本在属水平上的物种相对丰度
②属水平:老年健康组优势菌属大都属于拟杆菌门,菌属丰度依次为:拟杆菌属(Bacteroides)34.9%,普氏菌属_9(Prevotella_9)11.3%,粪杆菌属(Faecalibacterium)4.5%,埃希菌属(Escherichia-Shigella)1.7%,萨特菌属(Sutterella)1.7%,Alistipes1.5%;青年健康组优势菌属同样大都属于拟杆菌门,菌属丰度依次为:拟杆菌属32.6%,普氏菌属_9 19.1%,粪杆菌属4.4%,普氏菌属_2 3.8%,考拉杆菌属(Phascolarctobacterium)1.7%,Parasutterella1.7%,Alistipes1.4%(图6)。Metastats分析显示,在菌属水平上,两组间共有20余种菌属差异有统计学意义(P<0.05)。老年健康组八叠球菌属(Sarcina)、厌氧支原体属(Anaeroplasma)、甲基杆菌属(Methylobacterium)、丛毛单胞菌属(Comamonas)、Olsenella、Rheinheimera、Terrisporobacter、食酸菌属(Acidovorax)、红蝽菌科(Coriobacteriaceae_UCG-003)、粪芽孢菌属(Coprobacillus)、Gelria、Aqua-bacterium、氨基酸球菌属(Acidaminococcus)、Caldico-probacter、肠杆菌属(Enterorhabdus)、Turicibacter丰度显著增高,青年健康组厌氧乙酸属(Acetanaero-bacterium)、漫游球菌属(Vagococcus)、瘤胃菌科(Ruminococcaceae_UCG-009)、草酸杆菌属(Oxalo-bacter)、粪球菌属_2(Coprococcus_2)丰度显著增高。
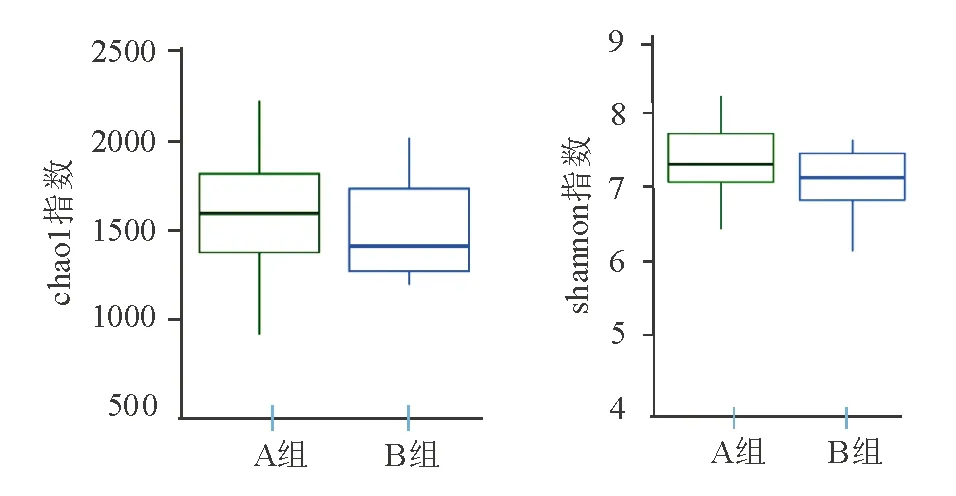
图3 老年健康组(A)和青年健康组(B)α多样性分析
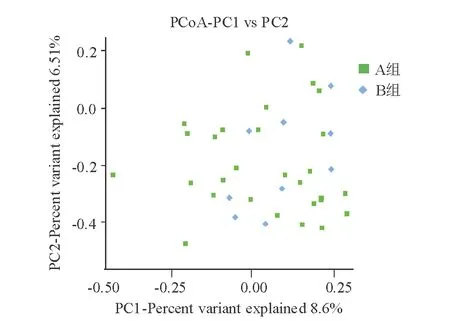
图4 老年健康组(A)和青年健康组(B)β多样性分析

图5 老年健康组(A)和青年健康组(B)样本菌群在门水平上的主成分
5. LEfSe分析:以LEfSe分析寻找有显著差异性的菌属,即宏基因组生物学标志物,结果显示老年健康组生物学标志物富集于食酸菌属、厚壁菌门的缠结优杆菌(Eubacterium_nodatum_group)和梭菌科_1(Clostridiaceae_1)、变形杆菌门的丛毛单胞菌科(Comamonadaceae);青年健康组生物学标志物富集于普氏菌属_2、罗氏菌属(Roseburia)、 厌氧乙酸属(图7)。
讨 论
2010年和2011年,Biagi等[3]和Claesson等[4]分别发表了关于意大利和爱尔兰老年人肠道菌群的研究,是衰老肠道菌群研究领域中两个标志性、引领性的研究。人体肠道菌群直至2~3岁才近似成人并趋于稳定。影响肠道菌群的因素包括遗传、种族、年龄、生活和饮食习惯、环境等,其中宿主遗传背景对肠道菌群结构的贡献率仅为1.9%,环境因素在肠道菌群的形成过程中起决定性作用[5]。老年人饮食结构的改变以及肠道蠕动功能下降,导致肠道屏障功能减弱,肠黏膜通透性增加,肠道微生态发生一定程度的变化。研究发现持续性低度炎症是老龄和各种年龄相关疾病最一致的生物学特征之一[6]。最近证据表明,肠道菌群可能在老龄相关炎症中起关键作用[7]。
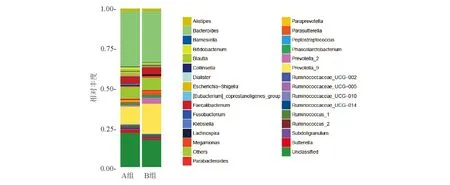
图6 老年健康组(A)和青年健康组(B)样本菌群在属水平上的主成分分布
本研究α多样性分析显示,老年健康组和青年健康组的OTU均数接近,两组物种多样性相似;β多样性分析显示,两组物种主要成分组成接近。在物种分类水平上,拟杆菌门和厚壁菌门是肠道菌群的主要组成成分,本研究中两组均为拟杆菌门占比最高,厚壁菌门/拟杆菌门比值在老年健康组和青年健康组分别为0.63和0.51,组间拟杆菌门和厚壁菌门丰度均无明显差异。上述结果与既往对老年肠道菌群的研究结果有所不同。Odamaki等[8]的研究发现,肠道菌群丰度和多样性随年龄增长而降低,老年人群拟杆菌门和变形菌门丰度随年龄增长而增加,厚壁菌门丰度则随年龄增长呈降低趋势。而Bian等[9]对中国健康人群的横断面研究显示,老年健康人群的肠道总体微生物组成与青年健康人群相似,差异主要存在于与<20岁的人群比较时。各研究结果不同的原因可能在于:①不同国家、地区的研究对象,在种族、生活和饮食习惯等方面存在差异。②研究采用的测序方法不同。分子生物学技术常用的测序方法包括高通量测序、实时荧光定量PCR和PCR-梯度凝胶电泳,不同方法各有其优势和局限性。③对老年人的定义不同,导致不同研究检测人群中老年人的平均年龄存在较大差异。
本研究Metastats分析显示,在菌属水平,产丁酸盐细菌在老年健康组与青年健康组间丰度差异较大,已鉴定出的产生丁酸盐细菌瘤胃球菌属(Ruminococcus)和粪球菌属[10]在青年健康组中明显增多。丁酸盐作为一种短链脂肪酸(SCFAs),是结肠细胞的能量来源之一,并可通过细胞代谢、菌群稳态、抗增殖、免疫调节以及基因/表观遗传调控等机制发挥抗炎和抗肿瘤作用。对丁酸的机制研究表明其可促进结肠细胞凋亡;通过抑制组蛋白脱乙酰基酶调控基因表达,从而调节肠道屏障的完整性,降低肠道通透性;通过cAMP依赖性机制激活糖异生作用,从而有益于机体能量稳态[11-12]。青年健康组草酸杆菌属丰度亦较高,其具有促进体内草酸排泄的作用,可调节宿主草酸稳态、预防草酸钙肾结石形成以及其他草酸相关病理改变[13]。何方等的团队分析了中国西南地区148例健康中老年人的肠道菌群,鉴定出的与年龄呈负相关的菌属中包括草酸杆菌属,同时发现衰老相关特定菌群与免疫老化之间可能存在关联[14]。老年健康组粪芽孢菌属和肠杆菌属数量明显增加。研究发现粪芽孢菌属在慢性功能性便秘患者和结肠癌模型大鼠肠道中的丰度显著增高,提示其可能为潜在致病菌[15-16];而肠杆菌科在肠易激综合征(IBS)患者肠道中的丰度较对照组显著增高[17]。芽孢杆菌属、肠杆菌属等参与了肠道内未消化蛋白的发酵,此种细菌腐败过程可能破坏肠道稳态,氨、腐胺等腐败产物参与了结直肠癌的发生[18]。此外,老年健康组柔膜菌门的厌氧支原体属、红蝽菌科丰度显著增高。在高脂饮食大鼠中,柔膜菌门被证明与肥胖相关代谢参数有关[19]。有研究[20]发现病态肥胖个体肠道菌群中红蝽菌科丰度增高,但在胃旁路术后可恢复至正常体质量个体水平,提示该物种可能具有致代谢紊乱作用。予小鼠饮食中长期添加左旋肉碱,可见盲肠菌群结构发生改变,厌氧支原体属数量增多并与参与动脉粥样硬化形成的三甲胺-N-氧化物合成增加有关[21]。上述发现提示了年龄对肠道菌群结构和功能的影响,老年人肠道中机会致病菌增多。然而本研究未发现既往研究公认的具有促进健康作用的双歧杆菌数量在老年健康组与青年健康组间存在显著差异,与部分作者的研究发现相一致[22]。分析其原因可能与不同研究的受检者来自不同地域、肠道菌群背景不同等因素有关。此外,产丁酸盐粪杆菌属以及与人体健康和代谢相关的Akkermansia在本研究老年健康组与青年健康组间亦无明显差异,进一步提示了肠道菌群与宿主间关系的复杂性,特定微生物的作用应在具体肠道菌群背景之上进行分析。
本研究通过LEfSe分析发现老年健康组食酸菌属、厚壁菌门的缠结优杆菌和梭菌科、变形杆菌门的丛毛单胞菌科富集。缠结优杆菌多见于口腔疾病患者[23]。近年对口腔菌群的研究发现,绝大部分口腔细菌可转移并定植于结肠,尤其是机会致病菌,结直肠癌患者肠道富集菌种中有数个来源于口腔[24]。本研究中老年健康组缠结优杆菌富集在肠道衰老中的作用有待进一步明确。梭菌科是肠道正常优势细菌,既包括有益菌也包括有害菌,有益梭菌主要有拟球梭菌亚群(Clostridiumcoccoidesgroup)和柔嫩梭菌亚群(Clostridiumleptumgroup)两大亚群,而有害梭菌则以艰难梭菌(Clostridiumdifficile)为代表。本研究老年健康组梭菌科富集,具有差异性的粪芽孢菌属数量增多,具有潜在致病可能。食酸菌属对人体肠道功能以及健康和疾病的影响有待进一步研究。青年健康组产丁酸盐罗氏菌属富集,研究[25]证明产丁酸盐罗氏菌属数量减少是溃疡性结肠炎患者肠道微生态失衡的特征之一。一项百岁老人肠道宏基因组学研究[26]显示,与衰老相关的肠道微生物组特征为产SCFAs相关基因缺失、糖类分解能力下降和蛋白水解功能增强。在较年轻的老年人队列中,同样发现其产SCFAs功能降低[27]。上述发现表明青年人可能拥有更健康的肠道微生态。
综上所述,本研究主要探讨了老年人群与青年人群肠道菌群结构的差异,部分差异性菌群的代谢功能及其对宿主影响的作用机制尚需开展更多动物实验加以证实。随着年龄的增长,肠道菌群结构和功能发生一定程度的改变,表现为老年人产丁酸盐细菌明显减少,同时机会致病菌增多。调节和维持肠道微生物健康无疑是一种值得关注和探索的医疗策略,是促进老年人身体健康、改善其生活质量,进而缓解医疗压力的研究方向。我国目前尚无大规模衰老人群的肠道菌群研究,本研究对年龄与肠道菌群结构变化的初步探索可为今后衰老肠道微生态的深入研究提供线索和参考。
猜你喜欢
安徽农业科学(2022年20期)2022-11-11
科学技术与工程(2022年26期)2022-11-01
河南医学研究(2022年19期)2022-10-19
今日农业(2022年14期)2022-09-15
中老年保健(2022年2期)2022-08-24
中国农学通报(2022年14期)2022-06-01
分子诊断与治疗杂志(2022年4期)2022-05-30
油气田环境保护(2022年2期)2022-05-09
中国饲料(2022年5期)2022-04-26
昆明医科大学学报(2022年3期)2022-04-19
