
间歇性大强度运动前后人体血清代谢组学特征
2019-10-11 09:03郭新明吴丽君向欢
中国运动医学杂志 2019年9期
郭新明 吴丽君 向欢
1太原师范学院体育系(太原 030619)
2山西大学体育学院
代谢组学(metabonomics/metabolomics)是自人类基因组计划、蛋白质组计划之后,迅速发展起来的以高通量检测和数据处理为手段、以信息建模与系统整合为目标、以群组指标分析为基础的一门生物学学科[1]。代谢物检测技术包括核磁共振波谱(NMR)和质谱(MS)等,可以不预先设定将要检测的具体指标而“全景式”扫描所有代谢物,从而反映药物、环境污染、运动等外界因素对机体代谢的影响,在此基础上,进一步探索或阐明相关生命过程的代谢网络调控应答机制。上述代谢组学思想与分析技术已广泛应用于生理学、毒理学、功能基因组学、疾病诊断、药物疗效与药物作用机理等方面,并取得了长足进步[2]。
目前,国外代谢组学在体育科学领域的应用主要集中在运动训练方法手段[3-4]、运动生理状态监控[5-6]、运动营养补充[7-8]、运动与疾病治疗[9-10]等方面。近些年,我国学者采用代谢组学方法进行的研究也呈蓬勃发展态势,主要集中在不同运动项目运动员的代谢特征[11-13]、运动对疾病的治疗[14-16]及营养补剂结合运动对机体机能的影响[17-18]等几个方面。随着工作压力增大、生活节奏加快,部分无法保障健康运动方式的健身锻炼者及某些项目的运动员往往采用间歇性大强度运动进行锻炼、训练和比赛,这会对机体产生较大扰动,使机体代谢变化更为剧烈。鉴于此,本研究采用三次大强度间歇3分钟的运动方式,探讨分析运动前后人体血清代谢物的变化状况,以探究间歇性大强度运动机体的代谢特征。
1 对象与方法
1.1 研究对象
根据研究需要,为减少个体差异对实验结果造成不良影响,按照身高、体重、运动等级筛选出体育学院2016级体育教育专业男生8人,年龄20.19±1.14岁,身高1.78±0.08 m,体重69.35±3.54 kg,均为二级运动员,身体健康,无抽烟、喝酒、熬夜等不良生活习惯,无慢性病史及家庭遗传病史。
1.2 实验仪器与试剂
选用瑞典产Monark874E定量负荷功率自行车完成一次间歇性大强度运动。选用 Brucker600-MHz AVACNCEⅢ NMR谱仪(600.13 MHz Proton frequency)、TGL-16高速台式冷冻离心机、XW-80A涡旋混合器、KQ5200E型超声波清洗器、D2O重水、PBS磷酸缓冲液完成血清核磁共振代谢物测定。
1.3 实验设计与运动方案
实验设定为7天。由于代谢组学研究对血样的要求较高,为避免饮食对实验数据的影响,本研究采用空腹运动、空腹采血的方式。实验第1天,8名受试者于清晨8时空腹进行第一次肘静脉采血,标记为E1;采血后立即进行蹬骑功率自行车运动,在运动后即刻、1小时分别采血,标记为E2、E3;实验第2天,受试者清晨8时空腹再次进行肘静脉采血,标记为E4;实验第7天,受试者清晨8时空腹进行最后一次采血,标记为E5。运动方式为受试者全力蹬骑功率车30 s×3/间歇3 min(负荷为0.075 kg/kg体重)的间歇性大强度运动[19]。
1.4 样本采集与处理
每次采集肘静脉血5 mL,分别置于抗凝管内,经3000 r/min离心30 min后,取上清液1 mL置于EP管内,储存于-80℃冰箱保存待测。
1.5 NMRNMR数据采集与处理
样品处理:将血液样品放置于冰水混合物中解冻后,取450 μL置于容量为2 mL的EP管中,加入900 μL甲醇。涡旋2 min使之充分反应,再放入TGL-16高速台式冷冻离心机中4℃、13000 r/min离心30 min。离心后取900 μL上清液置于容量5 mL的EP管中,经氮吹仪空气源吹干后加入600 μL PBS缓冲液在超声清洗器作用下复溶,待充分溶解后取550 μL加入内径为5 mm的核磁管,标记好,4℃保存、待测。
NMR序列设置:将放有样品的核磁管进行1HNMR检测,采用CPMG(Carr-Purcell-Meiboom-Gill)脉冲序列,CPMG脉冲序列的优点是可以压制水峰及大分子物质信号,从而增加样品中小分子物质敏感性。自旋弛豫延迟320 ms,自由感应衰减64 K数据点,谱宽8 KHz,64次扫描。
NMR数据处理:得到样本的核磁信号后,使用MestReNova软 件(Mesterlab Research,Santiagode compostella,Spain)对图谱进行傅立叶转换,再调整基线、校正相位。以TSP(total suspended particulate)的化学位移为基准校正1H-NMR谱图的位移。对0.5~9 ppm的图谱进行等宽度分割,切除4.5到6.2的水峰信号,再分段积分后将积分值保存于Excel中,将数据归一化处理后进行多元统计分析。用SIMCA-P13.0(Umetric,Sweden)对积分值进行中心化与规格化后,对其进行偏最小二乘判别分析(PLS-DA)、正交偏最小二乘判别分析(OPLS-DA),最终分别结合各组间配对样本T检验的P值与VIP值找出差异代谢物。
2 结果
使用Chenomx NMR Suite软件及相关文献对核磁共振代谢图谱加以指认,得到实验对象不同时期0.5~8.5 ppm的1H-NMR血清代谢图谱[20]。
2.1 受试者血清小分子代谢物的1 1H-NMRH-NMR代谢图谱指认
本实验所采集的1H-NMR图谱如图1,时间分别为:运动前(E1)、运动后即刻(E2)、运动后1小时(E3)、运动后1天(E4)、运动后1周(E5)。通过查阅人类代谢组数据库(Human Metabolome Database,HMDB)及相关文献[21-22],共指认出22种代谢标志物:脂质、异亮氨酸、亮氨酸、缬氨酸、乳酸、丙氨酸、醋酸、N-乙酰基-糖蛋白、谷氨酸、谷氨酰胺、柠檬酸、肌酸、肉毒碱、磷酸胆碱、氧化三甲胺、甜菜碱、乙酰乙酸、甘氨酸、甘油、葡萄糖、酪氨酸、组氨酸。
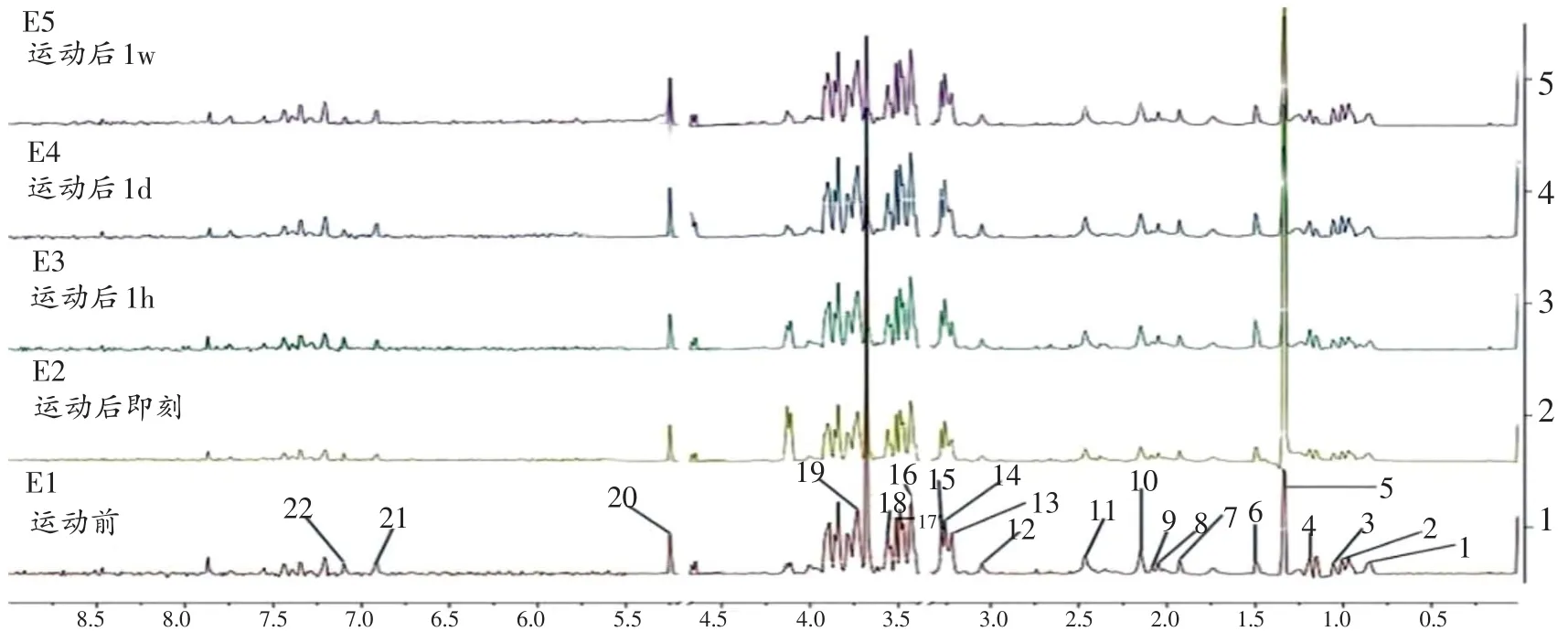
图1 受试者不同时期血清1H-NMR图谱(ppm)
2.2 受试者运动前与运动后不同时刻相比较的血清代谢标志物
将MestRenova软件处理后的数据导入到SIMCAP 13.0软件采用OPLS-DA方法进行分析,得到得分图与S-Plots图。得分图中每一个点代表一个样本,所含物质含量越相同的样本在位置上越接近,因此在图中我们可以看到同一组的样本距离较近,甚至发生重叠,而不同组的样本距离较远,因此可以在图上看到不同组的样本分布在不同区域。S-Plots中每个点代表一种小分子物质,VIP值越大的点(即离原点越远的点)对受试者不同时期的血清样本分离贡献越大。在运动不同阶段的几个R2和Q2分别代表在相应阶段的解释能力和预测能力。经过模型交叉验证,只有模型成立才能证明PLS-DA和OPLS-DA分析结果有意义。
由图2A可知,血清样本可以完全分开,说明人在间歇性大强度运动前和运动后即刻机体小分子物质的含量发生了明显变化。图2B载荷图中每一个点代表一个变量,变量的 VIP值越大,即距离原点越远,说明其对血清样本的分离贡献率越高。图2C表明检验PLS-DA模型验证成立。本研究对载荷图的数据进行筛选,确定找出的代谢标志物其VIP>1且满足与运动前E1之间配对样本t检验双尾检测结果P<0.05两个条件。
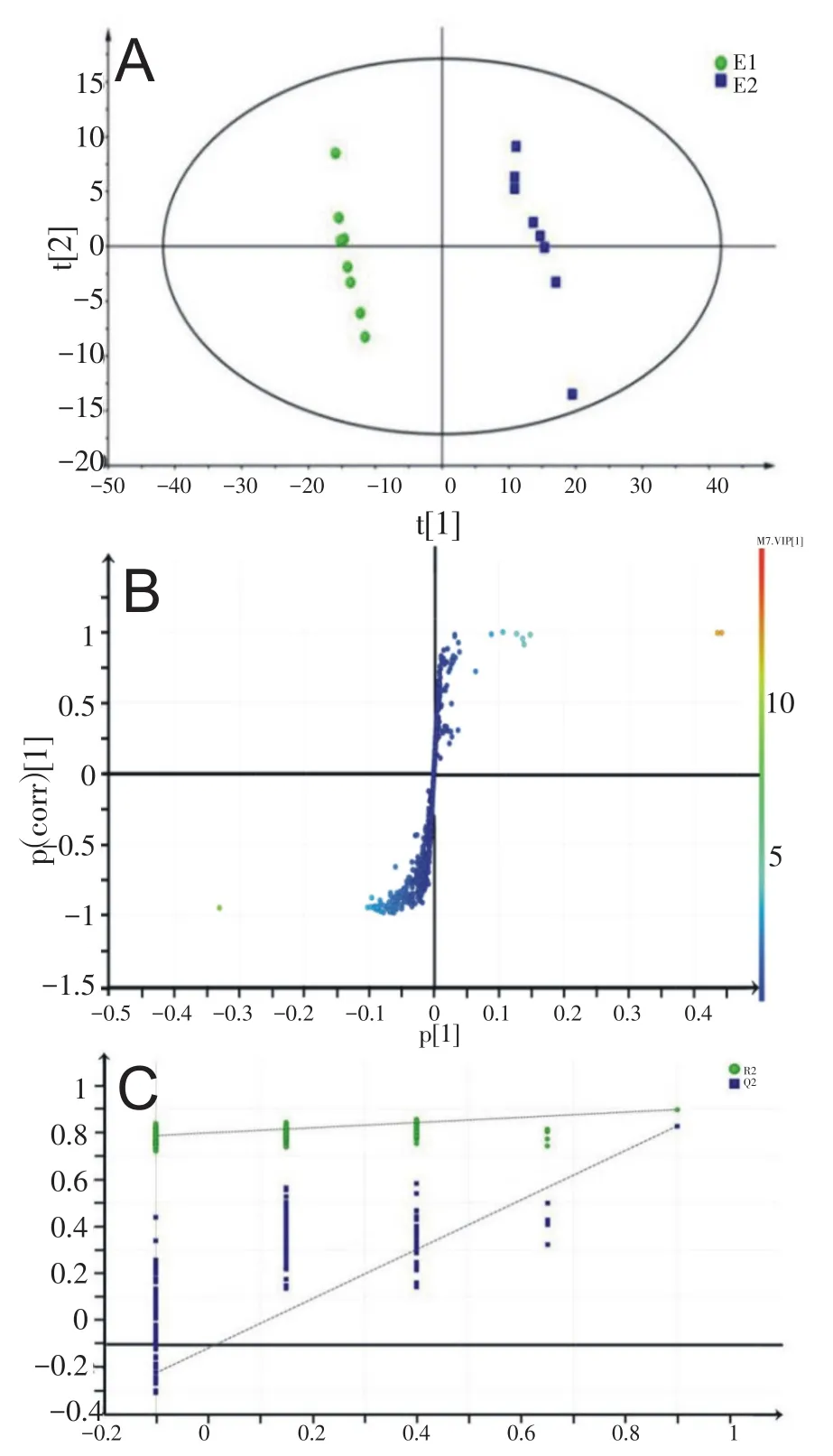
图2 E1与E2受试者血清代谢标志物分析图
由表1可见,将运动前E1与运动后即刻E2检测出的物质进行配对样本t检验,共检测出13种差异代谢物(P<0.01),分别是:亮氨酸、异亮氨酸、缬氨酸、乳酸、谷氨酸、谷氨酰胺、肌酸、肉毒碱、甜菜碱、乙酰乙酸、甘氨酸、甘油、葡萄糖,除乳酸呈上升趋势外,其余物质均呈下降趋势。E1与E3间共检测出8种差异代谢标志物,分别是:亮氨酸、异亮氨酸、缬氨酸、乳酸、甜菜碱、乙酰乙酸、甘油(P<0.01)、肌酸(P<0.05),除乳酸值仍明显高于运动前外,乙酰乙酸也呈现出明显上升趋势;E1与E4间的差异代谢标志物只有甜菜碱(P<0.05)和甘油(P<0.01);E1与E5间已无差异代谢物。
2.3 受试者运动后不同时刻相比较的血清代谢标志物
由表1可见,运动后即刻与运动后1 h,即E2与E3之间共检测出7种差异代谢物,分别是:谷氨酰胺、甜菜碱、乙酰乙酸、甘油、葡萄糖(P<0.01),谷氨酸、甘氨酸(P<0.05),均明显上升;E3与E4之间共检测出5种差异代谢物,分别是:亮氨酸、异亮氨酸、缬氨酸、乳酸、肌酸(P<0.01),除乳酸明显降低外,均明显上升;E4与E5间已无差异代谢物。
3 讨论
本研究采用核磁共振代谢组学方法研究间歇性大强度运动前后人体血清的代谢组学特征,在运动前与运动后不同时刻共发现13种差异代谢物。通过对这些差异代谢物变化状况的分析可发现间歇性大强度运动的供能特点。
肌酸(Cr)可快速合成ATP,是磷酸原供能系统的主要能源物质。有研究表明补充肌酸可抑制游泳运动员因短时间大强度运动引起的氧化应激标记物的增多,即补充肌酸可起到对抗机体氧化应激的作用[8]。还可以抑制急性大强度运动后炎症标志物血浆肿瘤坏死因子α(TNF-α)和C反应蛋白(CRP)的升高[23]。本研究中,运动后即刻血清中肌酸含量降低(P<0.01),运动后1小时仍较低,至运动后1天才明显高于运动后1小时(P<0.01),并已恢复至运动前水平且略高于运动前,说明本实验大量消耗肌酸供能,体现出大强度运动磷酸原系统供能的特点,但运动后1天可出现超量恢复。
表1 受试者运动前后不同时期血清代谢标志物寻找结果(±s,n=8)
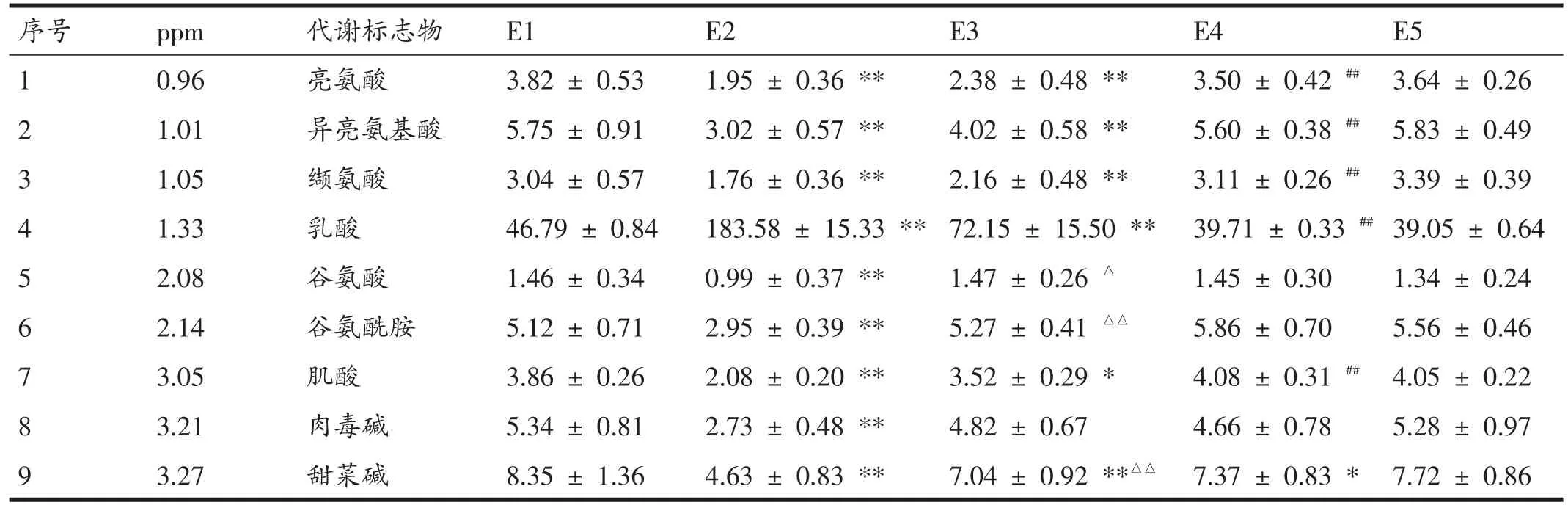
表1 受试者运动前后不同时期血清代谢标志物寻找结果(±s,n=8)
序号1 2 3 4 5 6 7 8 9 ppm 0.96 1.01 1.05 1.33 2.08 2.14 3.05 3.21 3.27代谢标志物亮氨酸异亮氨基酸缬氨酸乳酸谷氨酸谷氨酰胺肌酸肉毒碱甜菜碱E1 3.82±0.53 5.75±0.91 3.04±0.57 46.79±0.84 1.46±0.34 5.12±0.71 3.86±0.26 5.34±0.81 8.35±1.36 E2 1.95±0.36**3.02±0.57**1.76±0.36**183.58±15.33**0.99±0.37**2.95±0.39**2.08±0.20**2.73±0.48**4.63±0.83**E3 2.38±0.48**4.02±0.58**2.16±0.48**72.15±15.50**1.47±0.26△5.27±0.41△△3.52±0.29*4.82±0.67 7.04±0.92**△△E4 3.50±0.42##5.60±0.38##3.11±0.26##39.71±0.33##1.45±0.30 5.86±0.70 4.08±0.31##4.66±0.78 7.37±0.83*E5 3.64±0.26 5.83±0.49 3.39±0.39 39.05±0.64 1.34±0.24 5.56±0.46 4.05±0.22 5.28±0.97 7.72±0.86

(续表1)
葡萄糖(Glu)与乳酸(La)是人体在缺氧状态下糖酵解供能系统主要的能量供应物质和酵解产物。在啮齿动物及人类骨骼肌中,葡萄糖转运蛋白4(GLUT4)对葡萄糖的摄取起主要作用[24]。运动时GLUT4从细胞内部移位到肌膜和横向小管处以增加骨骼肌中葡萄糖的转运[25]。本研究中在运动后即刻血清中葡萄糖急剧下降(P<0.01),说明葡萄糖被快速转变成运动所需的能量,但在运动后1小时即明显恢复(P<0.01)并接近于安静时水平。
乳酸不仅可导致运动疲劳,还可激活缺氧诱导因子-1(HIF-1)以诱导肿瘤细胞侵润[26]。有研究表明,在缺氧状态下进行高强度运动可导致全身乳酸转化率提高18倍[27]。本研究中乳酸含量在运动后即刻显著升高,运动后1小时下降了约二分之一(P<0.01),运动后1天已明显低于运动后1小时(P<0.01),并基本恢复到安静状态。说明糖酵解是本实验的能量来源之一,并且产生的乳酸在运动后1天基本消除。
甘油(TG)是脂肪分解的产物。关于运动时甘油浓度的变化,Ueda等研究表明在中等强度运动过程中血清甘油含量持续增多,运动后即刻达到峰值,运动后30分钟可恢复至安静水平[28]。Ueda等的另一项研究表明递增负荷至最大摄氧量运动后即刻血清甘油含量达到最高值,运动后1小时基本恢复至安静状态[29]。本研究中在运动后即刻甘油大幅度下降(P<0.01),运动后1小时明显升高(P<0.01),但仍低于运动前(P<0.01),运动后1天的含量与运动后1小时基本持平,1周后恢复至运动前水平。说明本实验中脂肪也被氧化为机体供能,尤其在运动间歇时脂肪作为能量消耗,但其恢复速度较慢。
酮体(KB)包括乙酰乙酸、β-羟丁酸和丙酮,在人体内含量极少,约为0.3~5 mg/dL。Laffel研究表明机体在饥饿状态下KB产生增多以替代葡萄糖作为外周组织(大脑,心脏和骨骼肌)的主要能源物质[30]。Mark Evans等发现运动员在长距离自行车运动期间体内酮体被氧化而减少,运动后恢复期又显著增加,且训练有素者骨骼肌利用酮体的能力明显提高[31]。本研究中血清内的乙酰乙酸在运动后即刻急剧下降(P<0.01),运动后1小时明显上升且显著高于运动前和运动后即刻(P<0.01),与Mark的研究结果有相似之处,我们认为是由于受试者是在空腹状态下进行间歇性大强度运动,糖原耗竭、酮体供能加强,运动后又超量恢复所致。
肉毒碱(CAR):左旋肉毒碱是线粒体膜上活化的脂肪酸载体,可携带脂肪酸进入线粒体进行β-氧化[32]。Jamal等研究表明,继发性肉毒碱的缺乏可导致大鼠骨骼肌线粒体功能受损[33]。Wall等研究表明足球运动员口服左旋肉毒碱一个月脂肪供能明显增强[34]。本研究中肉毒碱在运动后即刻急剧下降(P<0.01),运动后1h和1天已基本恢复到运动前水平。再次说明本研究中脂肪也作为能源物质分解供能从而消耗肉毒碱。如果是在非空腹状态下运动,甘油、乙酰乙酸、肉毒碱等物质的代谢状况可能会不同,脂肪代谢的动用程度可能不会如此明显。
谷氨酸(Glu)、谷氨酰胺(Gln)的变化。Smith研究表明,运动员过度训练时,血液中谷氨酰胺浓度明显减少、谷氨酸浓度明显增加,他认为谷氨酰胺持续下降是过度训练的标志之一[35]。在进行高强度力量训练时机体内谷氨酰胺水平可能下降达50%,训练强度越大流失的越快[13]。Koo等的研究表明预先一周每天补充6 g谷氨酰胺,可促进人体急性大强度运动后恢复期血液中氨、磷和肌酸激酶浓度的降低,增加白细胞介素1(IL-1)等细胞因子的产生,进而增强人体免疫功能[7]。本研究运动后即刻,血清中谷氨酸、谷氨酰胺均急剧下降(P<0.01),推测谷氨酸和谷氨酰胺分解供能增加,但运动后1小时即基本恢复至运动前水平,尤其谷氨酰胺在运动后1小时显著高于运动后即刻(P<0.01),表明本实验采用的运动方式未导致机体过度训练。
甘氨酸(Gly)是一种非必需氨基酸,是合成谷胱甘肽、肌酸等的原料,也是一种抑制性神经递质。左群[36]、马海峰[13]等发现大负荷训练后代谢物中甘氨酸含量呈上升趋势,提示甘氨酸浓度上升可能是机体脂肪代谢供能比例上升的标志。但是在短时间、大强度急性运动后血浆甘氨酸含量普遍呈降低或不变的趋势[4,37-39]。本研究血清中的甘氨酸在运动后即刻急剧下降(P<0.01),与Makhro等[38]的研究结果一致,可能是由于甘氨酸用于合成肌酸所致。但运动后1小时甘氨酸浓度即恢复至运动前水平,同时机体甘油和乙酰乙酸含量较高,也证实了甘氨酸浓度上升是机体脂肪代谢比例上升的标志。
支链氨基酸(BCAA)是亮氨酸、异亮氨酸和缬氨酸的统称,属于必需氨基酸。BCAA摄入可降低血液内五羟色胺浓度,以缓解中枢疲劳 ;还可减少肌肉损伤物质肌酸激酶(CK)和乳酸脱氢酶(LDH)的浓度;并使血浆游离脂肪酸增多,加速脂肪供能[40]。有研究提出BCAA在机体无氧供能状态下还可取代葡萄糖供能以减少乳酸产生[41]。本实验中,受试者在运动后即刻血清中三种支链氨基酸均下降(P<0.01),恢复1小时后仍低于运动前水平(P<0.01),推测作为能量进行了消耗,运动1天后基本恢复到运动前水平并高于运动后1小时水平(P<0.01),表明支链氨基酸的消耗可在一天内得到恢复。
甜菜碱(Bet)主要成分是三甲基甘氨酸、强心甙和其他甾类成分,可为合成脂酰胆碱、肉碱等提供甲基。甜菜碱已被证明具有促进脂肪代谢、保护肝脏、抗肿瘤、降低心血管疾病、增强运动能力等功能[42,43]。Cholera等的研究表明运动员补充甜菜碱可防止柠檬酸合成酶热变性、维持渗透压,增强高温环境中高强度运动机体的抗疲劳性[44]。本实验中血清甜菜碱在运动后即刻急剧下降(P<0.01),运动后1小时大幅上升(P<0.01),至运动后1天仍呈上升趋势(P<0.05),运动后一周恢复至安静水平。可能是由于运动期间受试者肌酸、肉碱等大量消耗并合成,甜菜碱作为甲基供体被大量消耗,运动后随着代谢水平的下降逐渐恢复,这种变化状态进一步体现了机体磷酸原系统和脂肪供能的特征,它可能是脂肪供能的潜在代谢标志物。
本研究分析的代谢物也可以作为底物对信号通路产生影响,下一步的研究可通过探讨相关信号转导通路中酶活性、蛋白或基因表达等变化,从分子和代谢两个水平深入探究运动对特征代谢物和信号转导通路的整体调节作用。
4 结论
磷酸原系统、糖酵解系统及有氧氧化系统均参与间歇性大强度运动供能,糖、脂肪、蛋白代谢均发生变化,差异代谢物在运动后即刻与运动前差异最显著,但运动后1小时至1天恢复明显,1周内基本恢复。
猜你喜欢
养猪(2022年1期)2022-11-21
中国畜禽种业(2022年8期)2022-09-17
中国畜牧业(2022年4期)2022-04-02
中国现代中药(2022年1期)2022-03-03
中国学校体育(2018年9期)2018-05-14
国外畜牧学·猪与禽(2018年10期)2018-05-14
国外畜牧学·猪与禽(2018年8期)2018-05-14
国外畜牧学·猪与禽(2018年7期)2018-05-14
运动(2016年17期)2017-01-03
中国体育教练员(2016年4期)2016-11-29
