
濒危半红树植物玉蕊对盐胁迫的生理响应及其相关性分析
2019-10-10 06:07黄秋伟於艳萍黄秋艳刘细妹陈秋佑檀小辉
中南林业科技大学学报 2019年10期
梁 芳,黄秋伟,於艳萍,梁 惠,黄秋艳,刘细妹,陈秋佑,檀小辉
(1.玉林师范学院,广西 玉林 530700;2.广西壮族自治区亚热带作物研究所,广西 南宁 530001)
玉蕊Barringtonia racemosa为玉蕊科玉蕊属植物,俗名水茄冬,棋盘脚,常绿乔木,原产地高可达15m,是我国稀有植物资源,分布区狭窄,生长于非洲、亚洲和大洋洲的热带、亚热带地区,在我国的台湾省及海南岛都有自然分布,但数量很少,且散生居多[1-3]。玉蕊在海南常生长在海滨沿岸,有时和一些高潮位的红树植物混生在一起,是典型的半红树植物,广东、广西和福建沿海有少量引种。2012年廖宝文[4]在广东雷州半岛发现一块有1 000 多株、集中连片的玉蕊天然次生纯林。但钟军弟等[5]的研究发现雷州半岛的玉蕊次生纯林中低龄级和老龄级玉蕊个体数量较少,中龄级个体数量较多,玉蕊种群呈现衰退趋势,且遭受潜叶蛾等虫害,藤本植物攀援于玉蕊冠层,玉蕊生长受到威胁,以及潮水冲刷过急等诸多原因导致玉蕊幼苗向幼树过渡困难,该玉蕊林成龄种群有灭绝的可能。2017年覃海宁等[6]已把玉蕊列为濒危物种。
红树林生境具有高盐、水淹低氧等特征[7],在洪涝多发地区(如沿江、沿海等)培育耐受洪涝灾害的观赏树种,对其景观的营造和恢复具有重要意义[8]。目前,盐度也是红树植物的一个主要环境因素,它已成为红树林造林成活率高低的关键因子。由于长期适应潮间带海水环境,红树植物形成了一套适应于一定盐度的耐盐机制[9]。高盐度会明显阻碍植物的发育,但对红树植物而言,适当的盐度范围可促进其生长发育。林鹏等[10]研究表明,红树植物秋茄Kenaelia candel在海水盐度为7.5‰~21.2‰时生长旺盛。半红树植物与红树植物具有相似的生态功能,可以防风固岸、调节区域气候、保护生物多样性和对各种污染物进行净化处理等[5]。部分半红树植物还具特殊的药用价值和经济价值[11],但目前由于海湾城市构建等各方面的原因,现存半红树资源越来越少,半红植物资源保护日趋紧迫,然而目前国内外对半红树植物的研究还很不完善,在造林经营上的研究尤为缺乏[12]。国内外学者对真红树植物的研究较多,但对我国仅有的12 种半红树植物种适生盐度研究较少,其中杨叶肖槿Thespesia populnea和海芒果Cerbera manghas、银叶树Heritiera littoralis、水黄皮Pongamia pinnata等半红树植物在抗逆性上少量研究[13-15],另外海滨猫尾木Dolichandrone spathacea、玉蕊、莲叶桐Hernandia sonora、苦郎树Clerodendrum inerme4 种只做过体内盐分分布的比较研究[16],而玉蕊盐胁迫下的生理响应以及适生盐度研究尚未见报道。
玉蕊作为半红树植物之一,具有较高的观赏价值、生态保护价值和经济价值[17],在科学研究和环境保护上均具有重大意义,但由于种种原因已被列入濒危物种,可见对其开展相关的生境保护研究、抗水淹、抗盐育种及栽培等相关研究迫在眉睫。因此,本试验探究潮汐系统下半红树植物玉蕊对盐胁迫的生理响应及其各生理指标之间的相关性,以获得玉蕊适生的盐度范围,为玉蕊抗盐育种、栽植区域及人工造林提供理论依据。
1 材料与方法
1.1 试验材料
玉蕊种子采自海南儋州市天然玉蕊林,于2016年10月在广西壮族自治区亚热带作物研究所育苗棚育种,出苗后起苗装入营养袋,所用土壤为果园土∶椰糠=1∶1,培养成苗,待苗高至30±5 cm 时为试验用苗。
1.2 试验方法
试验由储水槽、植物培养槽,定时器、水泵、过滤系统、补光系统及遮阴系统等组成模拟潮汐自动控制装置,即用定时器来自动控制各处理槽里水泵的抽水时间,以达到所设置的试验时间梯度。试验于2018年7月29日—8月19日在广西壮族自治区亚热带作物研究所温室大棚内进行。用自来水添加海盐来配制不同盐度梯度的溶液,设水体盐度为0‰(对照,CK)、2‰、4‰、6‰、8‰、10‰、12‰,14‰共8 个处理,每处理3 个重复,每个重复6 株玉蕊幼苗。模拟半日潮,每12 h 为一个淹水周期,每个半日潮幼苗被淹没时间为3 h,淹浸水位至枝下高。每3 d 测一次生长指标,处理21 d 后采样,取玉蕊幼苗中上部成熟叶进行各项生理指标测定。
1.3 指标的测定
1.3.1 形态指标调查
形态指标以处理前后株高、叶片数、地径变化情况表示,每3 d 测一次。
株高:在淹水处理前后,用卷尺测定植物株高(精确到0.1 cm),每株测定3 次,取平均值。计算株高增长量H1=淹水结束时株高-淹水开始时株高。
地径:每株用游标卡尺东西南北方向各测定两次(精确到0.02 mm),取平均值,测定的植株选择同上。地径增长量D1=淹水结束时地径-淹水开始时地径。
叶片数的测定:植株叶片数直观且容易测定,直接读取,记录数据。叶片变化量L1=淹水结束时叶片数-淹水开始时叶片数。
1.3.2 生理指标测定
超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)法;过氧化物酶(POD)活性测定采用愈创木酚法;丙二醛(MDA)含量的测定采用硫代巴比妥酸(TBA)氧化法;游离脯氨酸含量测定采用茚三酮比色法测定其含量。吸光值测定均采用紫外分光光度计,各指标测定方法参照《植物生理学模块实验指导》中方法并加以调整[18]。
1.3.3 数据分析
所有试验数据采用Excel 2007 整理后再用GraPhPad Prism7.0 软件制图,数据统计分析使用SPSS 19.0 软件,平均值间的比较采用单因素方差分析(One-way ANOVA),多重比较采用邓肯多重范围检验(Duncan's multiple range test);显著性检验水平均设置为0.05。
2 结果与分析
2.1 株高变化
由图1可以看出,经不同水体盐度处理后,玉蕊株高平均总增量都有所生长,但随着盐度的不断增大,各处理株高总增量呈下降趋势(2‰盐浓度除外),当盐度达到12‰时达到最低值,且盐度在4‰~12‰时株高总增长量较对照(0‰)、2‰相比呈显著差异(P<0.05),可见玉蕊在盐度0‰~2‰株高增长较快,盐度在4‰~12‰时玉蕊株高增长较为缓慢,说明不同水体盐度对株高的生长速率有一定的影响。
2.2 地径变化
由图2可以看出,玉蕊幼苗地径总体均有一定程度的增长,随着盐浓度的增大,玉蕊幼苗地径总增量先增高后下降,整体呈双峰变化,峰值出现在4‰、10‰,分别是对照组的126.01%、104.05%,但各处理浓度较对照组均无显著差异。说明不同程度的盐胁迫对地径的增长影响不大,但地径总增量出现双峰值变化,这有可能是玉蕊通过调节地径的增量,进而调节根冠比来不断地适应不同程度的盐胁迫环境。
2.3 叶片数变化
叶片能反映植物生长和发育情况,且是对外界环境反应较为敏感的同化器官。经实验观察,处理期间玉蕊植株大部分叶片为正常绿色叶,极少部分出现叶尖、叶缘和叶脉变黄。随着处理时间的增长,玉蕊少部分老叶和不成熟叶变黄脱落,成熟叶基本无脱落,生长状态良好,因此实验期间对植株正常光合作用和水分代谢等功能影响不大。由图3可以看出,玉蕊叶片数变化整体呈先增加后下降的趋势,最高值出现在2‰盐度处理上,该处理叶片数变化量为正值,其他浓度处理都为负值,且最低值出现在10‰的盐浓度处理上,较对照比呈显著差异(P<0.05),其他处理浓度2‰~8‰植株叶片数变化量较对照比均无显著差异,说明0‰~8‰盐度处理对玉蕊叶片数影响不大,但较高盐度12‰、14‰时叶片脱落数反而小于10‰,说明10‰的盐度可能是玉蕊叶片抗盐能力的一个临界浓度,即此时玉蕊有可能在开始启动自身调节适应盐胁迫的响应机制。
2.4 SOD 活性变化

图3 不同盐度对玉蕊幼苗叶片数的影响Fig.3 Effects of different salinities on leaf number of Barringtonia racemosa

图4 不同盐度对玉蕊幼苗叶片SOD 活性的影响Fig.4 Effects of different salinities on SOD activity in leaves of Barringtonia racemosa
由图4可以看出,不同水体盐度盐胁迫下,玉蕊幼苗叶片SOD 活性的变化呈先升高后降低再升高的趋势,在盐度达到6‰时SOD 活性最低,10‰时达到最高值,但各处理较对照组均无显著差异,说明随着盐度的增大,玉蕊幼苗叶片SOD活性的响应并不明显。
2.5 POD 活性变化
由图5可以看出,不同水体盐度胁迫对玉蕊幼苗叶片POD 活性影响较大,随着盐度的增大,POD 活性均明显高于对照组,整体呈上升趋势,且出现较均匀三峰变化,峰值分别在2‰、6‰、12‰三个不同盐度上,是对照组的201.83%、214.16%、277.17%。POD 活性在12‰时达到最高值,较对照组呈显著差异(P<0.05)。但盐度在0‰~10‰之间POD 值较对照均无显著差异,说明玉蕊幼苗叶片POD 活性在不同水体盐度胁迫下的响应有一定的差异,其呈峰值变化可能是自身在不断调节适应逆境的一种响应机制,表明玉蕊有一定的耐盐性。
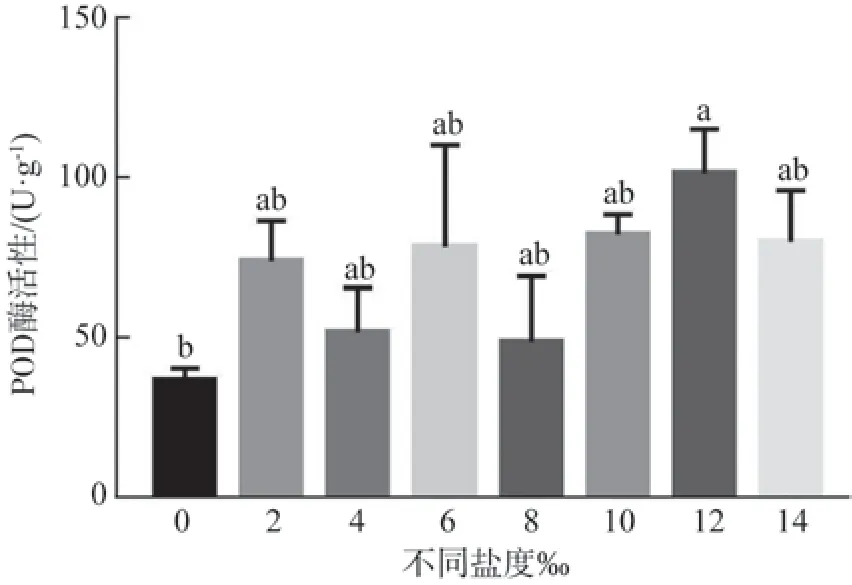
图5 不同盐度对玉蕊幼苗叶片POD 活性的影响Fig.5 Effects of different salinities on POD activity in leaves of Barringtonia racemosa
2.6 丙二醛(MDA)含量的变化
由图6可以看出,随着盐度的增大,玉蕊幼苗叶片的丙二醛(MDA)含量整体呈上升趋势,在盐度12‰时达到最大值,是对照组的136.11%,呈极显著差异(P<0.01)。这与POD 活性变化一致,表明在盐度为12‰时对玉蕊有一定的胁迫,此时植物启动POD 应对逆环境。但在盐度2‰~10‰之间丙二醛(MDA)含量较对照组无显著差异,说明当处理盐度≤10‰时,玉蕊叶片细胞膜受破坏程度不大,盐度为14‰时MDA 含量又恢复到对照水平,玉蕊幼苗表现出一定的抗盐性和适应性。

图6 不同盐度对玉蕊幼苗叶片丙二醛含量的影响Fig.6 Effects of different salinities on malondialdehyde content in leaves of Barringtonia racemosa
2.7 脯氨酸含量的变化
由图7可以看出,随着盐度的增大,玉蕊幼苗叶片的脯氨酸含量整体呈上升趋势,且由处理盐度在0‰~10‰的缓慢升高到12‰~14‰的迅速升高变化。但当盐度在2‰~8‰范围时脯氨酸含量较对照组无显著差异,说明小于8‰的盐度处理对玉蕊的没有造成胁迫。而当盐度在10‰、12‰时脯氨酸含量分别是对照组的233.59%、224.84%,较对照组呈显著差异 (P<0.05),当处理盐度增大到14‰时脯氨酸含量达到最高值,是对照组的681.15%,较对照组呈超显著差异(P<0.001),说明大于8‰的盐度处理对玉蕊可能是一种逆境胁迫,脯氨酸含量急剧上升,表明玉蕊有较好的抗盐性。

图7 不同盐度对玉蕊幼苗叶片脯氨酸含量的影响Fig.7 Effects of different salinities on proline content in leaves of Barringtonia racemosa
2.8 盐胁迫下玉蕊生长指标的相关性分析
由表1可见,不同水体盐度与玉蕊株高的增长呈极显著负相关(P<0.01),与叶片数和地径的变化也呈负相关,但不显著。另外,在控制盐浓度变量的偏相关分析发现,盐胁迫下玉蕊各生长指标之间呈负偏相关,但无显著性。表明随着盐浓度的不断升高,玉蕊的形态指标均受到不同程度的抑制,其中株高的增长受到的影响最为显著,地径和叶片受到的影响不大。

表1 盐胁迫下玉蕊生长指标的相关性†Table1 The correlation of growth indicators of Barringtonia racemosa under salt stress
2.9 盐胁迫下玉蕊生理指标的相关性分析
由表2可知,不同水体盐度与脯氨酸含量呈极显著正相关(P<0.01),与POD 活性呈显著正相关(P<0.05),与其他生理指标相关不显著。另外,在控制盐浓度变量的偏相关分析中发现,盐胁迫下脯氨酸含量与丙二醛含量呈显著负相关(P<0.05),其他指标之间相关性不显著。试验结果表明随着盐胁迫的不断增强,丙二醛随之增大,细胞膜受到破坏,玉蕊通过快速的增加脯氨酸含量来维持细胞正常渗透压,同时通过增强POD 来清除活性氧等有害物质,可见脯氨酸和POD 是玉蕊抵抗盐胁迫逆境的重要响应指标。
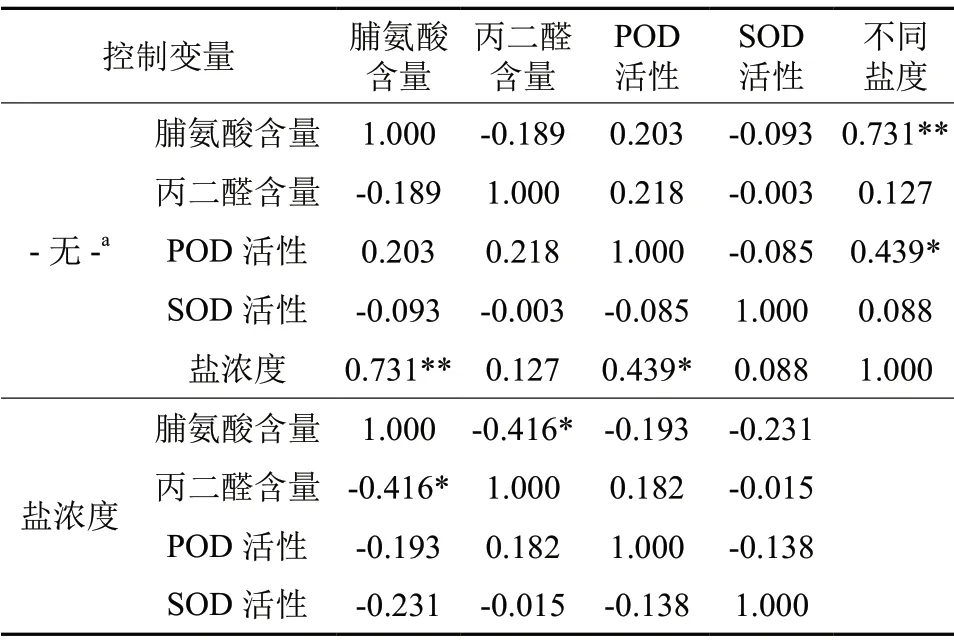
表2 盐胁迫下玉蕊生理指标的相关性†Table2 The correlation of physiological indicators of Barringtonia racemosa under salt stress
3 结论与讨论
3.1 结 论
综上试验结果得知,脯氨酸、POD 是玉蕊响应盐胁迫的重要指标;玉蕊的适生盐度范围是0‰~8‰,适生的临界盐浓度是10‰,盐度10‰~14‰范围表现出较好抗盐性。表明玉蕊对低浓度盐胁迫具有一定的耐受性和适应性。因此,可以考虑在广西、广东、海南、福建等部分红树林域,泥滩、沙滩、和滩涂地区等造林中选用玉蕊防风固沙消浪,固岸护堤、美化绿化,构建我国海岸、河岸等绿色保护屏障。
3.2 讨 论
不同程度的盐胁迫对半红树植物的生长发育有显著影响,生长量是反应植物盐胁迫耐受性的最直接的指标。在本研究中,所有处理的玉蕊幼苗株高、地径均有所增长,但随着盐度的不断升高,株高的增长量受到不同程度的抑制(2‰处理的植株除外),当水体盐度为4‰~12‰时,玉蕊株高生长速率受到显著影响,这与NaCl 胁迫后枸杞Lycium chinense株高变化相似[19]。地径平均增量呈双峰值变化,叶片脱落量先增大后减少(2‰处理的植株除外),这与红花玉蕊Barringtonia acutangula在低盐(0‰~5‰)[20]和秋茄在盐度(0%~2%)[21]下叶片数目增加的研究结果一致。各处理的地径和叶片数较对照组均无显著差异,这表明水体盐度在0‰~14‰范围内对玉蕊幼苗总生长量影响不大,且存在低盐促进高盐抑制的现象,这与王文卿等[22]和陈长平等[23]在研究红树植物木榄Bruguiera gymnorrhiza、无瓣海桑Sonneratia apetala幼苗的生长情况研究结果不同,此现象可能是因树而异因地而异。可见玉蕊具有一定的耐盐性,在构建红树林湿地景观上具有良好的应用前景。
超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)作为植物内源的活性氧清除剂,属于保护酶系统。盐胁迫下,欧李Cerasus humilis植株通过增强保护酶的活性,提高总呼吸及交替呼吸速率,改变离子含量等方式提高植株的抗逆性[24]。秋茄在高盐下启动抗氧化防御系统,通过提高SOD 和POD 活性来抵御高盐引起的氧化应激[25]。本试验结果中,随着水体盐度的不断升高,玉蕊幼苗各处理的叶片SOD 活性一直保持在稍高于对照组水平,较对照无显著差异。而POD 则非常活跃,呈现三峰变化,最大的峰值出现在盐度12‰上,这与叶片MDA 含量、脯氨酸含量变化一致,即这三个指标都在水体盐度为12‰时显著升高,较对照组均呈显著差异(P<0.05),这是玉蕊幼苗对盐胁迫的适应性反应。玉蕊幼苗叶片受高盐(12‰)胁迫后SOD 能与POD 协同作用,并主要启动POD清除盐胁迫植株体内增多的过氧化物,这与前人研究结果相符[26,27],可见POD 是玉蕊响应盐胁迫的重要指标之一,这与其同科同属的红花玉蕊[17]盐胁迫的响应指标一致。
丙二醛(MDA)是膜脂过氧化的终产物,是用来衡量膜质过氧化程度的指标之一。大量研究结果显示,盐胁迫下,植物叶片组织中丙二醛含量会随盐浓度的升高而升高[28-31],但也有研究者发现,紫叶矮樱Prunus×cistenaN.E.Hansen ex Koehne和美国红栌Cotinus coggyria在持续低温胁迫下丙二醛含量呈下降趋势[32]。本研究结果中,玉蕊受到0‰~10‰的水体盐度胁迫后,叶片膜系统受损不严重,但当盐度升高至12‰时叶片膜系统受到极显著影响,此时POD 活性达到最大值,较对照呈显著差异,脯氨酸也在显著上升,可见玉蕊在应对高盐胁迫时主要启动POD 抵抗和迅速提高脯氨酸含量以调节细胞渗透势,以致在盐度为14‰时MDA 含量又恢复到对照水平,这与彭英等[33]研究北沙参Glehnia littoralis盐胁迫下叶片组织的膜脂过氧化水平变化相似,此时脯氨酸含量达到了最高值,这与吴竹妍等[34]的研究结果一致。表明玉蕊应对高盐胁迫有其内在的响应机制,脯氨酸是玉蕊响应盐胁迫的重要指标之一,这与红花玉蕊盐胁迫下的响应指标一致。
通过相关性分析发现,在生长指标上,同一盐浓度下,株高地径叶片数相互呈负偏相关,但不显著。不同盐浓度下,盐度与株高呈极显著负相关(P<0.01),与地径叶片数负相关,但不显著,表明随着盐度的增大,玉蕊幼苗株高生长速率受到极显著抑制(P<0.01),但盐度对地径叶片数影响不大。在生理指标上,不同盐度与POD 酶活、脯氨酸均呈显著正相关,说明随着盐度的不断增大,玉蕊幼苗叶片的POD 和脯氨酸含量也不断增加,以抵抗逆环境。表明玉蕊具有较好的抗盐性。另外,在控制盐度下,脯氨酸含量和丙二醛含量呈显著负相关,表明玉蕊应对盐胁迫通过迅速提高脯氨酸含量调节细胞渗透势,降低了丙二醛含量,直接保护了细胞膜,这与红树植物小花木榄B.Parviflora[35]和白骨壤Avicennia marina[36]在高盐胁迫下的响应相似。
生长在海岸潮间带的半红树植物和红树植物一样均要面临高盐、水淹缺氧等复杂生境。目前在生理水平上虽有一定关于半红树植物对水淹和高盐胁迫下的光合作用特性、抗氧化酶活性、生物量变化的研究,但半红树植物玉蕊已属濒危植物,且在盐胁迫逆境中生理及分子水平上的研究尚未见报道,因此本试验设计中只借鉴部分半红树植物的研究方法,有一定的局限性,试验结果得出玉蕊在小于14‰的水体盐度有较好的抗性,但对大于14‰的盐度玉蕊的耐受性及适应性如何均未知,因此对高盐度、长时间淹水胁迫后玉蕊植株的生理响应机制及耐盐耐淹机理有待进一步研究,同时基于逆境胁迫后能直接与ROS 反应,也可作为酶的底物出现在ROS 清除机制中的一些次级代谢物质的研究也是非常重要的方向,以及胁迫后植物特异蛋白的合成及表达等分子水平上的相关内容将在以后的研究中做进一步探讨。
猜你喜欢
中老年保健(2022年3期)2022-11-21
小猕猴学习画刊(2022年4期)2022-05-05
思维与智慧·下半月(2022年2期)2022-02-16
老年博览·上半月(2021年10期)2021-11-02
安徽化工(2021年3期)2021-05-29
棉花学报(2020年3期)2020-08-08
河南水产(2020年1期)2020-06-29
水产科学(2020年3期)2020-05-25
科学与财富(2019年15期)2019-10-21
东坡赤壁诗词(2018年3期)2018-07-16
