
降水和氮沉降对内蒙古半干旱草地羊草和大针茅叶片碳交换的影响
2019-09-25 11:03张晓琳翟鹏辉黄建辉
草地学报 2019年4期
张晓琳, 翟鹏辉, 黄建辉
(1. 山西农业大学, 山西 太谷 030801; 2. 中国科学院植物研究所, 北京 100093)
水分是限制植物代谢活动最重要的因素,降水格局变化是全球气候变化研究中的重要内容,有报道指出中国北方地区的年降雨量在2070-2100年间可能会增加30%[1]。降雨量改变将对植物个体生长以及叶片水平碳循环产生很大的影响,尤其是在受水分限制严重的半干旱地区[2-3]。降雨的改变不仅改变物种的物候及寿命[4],而且能够影响植物叶片光合生理特性,大多数研究结果发现降雨增加会促进植物叶片光合速率[5-7]。降雪也是降水的重要形式,虽然中国北方地区的降雪量少,但持续时间可能长达5个月以上。据报道北半球高纬度地区变暖以及海冰融化引起水汽增加,导致我国北方地区降雪量增加[8-9]。近年来有关降雪对生态系统碳循环的影响逐渐引起了国内外学者的重视,但这些研究多数集中在高纬度或者高海拔地区的森林生态系统[10-11],关于温带草地生态系统的研究相对较少,尤其是对植物叶片水平碳交换的影响更是有待于进一步加强。
氮是影响植物生长至关重要的因素,有研究发现在过去的五十年全球氮沉降有明显的增加,而且在未来将持续增加[12]。氮沉降增加将影响生态系统的各个过程,尤其是对叶片水平碳交换过程有至关重要的作用[13]。有关氮沉降的研究逐渐展开,大部分研究发现氮沉降提高叶片水平碳交换,但也有研究发现氮沉降对叶片水平碳交换作用较弱甚至起到抑制作用[14-15]。因此,氮沉降对植被叶片水平碳交换是如何影响的需要进一步研究。
为研究植物叶片碳交换是如何响应全球降水格局变化和氮沉降的增加,本研究利用已经建立的氮、水添加平台,选取内蒙古锡林郭勒典型草原为研究对象,针对该草地群落两个优势物种羊草(Leymuschinensis)和大针茅(Stipagrandis)探讨以下问题:(1)增水的方式不同即春季增雪和夏季增雨,羊草和大针茅的叶片碳交换的响应是否相同?(2)施氮处理下,羊草和大针茅的叶片碳交换的响应机制是什么?本实验通过春季增雪、夏季增雨和施氮处理,研究羊草和大针茅的叶片碳交换的响应机制,对揭示未来全球变化背景下降水与氮沉降增加对草地植被的影响及其响应机制具有重要作用。
1 材料与方法
1.1 实验地概况:
本实验地点位于内蒙古草原锡林河流域,中国科学院内蒙古草原生态系统定位研究站附近(116°40′ E,43°33′ N,海拔1 251 m)。该地区年均温约0.4℃,年均降水量334.7 mm,其中降雨占91.9%,降雪占8.1%;2010年11月到2011年10月总降水量231.7 mm,其中降雨占86.2%,降雪占13.8%[16]。土壤类型属于栗钙土,pH值在6.6~8.0之间[17]。植被类型属于温带典型草原,主要植物有羊草(LeymuschinensisTzvel.),大针茅(StipagrandisSmirn.),西伯利亚羽茅(AchnatherumsibiricumKeng),冰草(AgropyroncristatumGaertn.)和糙隐子草(CleistogenessquarrosaKeng)等。
1.2 实验设计
本研究采用完全随机区组设计,包括模拟降水和氮沉降两种因素,其中降水处理包括增加降雪和降雨。本实验共设6个处理:对照(无增水无施氮,N0W0),春季增雪(N0W1),夏季增雨(N0W2),施氮(N1W0),施氮并春季增雪(N1W1),施氮并夏季增雨(N1W2)。各处理设3个重复,共计18个样方,样方面积大小为25 m2(5 m × 5 m),样方间过道大于1 m。
春季增雪始于2010年,每年3月初将雪加入到增雪样方中,增雪量为25 mm雪水当量;同年进行夏季增雨,从6月15日到8月中旬,每周一次,每次10 mm,共10次,增雨总量为100 mm。施氮始于2009年,每年7月初将尿素施入施氮样方中,施氮量为10 g·m-2。
1.3 取样与测定
1.3.1光响应曲线的测定 在实验样地中设置固定样方(1 m×1 m),各样方分别选择具有代表性的5株羊草和5丛大针茅植株并进行标记,每次选择1株或丛完整的叶片用于光响应曲线的测定。在自然状态下,利用便携式光合作用测量系统LI-6400(Li-Cor Inc.,Lincoln,NE,USA),连接红蓝光源探头,设定光照强度由高到低分别为2000,1500,1200,1000,800,500,300,200,150,100,50,20,0 μmol · m-2· s-1,测定叶片光合速率。于2011年7月中旬到9月底测定羊草和大针茅光响应曲线,测定时间为增雨处理后的第5-6 d,半月1次,共计5次。
为研究增水和施氮对羊草和大针茅的影响,分别测定其光响应曲线,根据公式计算得到最大光合速率(Amax)、光饱和点(LSP)、光补偿点(LCP)、最大表观量子效率(AQE) 和呼吸速率(Rd)[18-19]:

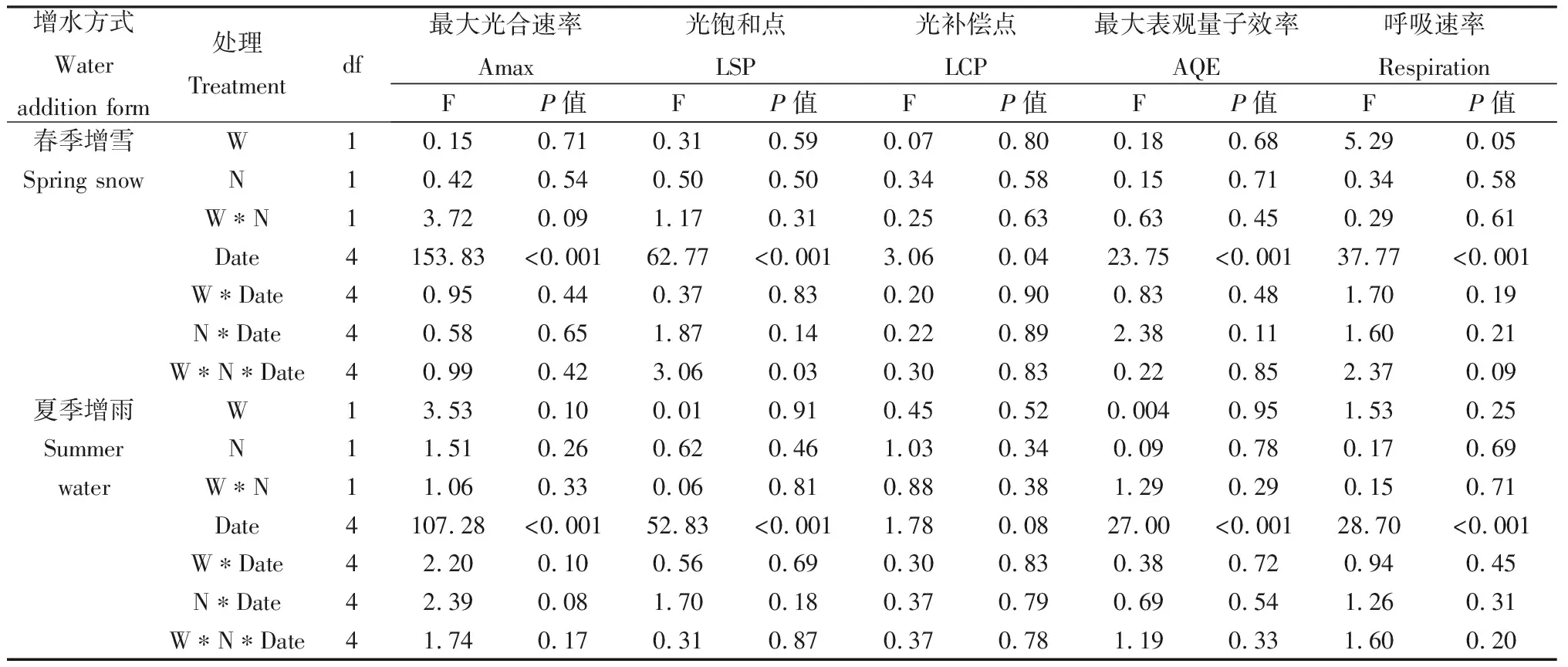
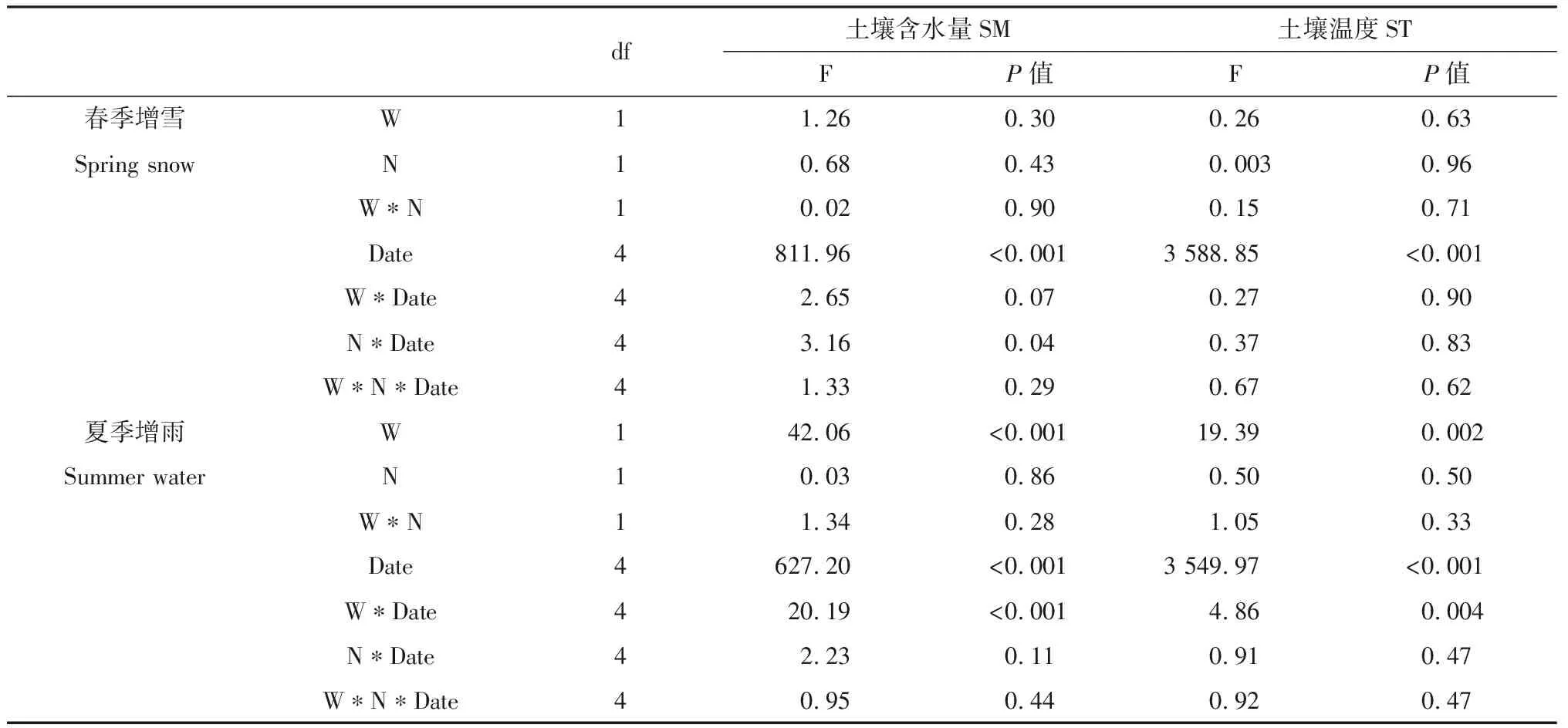
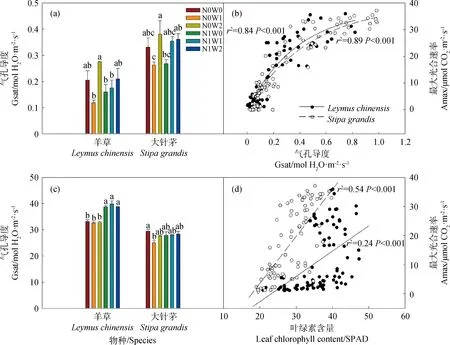
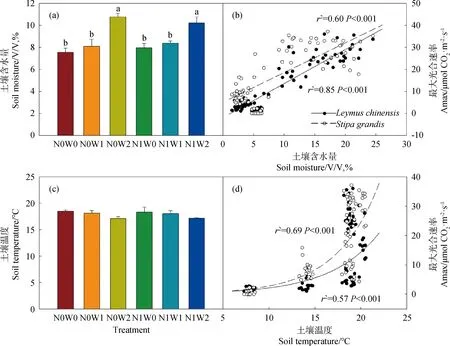
Pn代表净光合速率(μmol CO2· m-2· s-1),AQE代表最大表观量子效率(mol CO2· mol-1photons),PAR代表光合有效辐射(μmol · m-2· s-1),Amax代表最大光合速率(μmol CO2· m-2· s-1),k代表凸性系数(0 1.3.2叶绿素含量的测定 利用便携式叶绿素仪SPAD-502Plus(Konica Minolta Sensing,INC.)测定羊草和大针茅叶绿素含量。在测定光合速率的同时监测其叶绿素含量,重复5次取其平均值,计为该叶片的叶绿素含量。 1.3.3土壤含水量和土壤温度的测定 每次测定羊草和大针茅光合速率时,同时监测土壤含水量和土壤温度。利用土壤水分速测仪TDR-200 (Spectrum Technologies Inc.,Plainfield,IL,USA) 监测土壤含水量,连接10 cm探针,测量固定样方中部土壤体积含水量(V/V,%)。利用智能数字测温仪TH-212(北京海创科技有限公司,北京,中国)监测土壤温度,测量固定样方15 cm土壤温度(oC)。 用三因素重复测定方差分析法(Three-way repeated ANOVA)分析取样时间、增水(春季增雪或夏季增雨)和施氮及其交互作用对羊草和大针茅最大光合速率、光饱和点、光补偿点、最大表观量子效率、呼吸速率、气孔导度、叶绿素含量以及土壤含水量和土壤温度的影响。利用线性或非线性回归法分析羊草和大针茅的最大光合速率与光合有效辐射为2000 μmol · m-2· s-1下的气孔导度、叶绿素含量、土壤含水量和土壤温度之间的相关性。 羊草和大针茅光响应曲线的结果显示,随着光合有效辐射增加,两种植物净光合速率呈现饱和性响应特征,先快速增加,然后增长减缓,最终趋于逐渐饱和(图1a和c)。 图1 羊草(a和b)和大针茅(c和d)的光响应曲线和最大光合速率的均值图Fig.1 Mean light response curves and the maximum photosynthesis of L. chinensis (a and b) and S. grandis (c and d)注:不同小写字母表示处理间差异显著(P < 0.05),下同Note:Different lowercase letters indicate significant differences among treatments at the 0.05 level,the same as below 通过光响应曲线,我们进一步计算了羊草和大针茅最大光合速率、光饱和点、光补偿点、最大表观量子效率和呼吸速率。结果发现夏季增雨显著提高了羊草最大光合速率22.8%,一定程度上提高大针茅最大光合速率8.3%,但对以上其它光合作用特性参数影响不显著,而春季增雪仅显著降低大针茅呼吸速率23.4%,对以上各参数的作用均不显著(图1b和d,图2,表1和2)。 表1 三因素重复测定方差分析取样时间、增水(春季增雪或夏季增雨)和施氮及其交互作用对羊草光合作用生理指标的检验结果(F和P值)Table 1 Results (F and P-values) of three-way repeated ANOVA analysis on effects of sampling date, water addition including spring snow or summer water addition,N addition (N),and their interactions on the photosynthetic physiological characteristics of L. chinensis 表2 三因素重复测定方差分析取样时间、增水(春季增雪或夏季增雨)和施氮及其交互作用对大针茅光合作用生理指标的检验结果(F和P值)Table 2 Results (F and P-values) of three-way repeated ANOVA analysis on effects of sampling date, water addition including spring snow or summer water addition,N addition (N),and their interactions on the photosynthetic physiological characteristics of S. grandis 另外,在增雪处理下,施氮对羊草和大针茅所有光合作用参数均没有显著影响;在增雨处理下,施氮显著提高羊草光补偿点99.6%和呼吸速率37.3%,对羊草和大针茅其它光合作用指标影响均不显著。夏季增雨与施氮的交互作用仅显著提高了羊草的光补偿点,但春季增雪或夏季增雨和施氮对羊草和大针茅各参数指标的交互作用均不显著。随着时间变化,以上各指标变化显著,仅春季增雪处理下施氮对羊草最大表观量子效率和夏季增雨对羊草最大光合速率的作用随时间变化显著。 图2 羊草和大针茅光饱和点(a)、光补偿点(b)、最大表观量子效率(c)、呼吸速率(d)均值Fig.2 Mean values of light saturation points (LSP,a),light compensation points (LCP,b),the maximum appearent quatum efficiency (AQE,c) and respiration rate (d) of L. chinensis and S. grandis 气孔导度和叶绿素含量是决定叶片光合作用变化的重要因素。通过对监测到的气孔导度和叶绿素含量进行分析,发现春季增雪一定程度上降低大针茅叶绿素含量7.2%,但对羊草叶绿素含量、羊草和大针茅气孔导度的影响均不显著;夏季增雨一定程度上提高了羊草气孔导度32.8%,提高大针茅气孔导度24.9%,但对叶绿素含量的作用均不显著(图3a和c,表3)。 在增雪处理下,施氮显著提高羊草叶绿素含量19.7%,但对大针茅叶绿素含量、羊草和大针茅气孔导度的影响均不显著;在增雨处理下,施氮显著提高羊草叶绿素含量17.2%,对大针茅叶绿素含量、羊草和大针茅气孔导度作用均不显著;春季增雪与施氮对大针茅的气孔导度的交互作用显著,夏季增雨与施氮对羊草和大针茅气孔导度和叶绿素含量不存在显著交互作用。随着时间变化,以上各指标变化显著,仅春季增雪并施氮处理对大针茅叶绿素含量的作用随时间变化显著。 土壤含水量和土壤温度是影响植物生长最重要的非生物因子。本研究结果显示,夏季增雨显著提高土壤含水量35.3%,降低了土壤温度3.5%,但春季增雪和施氮对土壤含水量和土壤温度的影响均不显著,春季增雪或夏季增雨与施氮对土壤含水量和土壤温度也不存在显著的交互作用(图4a和c,表4)。随着时间变化,土壤含水量和土壤温度变化显著。夏季增雨处理对土壤含水量和土壤温度的作用,春季增雪处理下施氮对土壤含水量的作用,均随时间变化显著(表4)。 表3 三因素重复测定方差分析取样时间、增水(春季增雪或夏季增雨)和施氮及其交互作用对羊草和大针茅气孔导度和叶绿素含量的检验结果(F和P值)Table 3 Results (F and P-values) of three-way repeated ANOVA analysis on effects of sampling date, water addition including spring snow or summer water addition,N addition (N),and their interactions on the stomatal conductance and leaf Chlorophyll content of L. chinensis and S. grandis 表4 三因素重复测定方差分析取样时间、增水包括春季增雪或夏季增雨和施氮及其交互作用对土壤含水量和土壤温度的检验结果(F和P值)Table 4 Results (F and P-values) of three-way repeated ANOVA analysis on effects of sampling date,water addition including spring snow addition and summer water addition,N addition (N),and their interactions on soil moisture (SM,V/V %) and soil temperature (ST,℃) 气孔导度较低时,羊草和大针茅最大光合速率随叶片气孔导度增加快速增加;当气孔导度增加到一定程度时,羊草和大针茅最大光合速率增加速率逐渐下降,最终呈现一种相对稳定的状态;气孔导度对羊草和大针茅最大光合速率变化的解释度分别为84%和89%(图3b)。随着叶绿素含量增加,羊草和大针茅最大光合速率呈线性增加,其中对羊草和大针茅最大光合速率变化的解释度分别为24%和54%(图3d)。 图3 羊草和大针茅气孔导度和叶绿素含量均值与其最大光合速率的关系图Fig.3 Mean values of the stomatal conductance (Gsat) and leaf Chlorophyll content and their relationships with the maximum photosynthesis (Amax) of L. chinensis and S. grandis 羊草和大针茅最大光合速率与土壤含水量之间的关系显示,随着土壤含水量增加,羊草和大针茅最大光合速率均呈线性增加(图4b),水分对羊草和大针茅最大光合速率变化的解释度分别为85%和60%。羊草和大针茅最大光合速率与土壤温度之间的关系显示,随着土壤温度增加,羊草和大针茅最大光合速率指数增加(图4d),温度对羊草最大光合速率的解释度为57%,对大针茅的解释度为69%,表明内蒙古草原地区对植物光合作用特性影响的同时受水分和温度的影响。 水分是限制植物生长和养分循环过程的重要因素,尤其是干旱、半干旱草地生态系统。关于水分是如何影响植物叶片光合生理特性的研究,大部分结果表明增水促进光合能力,主要归因于其直接促进植物的生理活动或者通过提高微生物活性使得土壤中营养元素更易被植物吸收[20]。 首先,水分供给变化将改变植物叶片气孔导度和叶绿素含量[21-22],从而影响植物光合生理特性。气孔被认为是植物与外界环境进行CO2及其他气体交换的主要通道,外界环境对碳交换过程的影响主要通过调节气孔导度大小影响CO2吸收和水分耗散间的权衡达成的[23]。增水,尤其是夏季增雨,通过提高羊草和大针茅气孔导度,增加CO2吸收,促进胞间CO2浓度增加,促进其最大光合速率。由于水分是电子传递过程中不可或缺的供体,增水提高了羊草和大针茅的电子传递速率[24],从而增加了植物叶片光合作用能力。本研究结果也显示,春季增雪降低了大针茅叶绿素含量使得其光合能力降低,但同时降低大针茅的呼吸作用,导致大针茅最大光合速率的响应不显著。春季增雪对羊草光合生理特性的响应不显著,主要原因可能是春季增雪后经过较长时间的干旱状态,使得增雪引起的短期土壤增加水分很快由于强烈的蒸发而消失,对植物生长的影响很小,表现在对植物气孔导度等植物生理特性没有显著的影响。有研究发现不同降雪量大小对植物生长有至关重要的作用[16,25],但是由于本实验中年降雪量小,可能导致降雪量变化对叶片水平碳循环效果不显著。本研究表明增水对植物叶片碳交换的影响,主要通过改变气孔导度影响植物自身生理特性和光合能力,与叶绿素含量的变化关系较弱,从一定程度上表明水分对该地区草地植物的调控主要通过改变植物的气孔导度实现。 图4 土壤含水量和土壤温度均值及其与羊草和大针茅最大光合速率的关系图Fig.4 Mean values of soil moisture (V/V,%) and soil temperature and their relationships of the maximum photosynthesis (Amax) on L. chinensis and S. grandis 其次,增水提高土壤含水量和降低土壤温度,通过改善土壤微环境,提高微生物的活性,促进物质循环过程,增强植物对营养物质的吸收,促进植物生长,增加羊草和大针茅光合生理特性。另外,有研究表明羊草适宜水肥条件好的环境,而大针茅在干旱和养分胁迫的条件下更具竞争力[18],因此由于2011年降水量过低,导致即使增加100 mm降水,夏季增水样方的年降水量刚刚达到多年降水均值水平,所以对大针茅来讲只是一定程度增加叶片碳交换,而羊草对水分响应更敏感,所以增水作用更显著。 氮素是形成叶绿素的重要营养元素,直接或间接影响着植物的光合生理特性,进而影响植物的生长。普遍认为施氮通过影响土壤微生物群落结构和土壤理化性质[26],改变叶绿素含量合成,进而影响植物对大气CO2的羧化能力[22],从而改变植物对光的竞争能力,最终影响植物的生长发育[27-28]。但本研究发现在增雪或者增雨处理下,施氮对羊草和大针茅最大光合速率的影响均不显著,主要可能有以下原因:(1)施氮仅在增雨处理下提高了羊草的呼吸速率和光饱和点,但对羊草和大针茅光响应曲线其它参数指标影响不显著;(2)施氮对羊草和大针茅气孔导度的影响不显著,意味着进入叶片的CO2浓度和水分流失的平衡不变;(3)施氮提高羊草叶绿素含量,但对大针茅叶绿素含量的影响不显著,说明羊草的大气CO2羧化能力提高而针茅的大气CO2羧化能力不变,导致羊草对光的竞争能力提高而大针茅对光的竞争能力不变;(4)施氮对土壤含水量和土壤温度影响不显著,对植物的生长以及羊草和大针茅光合生理特性的影响较弱。说明氮素对植物的最大光合速率的影响共同决定于羊草和大针茅自身光合生理特性的响应情况和外界环境的变化,从一定程度上表明氮素对该地区草地植物的影响主要通过改变叶片叶绿素含量和植物呼吸速率实现。 大量研究发现,氮素对碳循环的影响依赖于外界降水量的大小[16,29],该年度降水量相对较低(较多年降水量均值低103 mm),导致氮素对叶片水平碳循环的作用较弱。另外,羊草和大针茅对气孔导度、叶绿素含量、土壤含水量和土壤温度的相关解释度说明,在增水和施氮处理下,羊草和大针茅叶片碳交换的响应同时受自身光合生理特性和环境因素的影响。 在该半干旱草原,水分和氮素对于植物叶片碳交换的影响主要通过改变羊草和大针茅自身的光合生理特性,进而改变其光合能力、影响其碳交换来实现。增水主要能调节气孔导度、改变叶片胞间CO2浓度和内部水分,进而影响植物的最大光合速率。同时,增水通过改变土壤含水量和土壤温度,间接影响植物光合速率,改变叶片碳交换。施氮对羊草和大针茅最大光合速率的影响均不显著,主要归因于施氮增加叶片叶绿素含量提高光合能力的同时,提高了植物呼吸速率,最终导致叶水平碳交换对氮素响应不显著。本研究说明降水和氮沉降改变对植物叶片碳交换的影响同时受植物本身光合生理特性和土壤微环境的影响。1.4 统计分析
2 结果与分析
2.1 增水和施氮对羊草和大针茅光响应的影响
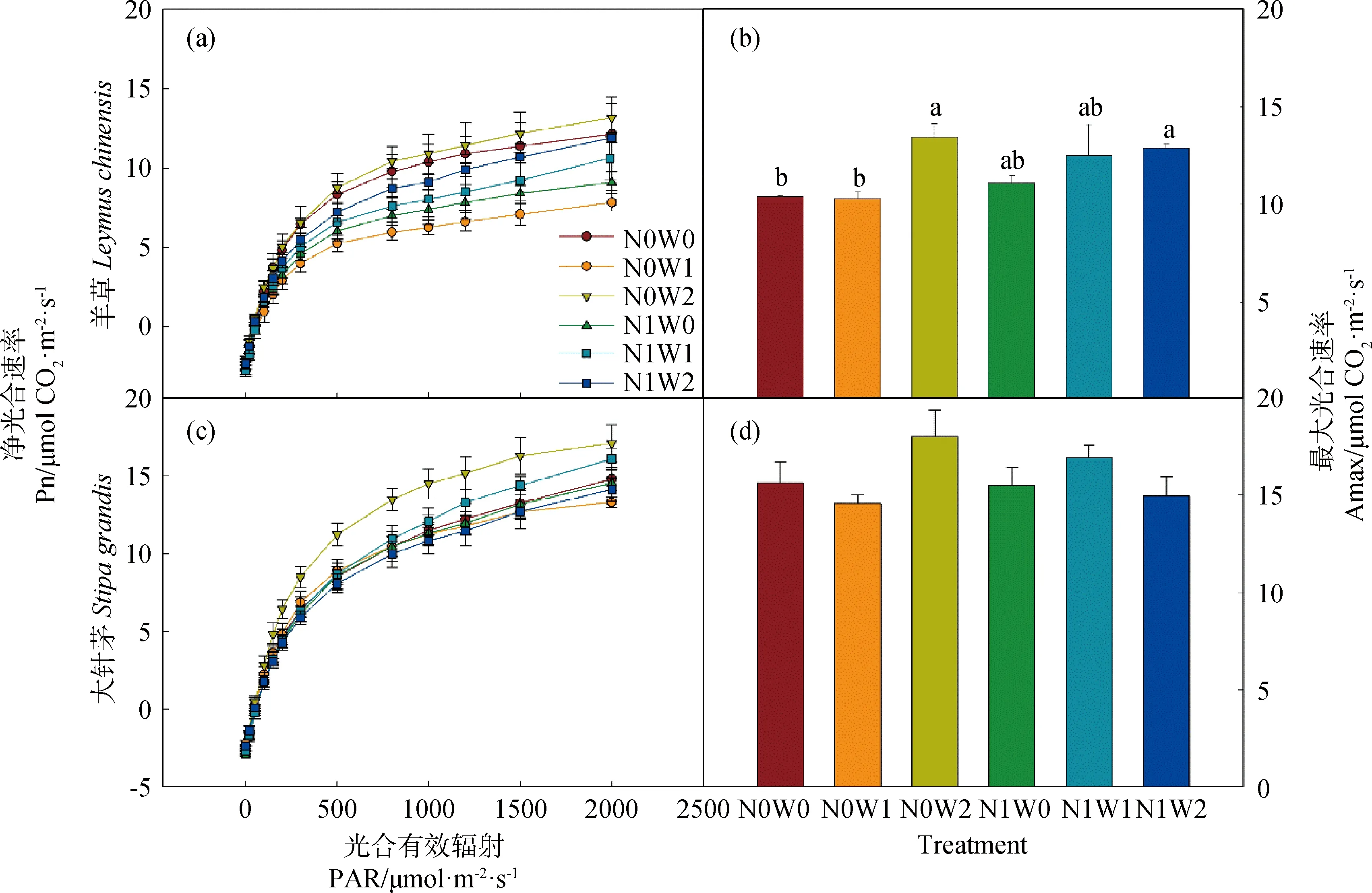


2.2 增水和施氮对羊草和大针茅气孔导度和叶绿素含量的影响
2.3 增水和施氮对土壤含水量和土壤温度的影响

2.4 羊草和大针茅最大光合速率与气孔导度、叶绿素含量和非生物因子的关系
3 讨论
3.1 增水对羊草和大针茅光合生理特征的影响
3.2 施氮对羊草和大针茅光合生理特性的影响
4 结论
猜你喜欢
中国农业科学(2022年11期)2022-06-27
农业工程学报(2022年3期)2022-04-16
草原与草业(2022年1期)2022-04-07
现代畜牧科技(2021年4期)2021-07-21
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
种子(2020年4期)2020-12-22
节水灌溉(2020年9期)2020-09-16
水土保持研究(2019年6期)2019-10-19
中国水土保持科学(2017年3期)2017-07-05
