
金川矿区本土植物光合生理生态日变化特征
2019-09-21 02:34王引弟巨天珍周玉霞万子栋方向文高天鹏
生态科学 2019年4期
王引弟, 巨天珍, 周玉霞, 万子栋, 方向文, 高天鹏
金川矿区本土植物光合生理生态日变化特征
王引弟1, 巨天珍1, 周玉霞1, 万子栋1, 方向文5, 高天鹏2, 3, 4, *
1. 西北师范大学地理与环境科学学院, 兰州 730070 2. 甘肃省矿区污染治理与生态修复工程研究中心, 兰州 730070 3. 祁连山北麓矿区生态系统与环境野外科学观测研究站, 金昌 737100 4. 西安文理学院生物与环境工程学院, 西安 710065 5. 兰州大学生命科学学院, 兰州 730070
采用便携式LI-6400光合测定仪, 选择晴朗天气对金川矿区自然生长的软紫草()、蒺藜()、木本猪毛菜()叶片的气体交换参数及环境因子进行了测定, 通过对比分析三种植物光合特性及其与环境因子间的关系, 以揭示三种植物对矿区环境的适应特征。结果显示: 软紫草和木本猪毛菜的净光合速率日变化有明显的“光合午休”现象, 软紫草产生“午休”的主要原因为非气孔限制因素, 而木本猪毛菜的主导因素为气孔限制因素。其中软紫草和木本猪毛菜的净光合速率日变化最大峰值均出现在11:00和17:00左右。蒺藜“光合午休”现象不明显, 主要原因为气孔限制因素, 最大峰值出现在9:00左右。总之, 三种植物的水分利用效率存在时空差异, 在强光、高温、低湿荒漠环境中都具有一定的生存优势, 对矿区生境的适应能力大小依次为蒺藜、木本猪毛菜、软紫草。
光响应曲线; 日变化; 软紫草; 蒺藜; 木本猪毛菜
0 前言
金川矿区地处金昌市西南部龙首山脉北坡, 由于人类频繁开采矿物资源, 造成植被退化、空气污染等一系列生态环境问题, 绿色植被作为生态系统的支柱和核心, 在保持水土、恢复植被等方面发挥着至关重要的作用。然而, 金川矿区植被种类单一, 群落层次简单, 主要植被优势种有木本猪毛菜()、盐生草()、软紫草()、蒺藜()等。因此, 探讨本土植被对生境的适应性和生理机制, 对于矿区生态系统的恢复具有现实意义。
光合作用是一个非常复杂的生物、化学、物理过程[1], 对绿色植物的生长发育至关重要, 其受到外界环境和内部因素的共同影响, 如大气温度、光合有效辐射、空气相对湿度等环境因子变化都会引起净光合速率, 气孔导度, 胞间CO2浓度等光合参数的变化。测定植物的光合特性可以获得其生理参数, 有助于判定植物光合作用运转状况、光合能力及环境变化对其的影响程度等[2-3]。木本猪毛菜()为小灌木, 叶半圆柱状、肉质, 生于砾质荒漠、干旱山坡、砂丘和山前平原, 具有抗风耐旱、耐瘠薄的特性, 对维护荒漠生态系统的稳定性起重要作用[4]。软紫草()属紫草科软紫草属, 多年生草本, 分布于青海、甘肃、宁夏等地, 生长在海拔2000—4000 m的砾石山坡、草原及草甸等处[5]。蒺藜()生于沙地、荒地、山坡, 全球温带广泛分布[6]。目前, 国内外对这几种植物的研究主要集中在植物滞尘[7]、叶片表型结构[8]、不同生境条件下结实特性的差异[9]以及生理生化[10]等方面, 而对其光合特性研究较少, 尤其是光合日变化的研究尚未见报道, 为此, 本文探讨了荒漠植物软紫草、蒺藜、木本猪毛菜在金川矿区的光合生理生态日变化特征, 为金川矿区本土植物的恢复和保护提供一定的科学依据。
1 材料与方法
1.1 研究区概况与材料
试验在金昌市金川区尾矿坝, 位于101°04'35"—102°43'40"E, 37°47'10"—39°00'30"N, 属大陆性温带干旱气候, 太阳辐射高, 干旱少雨, 昼夜温差大, 蒸发量大, 年均气温4.8 ℃, 年均日照时间2 878 h, 无霜期141 d, 多年平均降水量在110 mm左右[11]。于2017年8月下旬(8月20日—8月31日), 选择晴朗的天气, 在固定样地对自然条件下生长的软紫草、木本猪毛菜、蒺藜叶片的光合日变化及环境因子等指标进行测定。
1.2 指标测定与方法
选择晴朗天气于9:00—12:00, 打开并调试好便携式光合系统分析仪Li-6400XT(安装簇状叶室), 使用开放气路, 设定相关参数, 叶室内CO2浓度设置为400 μmol·mol–1, 叶温为25 ℃, 空气流速为500 μmol·s–1。每个光照强度下适应5 min后测定, 设定光强梯度为2000、1800、1600、1400、1200、1000、800、600、400、300、200、100、50、20、0 μmol·m–2·s–1。测定时每隔3 min 由系统自动记录每个光照强度下软紫草、蒺藜和木本猪毛菜叶片的净光合速率(Pn ), 进行Pn—PAR响应曲线的拟合。用低光强下(200 μmol·m-2·s-1以下)Pn—PAR响应曲线的斜率作为表观量子效率[12]。

注: 研究区边界使用中国行政区划1:400万地图数据, Dem高程数据来源于地理空间数据云。
Figure 1 Topographic map of the study area
选择生长健壮, 长势一致, 同一叶位的成熟叶片, 从当地时间9:00—19:00, 每隔2 h采用便携式Li-6400XT光合作用气体分析系统, 分别测定软紫草, 蒺藜, 木本猪毛菜叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)及主要环境因子如光合有效辐射(PAR)、叶片温度(TL), 大气温度(Ta)、空气相对湿度(RH)及大气CO2浓度(CO2)等生理生态指标, 每种植物测定3株, 在各自光强下重复3次读数取均值。实验采用2 cm×3 cm 标准叶室。为避免相互遮挡, 测定时植株叶片应尽量平铺于整个叶室, 光合测定结束后剪下叶室中的叶片, 经扫描后用CI-400 图像分析软件(CIAS, CID Inc.USA)进行叶面积校正, 根据实际叶面积折算净光合速率、蒸腾速率、气孔导度等气体交换参数。并计算以下指标: 水分利用效率(WUE)(WUE= Pn/Tr)、羧化效率(CE)、气孔限制值(LS)[13,14]。
1.3 数据处理
实验数据采用Excel2010软件进行整理制图, 使用Spss21.0统计软件进行相关性分析。
2 结果分析
2.1 三种植物的光合—光响应
由图2显示: 软紫草、木本猪毛菜、蒺藜的净光合速率随光合有效辐射的增加而升高, 当光合有效辐射达到一定值后, 净光合速率达最大值, 其值分别为8.33、2.37、10.46 μmol·m-2·s-1, 即软紫草、木本猪毛菜、蒺藜达到光饱和点1600 μmol·m-2·s-1、1400 μmol·m-2·s-1和1600 μmol·m-2·s-1。当光合有效辐射继续增加时, 净光合速率略有所下降。采用二次曲线方程拟合, 拟合效果较好(R2>0.984), 得响应曲线方程分别为:

由图3显示, 在弱光下(200 μmol·m-2·s-1时), 植物的净光合速率和光合有效辐射呈线性相关, 对其光响应曲线进行线性回归, 得软紫草、木本猪毛菜和蒺藜的方程分别为:

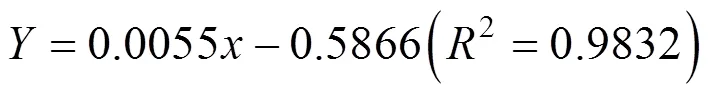
由方程可知此时软紫草、木本猪毛菜和蒺藜的光补偿点分别为66.95 μmol·m-2·s-1、106.65 μmol·m-2·s-1和77.32 μmol·m-2·s-1, 表观量子效率分别为0.016、0.0055和0.0188。
2.2 主要气象因子的日变化特征
2.2.1 光合有效辐射(PAR)与大气温度(Ta)日变化
由图4显示, 该研究区光合有效辐射日变化呈明显的倒“U”型曲线, 9:00较低, 之后逐渐增大, 在14:00左右达到最大值, 为1883.51 μmol·m-2·s-1, 在14:00之后呈下降趋势, 而大气温度在9:00-15:00左右上升较快, 15:00最高, 之后缓慢降低。
2.2.2 大气温度(Ta)和叶片温度(TL)日变化的比较
由图4显示, 三种植物的叶片温度与大气温度变化成正比, 因此叶片温度的变化在一定程度上体现出气温的变化。

图2 三种植物的光响应曲线
Figure 2 Light Response Curve of three plants
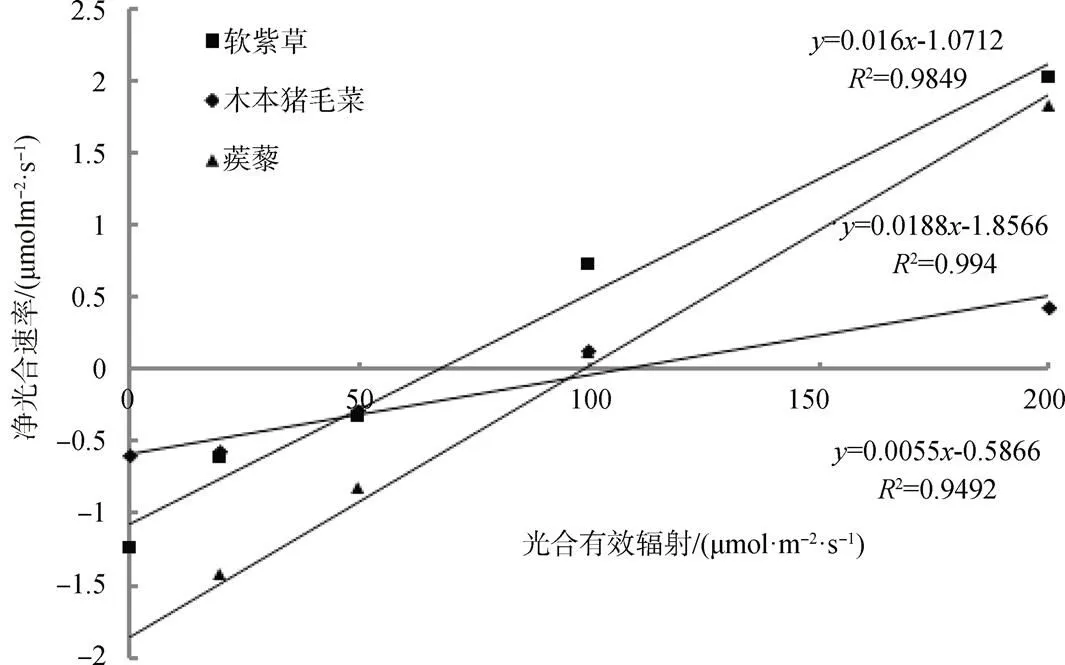
图3 弱光下三种植物的光响应曲线
Figure 3 Light response curve of three plants under low light
2.3 三种植物光合作用参数日变化特征
软紫草和木本猪毛菜叶片净光合速率的日变化呈“双峰型”曲线, “午休”现象明显, 而蒺藜净光合速率在15:00有一短暂波动, 之后呈下降趋势。软紫草和木本猪毛菜叶片的净光合速率峰值出现在11:00和17:00左右, 分别为5.34、6.92 μmol·m-2·s-1和3.74、7.19 μmol·m-2·s-1。而蒺藜的净光合速率峰值出现在9:00左右, 为11.72 μmol·m-2·s-1, 光合“午休”现象不明显如图5(a)。
软紫草、木本猪毛菜叶片的气孔导度日变化与净光合速率变化趋势大体一致, 呈“双峰型”曲线, 这与王连喜, 金鑫[15]的研究结果一致。9:00气孔导度快速升高, 在11:00出现主峰, 后逐渐降低, 在15:00之后随气孔开放度增加, 气孔导度开始增大, 软紫草在17:00出现次峰, 木本猪毛菜在15:00出现次峰, 随后由于气孔进一步关闭, 气孔导度一直呈下降趋势。而蒺藜叶片的气孔导度与净光合速率变化趋势基本一致, 在13:00—17:00变化幅度较大, 9:00左右较高, 后逐渐降低, 在13:00出现低谷, 谷值为0.084 mol·m-2·s-1, 之后随气孔开放度增加, 气孔导度开始上升, 在15:00出现次峰, 峰值为0.216 mol·m-2·s-1, 后由于气孔进一步关闭, 气孔导度又迅速下降且降幅较大, 最小值为0.02 mol·m-2·s-1如图5(b)。
软紫草和木本猪毛菜叶片的胞间CO2浓度呈不规则的“W”型曲线。软紫草低谷出现在11:00和17:00左右, 谷值分别为190.74和136.31 μmol·CO2·mol-1,木本猪毛菜低谷出现在13:00和17:00左右, 谷值分别为126.45和130.59 μmol·CO2·mol-1, 这两种植物在9:00左右, 胞间CO2浓度较高, 之后随着净光合速率的缓慢增加, 胞间CO2浓度逐渐下降, 在11:00—15:00之间, 由于太阳辐射和叶面温度的过高, 植物出现“光合午休”, 气孔导度减小或者完全关闭, 导致净光合速率下降, 胞间CO2升高, 15:00之后又开始下降, 出现次低谷, 其值为136.31 μmol·CO2·mol-1和130.59 μmol·CO2·mol-1, 这与净光合速率变化规律相对应, 即随着净光合速率的升高, 部分CO2消耗, 相对应胞间CO2浓度降低。而蒺藜的胞间CO2浓度大体呈“V”型, 且变化幅度较小。由于受大气CO2的影响, 在早、晚较高, 峰值出现在9:00和19:00左右, 值分别为272.77 μmol·CO2·mol-1和239. 18 μmol·CO2·mol-1, 正午随光强和温度的上升, 开始呈下降的趋势, 在11:00—13:00基本保持平稳的状态, 之后又开始上升, 在19:00达到最大值, 值为239.18 μmol·CO2·mol-1如图5(c)。

图4 三种植物的环境因子日变化
Figure 4 Diurnal variation of environmental factors of three plants
软紫草、木本猪毛菜的蒸腾速率日变化曲线与净光合速率变化趋势基本同步, 均呈“双峰型”曲线, 而蒺藜的蒸腾速率呈不规则的“M”曲线。软紫草的蒸腾速率峰值出现在11: 00和17:00, 为4.26 mmol·m-2·s-1和 5.13 mmol·m-2·s-1, 蒺藜和木本猪毛菜的峰值出现在11:00和15:00左右, 值分别为6.28、7.44 mmol·m-2·s-1和6.29 mmol·m-2·s-1、7.32 mmol·m-2·s-1如图5(d)。
软紫草和蒺藜的水分利用效率日变化均呈“双峰型”曲线, 随时间的推进, 呈先下降后上升再下降的趋势, 峰值出现在9:00和17:00左右, 其值分别为2.58 mmol·mol-1、1.35 mmol·mol-1和2.96 mmol·mol-1、1.77 mmol·mol-1, 软紫草、蒺藜、木本猪毛菜的水分利用率日均值分别为1.24、1.51、1.32 mmol·mol-1如图5(e)。
软紫草、木本猪毛菜的羧化效率日变化呈“双峰型”曲线, 软紫草在11:00和17:00出现峰值, 值分别为0.028和0.05 μmol·m-2·s-1, 木本猪毛菜在13:00和17:00出现峰值, 值分别为0.027和0.021 μmol·m-2·s-1, 蒺藜的羧化效率日变化整体呈下降趋势, 在15:00有一短暂波动, 之后一直呈下降趋势如图5(f)。
软紫草的气孔限制值与羧化效率的日变化基本一致, 蒺藜的气孔限制值呈不规则“V”型, 谷值出现在15:00, 木本猪毛菜的气孔限制值变化幅度较大, 峰值出现在13:00左右如图5(g)。

图5 软紫草、蒺藜、木本猪毛菜的光合作用参数日变化特征
Figure 5 Diurnal variation of photosynthetic parameters of,and
2.4 植物光合参数与环境因子的相关性分析
使用Spss21.0软件对软紫草、蒺藜、木本猪毛菜的净光合速率、气孔导度、胞间CO2浓度, 蒸腾速率等光合参数与环境因子光合有效辐射、大气温度、叶片温度等进行相关性分析。由表1可知, 软紫草在光合参数方面, 气孔导度与净光合速率在0.05水平上呈显著正相关, 胞间CO2浓度与净光合速率、气孔导度在0.01水平上呈显著负相关, 蒸腾速率与气孔导度在0.01水平上呈显著正相关, 与胞间CO2浓度在0.01水平上呈显著负相关。在环境因子方面, 叶片温度与气孔导度在0.05水平上呈显著负相关, 与胞间CO2浓度在0.01水平上呈显著正相关。由此可知: 气孔导度、蒸腾速率对软紫草的光合作用影响最大, 而环境因子中大气温度、空气相对湿度对软紫草影响较大。蒺藜在光合参数方面, 气孔导度与净光合速率在0.01水平上呈显著正相关,胞间CO2浓度与净光合速率在0.05水平上呈显著负相关, 与气孔导度在0.01水平上呈显著负相关, 蒸腾速率与净光合速率、气孔导度在0.01水平上呈显著正相关, 与胞间CO2浓度在0.01水平上呈显著负相关。在环境因子方面, 光合有效辐射与净光合速率在0.05水平上呈显著正相关, 与气孔导度、蒸腾速率在0.01水平上呈显著正相关, 与胞间CO2浓度在0.01水平上呈显著负相关, 叶片温度与气孔导度、蒸腾速率、大气温度在0.01水平上呈显著正相关, 与光合有效辐射在0.05水平上显著正相关。由此可知, 对蒺藜光合作用影响最大的参数为气孔导度, 蒸腾速率。而环境因子如光合有效辐射、叶片温度、空气相对湿度影响最大。木本猪毛菜在光合参数方面, 蒸腾速率与气孔导度在0.01水平上呈显著正相关, 同理: 对木本猪毛菜光合作用影响最大的参数为气孔导度、蒸腾速率, 而与其他环境因子相关性均不显著。
3 讨论
光合作用作为判断绿色植物生长和抗逆性强弱的指标[16], 本研究中净光合速率由大到小依次为蒺藜、软紫草、木本猪毛菜, 且蒺藜具有较高的气孔导度和最大净光合速率, 表明三种植物中, 蒺藜具有较强的光合能力。蒸腾速率受多种因素的影响, 如气候、土壤水分、植物种类等, 因此不同植物表现出不同蒸腾速率的变化[17]。气孔多分布于植物叶片表面, 与外界环境进行气体交换的同时, 还可以适当抑制植物的蒸腾作用。本研究中, 三种植物在9:00-11:00间, 呈上升趋势, 在11:00蒸腾速率达到峰值, 此后随温度的升高而下降, 到15:00、17:00达到第二个峰值, 三种植物蒸腾速率均呈“双峰型”。由于正午光照强度大, 气温高, 蒸腾速率逐渐增大, 导致植物水分供应不足, 为了防止叶片被灼伤, 减少水分的流失, 气孔开放度会减小甚至关闭, 蒸腾速率降低。午后, 由于光强和气温逐渐减弱, 蒸腾作用增强, 导致植物蒸腾速率上升, 最后, 随光合有效辐射的减弱, 气温下降等因素, 蒸腾速率又呈下降的趋势。蒸腾作用在午间降低是荒漠植物通过关闭气孔来防止叶片过度蒸腾, 从而减少水分损耗的一种生态适应对策[18]。水分利用效率是评价植物对环境适应能力的重要指标, 具有重要的生态学意义。在同一种生境中, 水分利用率值越大, 植物的耐旱能力越强[19-21]。在本研究中, 三种植物的耐旱能力大小依次为: 蒺藜、木本猪毛菜、软紫草, 蒺藜更能适应沙区的干旱环境, 是恢复植被、改善生态环境的优良物种, 但一天中三种植物的水分利用率存在时空差异, 其原因可能与净光合产物的积累密切相关[22]。

表1 三种植物各种光合参数与环境因子间的相关性分析
注: **.在0.01水平(双侧)上显著相关。*.在0.05水平(双侧)上显著相关。
自然条件下, 软紫草和木本猪毛菜叶片的净光合速率日变化存在明显的光合“午休”现象, 这与大多数干旱荒漠区植物光合日变化趋势一致[23]。有学者指出, 植物出现“午休”是多种环境因素变化的结果, 如光照、温度及湿度等[24]。由于中午强光, 高温和低湿的原因, 可能会造成净光合速率下降和光合作用减弱。按照Faquhar 和 Sharkey 研究提出的当正午光合速率降低时, 如果胞间CO2浓度随之也呈降低的趋势, 而气孔限制值升高则属于光合作用的气孔限制, 反之, 则为光合作用的非气孔限制的判断原则[25], 软紫草的净光合速率在正午降低时, 胞间CO2浓度呈缓慢上升, 而气孔限制值却呈下降的趋势, 表明引起软紫草正午净光合速率降低的主要原因为非气孔限制, 即随着午间光合作用环境因子的变化, 使得叶肉细胞光合活性下降, 从而引起净光合速率的下降。木本猪毛菜和蒺藜的净光合速率在午间下降时, 胞间CO2浓度也下降, 而气孔限制值升高, 则表明引起木本猪毛菜和蒺藜午休的原因主要为气孔限制因素, 即气孔关闭, 叶肉细胞间隙的CO2减少, 碳源上的限制是引起光合“午休”的主导因素[26]。
通过对三种植物的净光合速率与其生理生态因子进行相关性分析得出, 影响三种植物净光合速率的环境因子, 生理生态因子既有相似性, 又有所差异。综合分析, 可以得出影响三种绿色植物净光合速率的主要影响因子为气孔导度、蒸腾作用、光合有效辐射、空气相对湿度, 而与其他因子相关性不显著。
4 结论
(1)软紫草和木本猪毛菜存在明显的光合“午休”现象, 而蒺藜“午休”现象不明显, 软紫草产生光合“午休”现象主要受非气孔限制因素影响, 而蒺藜和木本猪毛菜主要是气孔限制因素。
(2)在同一种生境下, 植物具有不同的抗旱机制。软紫草与木本猪毛菜选择了较高的净光合速率、蒸腾速率和较低的水分利用效率的生存策略, 属于避旱型。而蒺藜则通过较高的水分利用效率来适应水资源不太丰富的环境, 属于耐旱型。综合来看, 三种植物对矿区生境的适应能力依次为蒺藜、木本猪毛菜、软紫草。
(3)本研究中软紫草和蒺藜具有较低的光补偿点, 较高的光饱和点, 对其生境的光环境具有较强的适应性, 表现在能够利用弱光或者高光强的环境而生长良好。而木本猪毛菜的光补偿点、光饱和点均较高, 属于喜阳植物, 对干旱环境适应性很强, 这三种植物在强光、高温、低湿荒漠环境中都具有一定的生存优势。
[1] 程业森, 赵晨光, 高立平. 阿拉善白刺生长季光合速率及蒸腾速率特征研究[J]. 干旱区资源与环境, 2017, 31(11): 161–168.
[2] 蔡福, 米娜, 纪瑞鹏, 等. 关键生育阶段持续干旱对东北春玉米光合特性的影响[J]. 生态学杂志, 2017, 36(6): 1578–1589.
[3] 叶子飘, 于强. 光合作用光响应模型的比较[J]. 植物生态学报, 2008. 32(6): 1356–1361.
[4] 南江, 赵晓英, 原慧, 等. 霸王和木本猪毛菜在遮风和不遮风环境下的表型特征差异[J]. 生态学报, 2014, 34( 20): 5758–5765.
[5] 文娥, 刘文辉, 宋海龙, 等. 新疆哈密地区野生黄花软紫草的资源调查[J]. 中国现代中药, 2016, 18(11): 1479– 1483.
[6] 张婷, 付卫东, 张瑞海, 等. 蒺藜草入侵对沙质草地氮库的影响[J]. 应用生态学报, 2017, 28(5): 1522–1532.
[7] 唐明. 唐山市工矿区主要园林植物滞尘能力的初步研究[J].湖北农业科学, 2017, (19): 3664–3668.
[8] 高松, 苏培玺, 严巧娣, 等. C4荒漠植物猪毛菜与木本猪毛菜的叶片解剖结构及光合生理特征[J]. 植物生态学报, 2009, 33(2): 347–354.
[9] 付义, 何承忠, 巨苗苗, 等. 基于转录组的疏花软紫草低拷贝核基因引物开发[J]. 西南林业大学学报, 2016, 36(3): 86–90.
[10] 邵麟惠, 郑兴卫, 李聪. 蒺藜苜蓿E3泛素连接酶U-box基因克隆及表达分析[J]. 草业学报, 2016, 25(7): 62–72.
[11] 路桃香, 高天鹏, 薛林贵, 等. 金川矿区重金属污染对土壤微生物分布的影响[J]. 环境科学与技术, 2016(9): 37–44.
[12] 郭自春, 曾凡江, 刘波, 等. 疏叶骆驼刺和多枝柽柳不同时期光合特性日变化及其与环境因子的关系[J]. 西北植物学报, 2015, 35(1): 189–198.
[13] 肖小雨, 龙婉婉, 柳正葳, 等. 吉安地区典型景观湖泊浮游植物群落特征及其与水环境因子的关系[J]. 生态学杂志, 2016, 35(4): 934–941.
[14] 于永畅, 王厚新, 李承秀, 等. 四倍体与二倍体紫薇光合特性研究[J]. 中国农学通报, 2013, 29(22): 10–14.
[15] 王连喜, 金鑫, 李剑萍, 等. 短期高温胁迫对不同生育期马铃薯光合作用的影响[J]. 安徽农业科学, 2011. 39(17): 10207–10210+10352.
[16] 罗青红, 李志军, 伍维模, 等. 胡杨、灰叶胡杨光合及叶绿素荧光特性的比较研究[J]. 西北植物学报, 2006, 26(5): 983–988.
[17] 母贵琴, 张力文, 陈静. 宝兴鼠尾草的光合特性研究[J]. 中国农学通报, 2011. 27(13): 42–48.
[18] 张淑勇, 周泽福, 张光灿, 等. 半干旱黄土丘陵区天然次生灌木山桃()与山杏(L.)叶片气体交换参数日动态差异[J]. 生态学报, 2009, 29(1): 499–507.
[19] 丁俊祥, 邹杰, 唐立松, 等. 克里雅河流域荒漠-绿洲交错带3种不同生活型植物的光合特性[J]. 生态学报, 2015, 35(3): 733–741.
[20] 许大全. 光合作用速率[M]. 上海: 上海科技出版社, 2002.
[21] 赵丹丹, 马红媛, 杨焜, 等. 优质牧草水分利用效率提高技术研究综述[J]. 生态学杂志, 2017, 36(8): 2312– 2320.
[22] 张诚诚, 文佳, 曹志华, 等. 水分胁迫对油茶容器苗叶片解剖结构和光合特性的影响[J]. 西北农林科技大学学报: 自然科学版, 2013. 41(8): 79–84.
[23] 左伟, 马淼, 马智, 等. 荒漠旱生植物刺山柑的光合生理生态学研究[J]. 石河子大学学报(自科版), 2012. 30(3): 282–288.
[24] 余红兵, 曾磬, 肖润林, 等. 花叶芦竹光合特性日动态及其影响因素分析[J]. 湖南农业大学学报(自科版), 2013. 39(2): 200–203.
[25] 蔡金桓, 薛立. 高山植物的光合生理特性研究进展[J]. 生态学杂志, 2018, 37(1): 245–254.
[26] 李征珍, 杨琼, 石莎, 等. 蒙古沙冬青光合作用特征及其影响因素[J]. 生态学杂志, 2017, 36(9): 2481–2488.
Diurnal variation of photosynthetic physiology and ecology of native plants in jinchuan mining area
WANG Yindi1, JU Tianzhen1, ZHOU Yuxia1, WAN Zidong1, FANG Xiangwen5, GAO Tianpeng2,3,4, *
1. College of Geography and Environmental Science, Northwest Normal University, Lanzhou 730070, China 2. Research Center for Pollution Control and Ecological Restoration Engineering of Mining Areas in Gansu Province, Lanzhou 730070, China 3. Ecological and Environmental Field Observation and Research Station of Beilu Mining Area, Qilian Mountains, Jinchang 737100, China 4. School of Biological and Environmental Engineering, Xi’an University, Xi’an710065,China 5.Lanzhou University School of Life Sciences, Lanzhou 730070, China
A portable LI-6400 photosynthesis analyzer was used to select the gas exchange parameters and environment of the leaves ofandgrowing naturally in Jinchuan mining area on sunny days. The factors were measured and the photosynthetic characteristics of the three plants and their relationship with environmental factors were compared to reveal the adaptation characteristics of three plants to the mining environment. The results showed that the diurnal changes of the net photosynthetic rate ofandhad a “noon break” phenomenon. The main reason for the “noon break” ofwas non-stomatal limitation, while that ofthe dominant factor was the stomatal limitation factor. The highest daily peak valuesof net photosynthetic rate ofandall appeared at around 11:00 and 17:00.phenomenon of “noon break in photosynthesis” was not obvious, mainly due to the limitation of stomata, and the maximum peak occurred around 9:00. In short, the water use efficiency of the three plants has spatial and temporal differences, and has certain survival advantages in strong light, high temperature and low humidity desert environment. The adapt ability to the habitat of the mining area is in order of,and.
light response curve; diurnal variation;;;
10.14108/j.cnki.1008-8873.2019.04.004
Q14
A
1008-8873(2019)04-021-08
2018-07-10;
2018-08-13基金项目:国家自然科学基金项目(31860176, 31460162); 甘肃省科技支撑计划(144FKCA086)资助
王引弟(1992—), 女, 硕士研究生, 主要从事植物生理生态学的研究工作, 1721817399@qq.com
高天鹏(1964—), 男, 博士, 教授, 主要从事环境植物学教学与研究工作, E-mail: zkgtp@163.com)
王引弟, 巨天珍, 周玉霞, 等. 金川矿区本土植物光合生理生态日变化特征[J]. 生态科学, 2019, 38(4): 21-28.
WANG Yindi, JU Tianzhen, ZHOU Yuxia, et al. A preliminary study on the diurnal variation of photosynthetic physiological ecology of native plants of Jinchuan mining area[J]. Ecological Science, 2019, 38(4): 21-28.
猜你喜欢
土壤学报(2022年4期)2022-10-22
农业工程学报(2022年3期)2022-04-16
亚太传统医药(2021年9期)2021-10-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
文艺生活·中旬刊(2020年8期)2020-11-23
节水灌溉(2020年9期)2020-09-16
老年教育(2018年12期)2018-12-29
读者·校园版(2018年11期)2018-05-15
现代园艺(2017年19期)2018-01-19
