
广西稻田养殖金边鲤群体遗传多样性分析
2019-09-18 10:29甘宝江韦玲静莫洁琳甘习军滕忠作叶香尘
水产科学 2019年5期
甘宝江,张 盛,韦玲静,黄 杰,莫洁琳,甘习军,滕忠作,叶香尘
( 1.广西水产引育种中心,广西 南宁 530031; 2.柳州渔业技术推广站,广西 柳州 545066;3.三江侗族自治县水产技术推广站,广西 柳州 545500 )
鲤鱼(Cyprinuscarpio)属鲤形目、鲤科、鲤亚科,是一种广泛分布于中国各淡水水域的经济鱼类。鲤鱼的品种繁多,形态各异,包括野生品种[黄河鲤(C.carpiohaematopterus)、黑龙江鲤(C.carpiohaematoperus)、太湖鲤]、育成品种[建鲤(C.carpiovar.jian)、福瑞鲤(C.carpiovar. FFRC)、松浦镜鲤(C.carpiovar.specularis)]及具有地方特色的品种[荷包红鲤(C.carpiovar.wuyuanensis)、兴国红鲤(C.carpiovar.singuonensis)]。这些丰富的鲤鱼种质资源库,拥有众多优良的经济性状(高产、抗逆、耐寒等),可作为鲤鱼品种选育和改良的重要资源库。
我国的稻田养鱼历史悠久,可分为传统养殖、探索发展、现代养殖3个阶段[1],在这期间稻田养鱼的生产模式由“以稻为主”向“以鱼为主”转变,促使稻田鱼类养殖产量逐年飙升,根据《2017年中国渔业统计年鉴》的统计数据显示,2016年稻田养殖面积达1 516 093 hm2,比去年同比增长0.96%,稻田养成鱼产量达1 632 263 t,比去年同比增长4.75%。稻田养鱼将稻田种养和鱼类养殖相结合不仅可以减少农药化肥的使用,保证谷物和鱼肉安全、卫生,还可以极大限度地利用稻田资源,缓解人地冲突,保证谷物丰收及增加农民收入。目前,我国稻田养殖鱼类主要以鲤鱼为主,鲫鱼(Carassiusauratus)、鲢鱼(Hypophthalmichthysmolitrix)、草鱼(Ctenpharyngodonidellus)等为辅,其中禾花鲤(C.carpiorubrofuscus)以其肉质口感细嫩、味道鲜美深受养殖户和消费者的青睐。金边鲤(C.carpiovar.Jinbian)[2]是由广西水产引育种中心联合多个单位以柳州融水苗族自治县的融水田鲤为基础群体,针对稻田养殖模式而选育的鲤新品系,该鱼体色大部分呈青灰色,背鳍两侧延伸至头部和尾部的体色呈连续或断续金色,宽度为0.2~0.5 cm,该鱼在水中时,似有一条“金边”镶嵌在鱼的背部,因此得名。金边鲤具有身型短小、肤黄腹圆、性情温顺、起捕率高、鱼肉品质高等特点,而且洪水暴发时,该品种的鱼身居塘底不易随洪水游走,减少稻田养殖鱼类流失的风险,因此被认为非常适用于稻田养殖生产。近几年金边鲤已陆续覆盖云南、贵州、四川、广东等地的水产养殖业,其推广快,经济价值高,市场前景不可估量。然而,金边鲤品种遗传信息背景仍是一片空白,为更好地开展稻田金边鲤品种的选育和改良工作,其遗传背景的研究意义重大。
优质的亲本资源是鱼类选育和遗传改良的基础,然而盲目、缺乏科学的繁殖计划必将造成严重的损失,如群体遗传多样性下降、经济性状丢失、经济效益降低等。使用遗传标记辅助育种不仅可以快速、准确地对物种遗传信息进行评估,而且还可在科学指导下进行生产工作。其中,微卫星标记作为第二代遗传标记,具有基因组中数量众多、分布均匀、共显性、多态性高及遗传稳定等特点,已被应用于鲤的构建遗传图谱[3]、群体遗传分析[4]、亲权鉴定[5]及分子辅助育种[6]等遗传学研究领域;并且相对其他遗传标记(如同工酶、RAPD、RFLP等),微卫星标记更适合用于群体遗传分析、系谱认证、品种及亲子鉴定等研究[7-8]。笔者通过选取10个高度多态的微卫星标记,利用高分辨率的全自动DNA测序仪分析技术为检测手段[9],以野生、池塘和稻田养殖鲤鱼群体遗传变异现状作为参照对金边鲤群体遗传结构进行分析,旨在对金边鲤遗传信息背景进行初探,并为进一步开展金边鲤的遗传改良和保护提供群体遗传学参考信息。
1 材料与方法
1.1 试验材料
本研究共7个鲤鱼群体,包括2个野生群体(黑龙江鲤、太湖鲤)、3个池塘养殖群体(兴国红鲤、福瑞鲤、建鲤)及2个稻田养殖群体(金边鲤、晒江鲤)。各群体样本于2015—2018年分别采集于黑龙江流域(黑龙江鲤,n=22)、江苏无锡太湖(太湖鲤,n=36)、江西兴国县(兴国红鲤,n=36)、中国水产科学研究院淡水渔业研究中心(福瑞鲤,n=32)、广西南宁市武鸣区(建鲤,n=32)、广西柳州市融水苗族自治县(金边鲤,n=34)和广西柳州市三江侗族自治县(晒江鲤,n=32)(表1)。每尾样本鱼剪取尾鳍约0.5 g浸泡于无水乙醇中保存备用,采用经典的酚—氯仿法提取基因组DNA[10]。
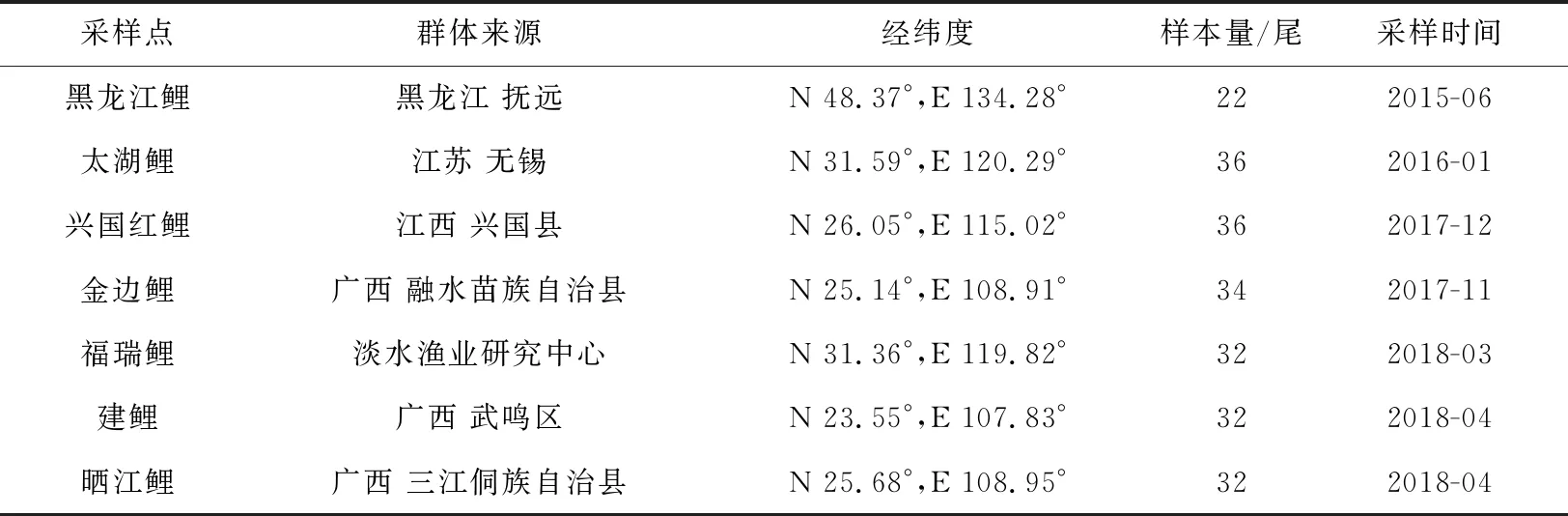
表1 本研究中所用的鲤鱼样本信息
1.2 引物选取及PCR扩增
本研究中所使用10对微卫星标记选自HLJ系列、XG系列和CCE系列(共500对)中高度多态且扩增稳定的标记(表2)。PCR反应总体积为15 μL:36~60 ng模板DNA,0.48 U Taq DNA聚合酶,1.5 μL 10×PCR buffer,0.48 μL dNTP(2.5 mmol/L),0.48 μL正反向混合引物(各2.5 μmol/L),11.22 μL灭菌去离子水。PCR扩增条件:94 ℃预变性5 min;扩增32个循环,每个循环94 ℃变性35 s,56~60 ℃退火35 s(退火温度见表2),72 ℃延伸40 s;72 ℃终延伸10 min。微卫星荧光引物由北京六合华大基因科技有限公司合成,每对微卫星标记的正向引物5′端使用FAM或HEX荧光染料进行标记,PCR扩增产物由武汉天一辉远生物科技有限公司的3730型DNA测序仪(ABI,USA)进行测序分型。
1.3 数据统计及分析
使用GeneMarker 2.2软件读取3730型DNA测序仪测序结果后进行数据汇总。利用软件PopGene 32计算等位基因数、有效等位基因数、观测杂合度、期望杂合度以及Nei′s遗传距离等遗传参数[11]。Microsoft-toolkit插件用于计算多态信息含量。使用MEGA 4软件[12]基于7个鲤鱼群体Nei′s遗传距离参数进行聚类分析。Arlequin 3.1软件[13]计算各群体间的遗传分化系数及进行哈迪—温伯格平衡检验。进行多重比较时,概率的显著性水平需要进行Bonferroni校正[14]。利用分子变异分析方法[15]计算群体遗传结构,利用Structure 2.2软件[16]中的贝叶斯方法[17]混合模型来构建鲤鱼群体遗传结构图,根据公式计算出每个K值所对应的ΔK值,当ΔK值最大时所对应的K值即为最佳理论群体数。
ΔK=[L″(K)]/s[L(K)]
式中,K为群体数,L(K)为各K值对应lnP(D)的均值,L′(K)=L(K)-L(K-1),|L″(K)|=|L′(K+1)-L′(K)|,s[L(K)]为L(K)的标准差。
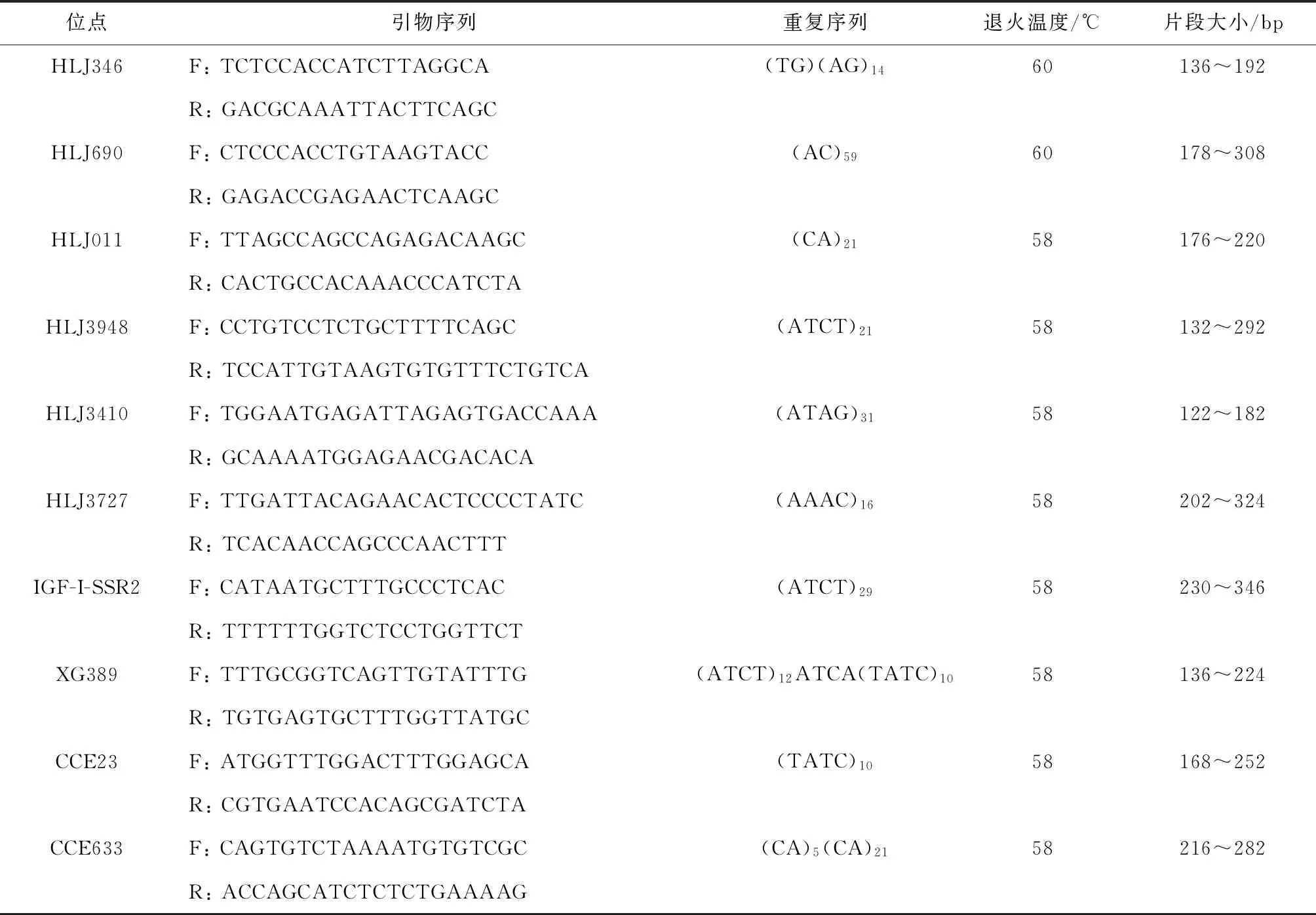
表2 鲤鱼微卫星标记的基本信息
2 结 果
2.1 微卫星位点在鲤鱼群体中的群体遗传学信息
10个微卫星位点在7个鲤鱼群体中遗传参数信息见表3。研究结果显示,等位基因数在HLJ3727位点最小(16),在HLJ690位点最大(48)。有效等位基因数为6.4~21.3。10个微卫星标记的期望杂合度、观测杂合度以及多态信息含量分别为0.846~0.955、0.475~0.897、0.829~0.951,表明这些标记均为高度多态性位点(多态信息含量>0.5)。7个群体在各位点的遗传多样性信息见表4。在7个鲤群体中共检测到242个等位基因。每个群体的平均等位基因数和平均有效等位基因数分别为6~16和3.8~9.4,平均期望杂合度、平均观测杂合度以及平均多态信息含量为0.736~0.900、0.620~0.800、0.689~0.878(表5),其中兴国红鲤群体各项平均遗传参数较低。野生群体(黑龙江鲤、太湖鲤)、稻田养殖群体(金边鲤、晒江鲤)和池塘养殖群体(兴国红鲤、福瑞鲤、建鲤)分别检测到302、258和311个等位基因(表4)。由表4、表5可见,7个鲤鱼群体均具有较高的遗传多样性,平均期望杂合度、平均观测杂合度、平均多态信息含量等遗传多样性参数由高至低分别为野生群体、稻田养殖群体和池塘养殖群体。
哈迪—温伯格平衡检验显示(表4),除了CCE23位点外,其余9个位点在7个群体中的等位基因频率显著偏离哈迪—温伯格平衡(P<0.05)。其中,黑龙江鲤、太湖鲤、兴国红鲤、金边鲤、福瑞鲤、建鲤、晒江鲤群体分别在5个位点(HLJ346、HLJ690、HLJ011、HLJ3727、IGF-I-SSR2)、4个位点(HLJ346、IGF-I-SSR2、HLJ011、CCE633)、3个位点(HLJ346、HLJ011、CCE633)、5个位点(HLJ346、HLJ690、HLJ011、HLJY3948、IGF-I-SSR2)、6个位点(HLJ346、HLJ690、HLJ011、HLJ3948、XG389、CCE633)、3个位点(HLJ690、HLJ011、CCE633)和2个位点(HLJ690、IGF-I-SSR2)极显著偏离哈迪—温伯格平衡(P<0.01)。整体水平上,野生群体在6个位点极显著偏离哈迪—温伯格平衡,而稻田养殖群体和池塘养殖群体分别在5个和7个位点极显著偏离哈迪—温伯格平衡。由此可见,本研究中池塘养殖群体偏离哈迪—温伯格平衡位点最多。

表3 10个微卫星标记的遗传参数信息
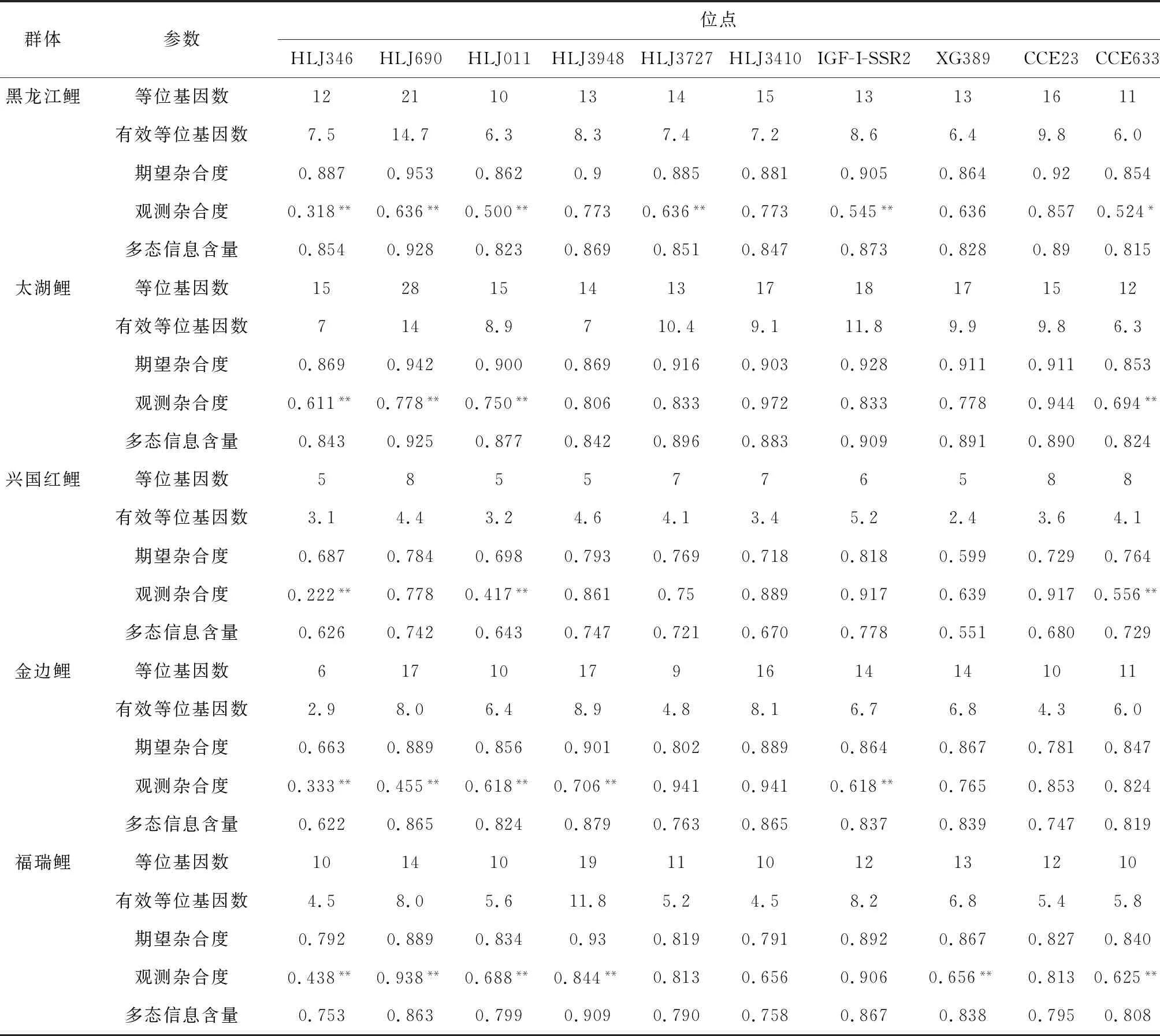
表4 7个鲤鱼群体在10个微卫星位点的多态性信息
续表4
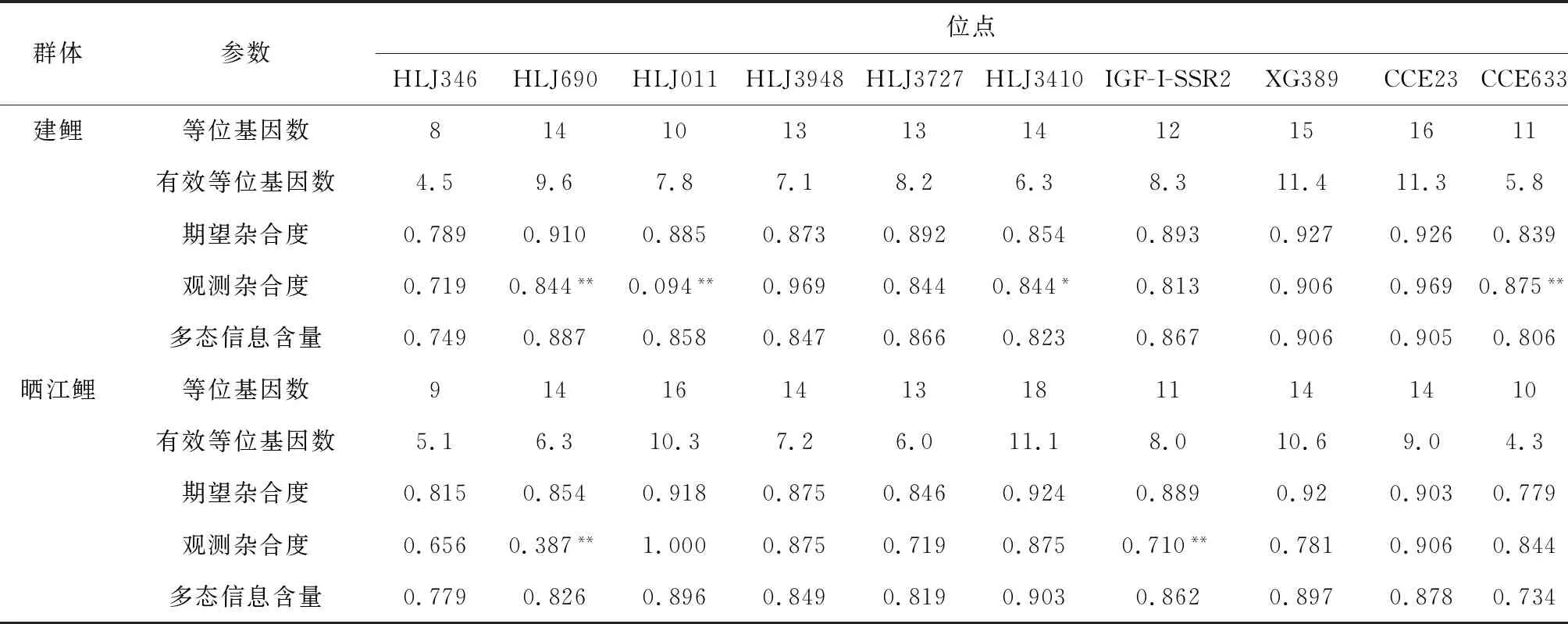
群体参数位点HLJ346HLJ690HLJ011HLJ3948HLJ3727HLJ3410IGF-I-SSR2XG389CCE23CCE633建鲤等位基因数8141013131412151611有效等位基因数4.59.67.87.18.26.38.311.411.35.8期望杂合度0.7890.9100.8850.8730.8920.8540.8930.9270.9260.839观测杂合度0.7190.844**0.094**0.9690.8440.844*0.8130.9060.9690.875**多态信息含量0.7490.8870.8580.8470.8660.8230.8670.9060.9050.806晒江鲤等位基因数9141614131811141410有效等位基因数5.16.310.37.26.011.18.010.69.04.3期望杂合度0.8150.8540.9180.8750.8460.9240.8890.920.9030.779观测杂合度0.6560.387**1.0000.8750.7190.8750.710**0.7810.9060.844多态信息含量0.7790.8260.8960.8490.8190.9030.8620.8970.8780.734
注:*表示位点显著偏离哈迪—温伯格平衡(P<0.05),**表示位点极显著偏离哈迪—温伯格平衡(P<0.01).
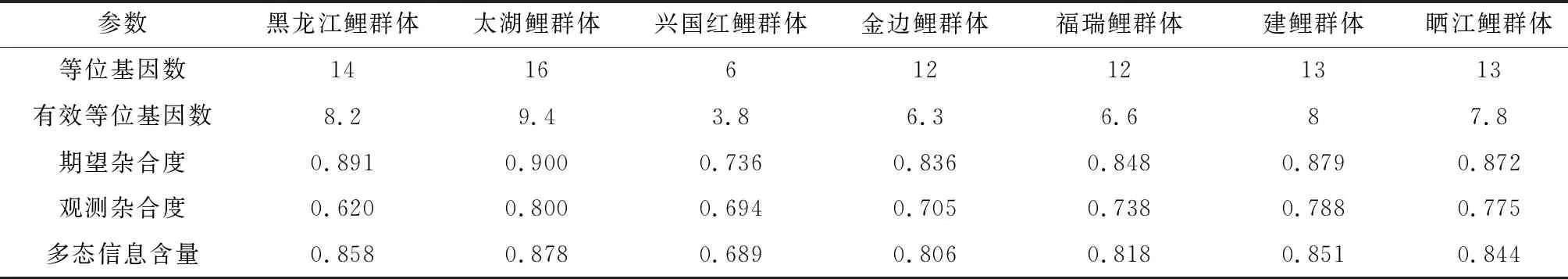
表5 7个鲤鱼群体的平均遗传参数
2.2 群体间的遗传分化
7个鲤鱼群体遗传分化系数为0.015~0.141(表6),存在不同程度的遗传分化。其中,黑龙江鲤与太湖鲤、建鲤、晒江鲤,太湖鲤与建鲤、晒江鲤,晒江鲤与金边鲤、建鲤群体间遗传分化系数为0.015~0.046(遗传分化系数<0.05),遗传分化均极显著(P<0.01),表明这些群体间出现了轻微程度的遗传分化。而其余群体两两间遗传分化系数均处于0.05~0.15间,遗传分化均极显著(P<0.01),表明这些群体间属于中等程度的遗传分化。AMOVA分析结果显示,来自群体间的遗传变异占7.15%,而群体内的遗传分化占92.85%,达到极显著水平(P<0.01),表明鲤鱼群体主要的遗传分化来自于群体内部。利用软件Structure 2.2对鲤鱼种群进行的遗传结构分析。本研究选取K值为2~7,重复计算20次,根据公式ΔK=[L″(K)]/s[L(K)]计算得出ΔK最大时K=4(图1)。当K取值为4时,贝叶斯分类结果显示(图2),7个鲤鱼群体分属于4个类群,其中,黑龙江鲤群体、太湖鲤群体和建鲤群体与其他鲤鱼群体间存在明显的遗传分化,划分为一个类群;金边鲤群体和晒江鲤群体较其他群体遗传结构分化明显,划为一个类群;剩余两个池塘养殖兴国红鲤群体、福瑞鲤群体各自独立划为一个类群。
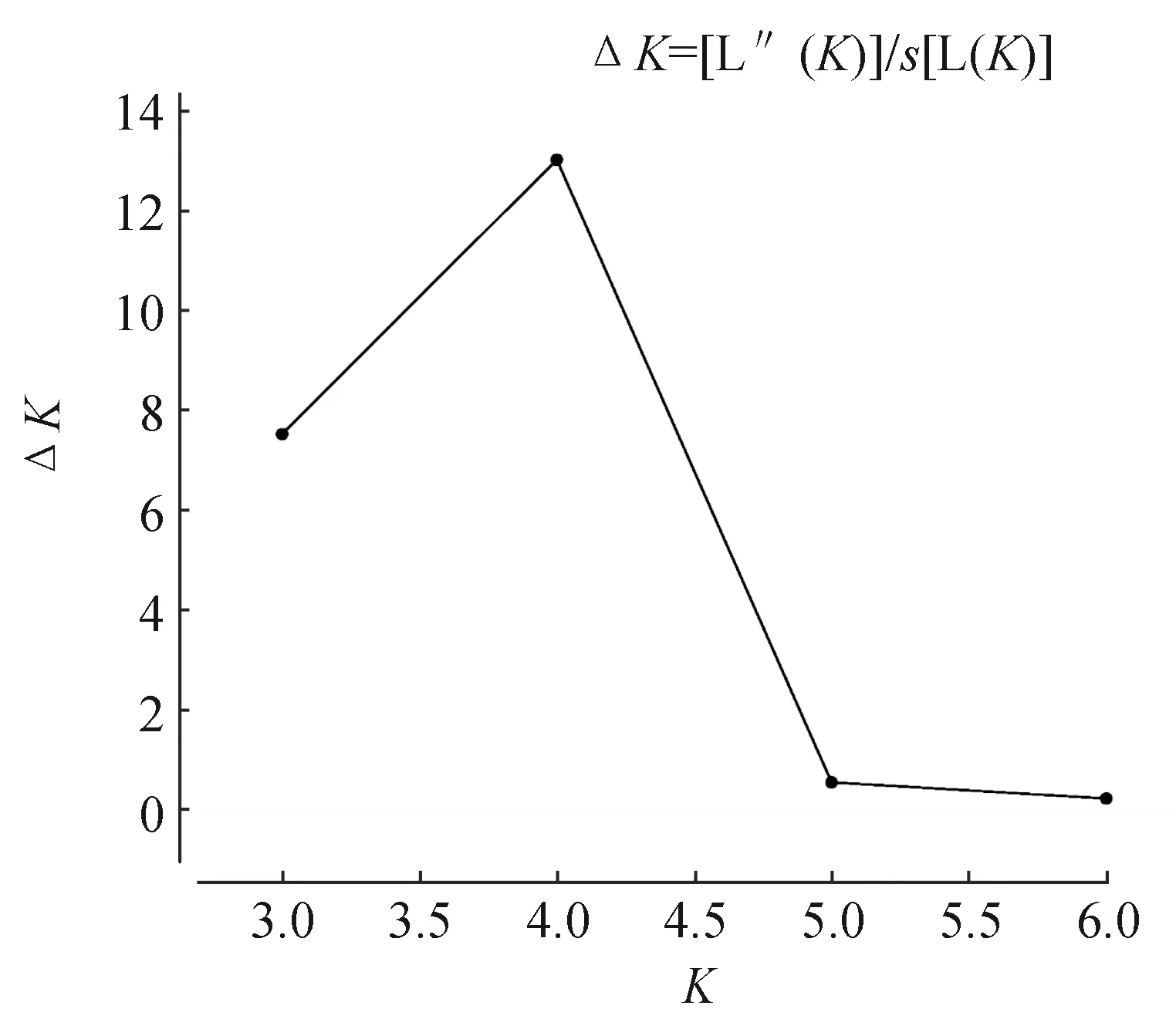
图1 ΔK在不同K值时的计算结果
各个鲤鱼群体间的Nei′s遗传距离为0.286~0.977(表6),其中以黑龙江鲤与太湖鲤群体之间的遗传距离最小,而兴国红鲤与福瑞鲤群体之间的遗传距离最大。据Nei′s遗传距离构建7个鲤群体的聚类树,结果表明(图3),黑龙江鲤与太湖鲤两个野生群体聚为一支后再与建鲤群体聚为一大支;稻田养殖金边鲤和晒江鲤群体聚为的一支;兴国红鲤以及福瑞鲤群体再分别依次与其聚类。聚类结果与贝叶斯分类结果相符。除金边鲤和晒江鲤群体地理位置距离近而聚为一类外,其余各群体间遗传距离与群体间的地理距离没有直接联系。

图2 7 个鲤鱼群体的遗传组分分析(K=4)
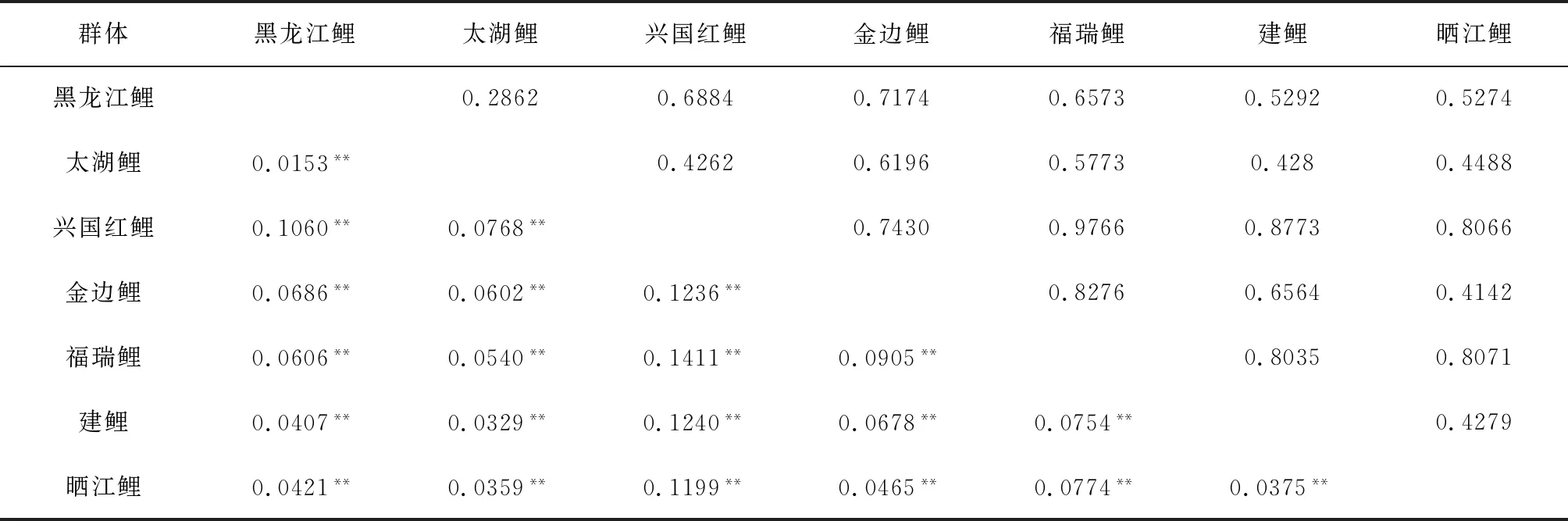
群体黑龙江鲤太湖鲤兴国红鲤金边鲤福瑞鲤建鲤晒江鲤黑龙江鲤0.28620.68840.71740.65730.52920.5274太湖鲤0.0153**0.42620.61960.57730.4280.4488兴国红鲤0.1060**0.0768**0.74300.97660.87730.8066金边鲤0.0686**0.0602**0.1236**0.82760.65640.4142福瑞鲤0.0606**0.0540**0.1411**0.0905**0.80350.8071建鲤0.0407**0.0329**0.1240**0.0678**0.0754**0.4279晒江鲤0.0421**0.0359**0.1199**0.0465**0.0774**0.0375**
注:**表示极显著变化(P<0.01).
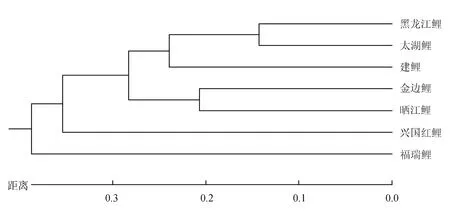
图3 基于Nei′s遗传距离构建的7个鲤鱼群体的UPGMA聚类树
3 讨 论
3.1 适用于检测鲤群体遗传结构的微卫星标记
在水产动物遗传多样性研究中,试验样本数量和微卫星标记数量及类型,均会对研究结果的各遗传参数产生一定的影响。研究表明,在使用微卫星检测群体遗传多样性试验中,当群体样本量大于30时,各遗传参数变化幅度逐渐变小并趋于稳定,得到的试验结果可靠性高[18-19]。当使用的微卫星标记量为5~15个时,各遗传参数有较大的浮动,一般达到25个之后才逐步趋于稳定,同时有研究也表明,使用标记数为10个时,各遗传参数数值呈现最低水平[18-19]。微卫星标记的多态性基于微卫星核心重复单元及其重复次数[20]。其中,微卫星的重复单元为2~6碱基,数量以二碱基为主,扩增稳定性属三、四碱基最高,而扩增稳定性越高则可以更忠诚地反映物种的遗传信息[21-22];微卫星的重复次数与其多态性之间成正比例关系[23]。本研究中所使用的试验材料,除黑龙江鲤群体外,其余群体样本量均大于30个,达到最低样本量要求;所使用的微卫星标记的核心重复序列绝大多数为四碱基,重复次数均在10次以上(表3),可更准确地映射出物种的遗传信息;而本研究使用10个微卫星标记可用于检测群体各遗传参数的下限,在一定程度上会对试验结果造成一定影响,但用于群体遗传多样性的初探,不仅可以省时、省力还可以节约成本,而且该结果还可为进一步的研究做好铺垫。
10个SSR标记在7个鲤鱼群体中,共检测出242个等位基因,每个微卫星位点遗传参数显示,等位基因数、期望杂合度、观测杂合度以及多态信息含量分别为15~45,0.851~0.954,0.390~0.881,0.833~0.949(表3),均为高度多态性位点。鲁翠云等[24]在福瑞鲤中检测到平均有效等位基因数、观测杂合度、期望杂合度和多态信息含量分别为4.559、0.695、0.741和0.702,说明该群体处于高度多态水平(多态信息含量≥0.5);曾繁振等[25]对3个鲤鱼品种进行遗传多样性分析得到,荷包红鲤、兴国红鲤和长江野鲤群体的平均多态信息含量分别为0.6253、0.6969和0.7775。二者的研究结果与本研究结果相似。而Li等[26]在研究6个鲤鱼野生群体时发现,各个群体的平均多态信息含量仅为0.44~0.64;鲁翠云等[27]用鲤鱼EST-SSRs分子标记检测长江及黑龙江鲤群体结构时,得到的各遗传参数平均有效等位基因数、观测杂合度、期望杂合度和多态信息含量分别为3.9135、3.4820,0.6067、0.5667,0.6737、0.6194和0.6308、0.5714。上述后二者的研究结果与本研究结果相比均偏低。分析造成研究结果差异的几个因素:第一,微卫星类型。本研究中所使用的微卫星标记核心序列重复次数均大于10,而鲁翠云等[24]微卫星标记核心序列重复次数最少为7。第二,等位基因的检测方法(分辨率)。不同检测方法检测微卫星的多态性时会产生差异,传统方法(聚丙烯酰胺凝胶电泳)分辨率低,而DNA测序仪检测仪分辨率高,后者检测到的位点数更多[28-29]。最后,微卫星位点在基因组中的位置。理论上来说,EST-SSR标记多态性较基因组微卫星(G-SSR)标记的要低,在动植物遗传多样性研究中均被报道[30-32]。G-SSR和EST-SSR标记分别来源于基因组的非编码区段和编码区内或靠近编码区[33]。物种在进化过程中,位于编码区的EST-SSR标记相对稳定、变异率低[34],而G-SSR则相反。微卫星的重复单元呈现一定规律,EST-SSR标记多为二核苷酸重复的微卫星,G-SSR标记的核心重复单元则包括二核苷酸和多核苷酸,本研究微卫星重复单元以四碱基重复单元为主,而鲁翠云等[24,26]所使用的微卫星标记多以二碱基重复单元为主。综上所述,本研究中检测到7个鲤鱼群体具有较高的遗传多样性,稻田鲤群体可提供丰富的多态性位点,是优质的育种材料;所使用的微卫星位点能提供较丰富遗传信息,是理想分子遗传标记,适用于检测鲤鱼群体遗传多样性及遗传结构。
3.2 稻田养殖群体遗传多样性及群体间的比较

在本研究中另一个值得注意的是,兴国红鲤养殖群体的各项遗传参数指标均明显低于野生群体,该群体共检测到64个等位基因,并且在每个位点的等位基因数均明显少于其他群体,等位基因数丢失高达63.1%。类似的现象在其他物种中也有报道,Li等[43]利用微卫星研究皱纹盘鲍(Haliotisdiscushannai)群体时发现,养殖群体的等位基因数丢失率高达76%,推测原因为,基础群体少从而加剧遗传漂变现象的发生;Skaala等[44]在研究大西洋鲑(Salmosalar)时发现,养殖群体杂合度虽然无明显变化,但却丢失了42%的等位基因,其推测原因可能为奠基者效应。本研究中,兴国红鲤群体的各遗传参数与上述两种研究结果较为相似(等位基因数=6,期望杂合度=0.736,观测杂合度=0.694),推测其可能受到奠基者效应、遗传瓶颈效应和人为选择的影响。其余群体(金边鲤、晒江鲤、福瑞鲤和建鲤)的杂合度较野生群体偏低,但却不明显,推断这些养殖群体的基础群体遗传多样性丰富,有效亲本量大,繁殖过程较为科学。本研究中养殖群体中有8个位点经Bonferroni校正后,仍极显著偏离哈迪—温伯格平衡,可能为养殖群体近亲杂交、人工选择等致使群体遗传结构发生改变。综上所述,稻田金边鲤群体具有较高的杂合度、等位基因丰度和遗传多样性水平,在遗传育种和改良过程中可作为优质育种材料,应合理开发和利用。
3.3 稻田鲤群体遗传进化
遗传分化系数值可作为反映群体间遗传分化程度的参考指标。本研究中7个鲤鱼群体的遗传分化分析结果表明,群体间存在不同程度的遗传分化,其中金边鲤群体与晒江鲤群体间遗传分化属低等程度[45](0<遗传分化系数<0.05),与其余群体间遗传分化达到中等程度[45](0.05<遗传分化系数<0.15)。本研究以野生、池塘和稻田养殖群体作为参照,对金边鲤群体进行遗传分化分析,其中,黑龙江鲤和太湖鲤可作为野生群体的代表;建鲤和福瑞鲤是育成品种,而兴国红鲤则是具有悠久养殖历史的特色地理品种;稻田群体晒江鲤与金边鲤均是广西柳州地区具有地方特色的稻田养殖品种。建鲤的育种基础群中母本为荷包红鲤,父本为元江鲤(C.carpioyuankiang)[46];兴国红鲤源于江西省,在当地已有1300多年的养殖史[47],与荷包红鲤和玻璃红鲤(C.carpiovar.wananensis)均是优良的鱼类种质资源,常用作亲本杂交出许多有较高经济价值的杂交种,许多杂交鲤组合几乎均含有兴国红鲤或荷包红鲤的遗传因子,如丰鲤、荷元鲤、三杂交鲤、建鲤等;福瑞鲤是中国水产科学研究院淡水渔业研究中心以野生黄河鲤和建鲤为基础群体,通过1代群体选育后,连续4代BLUP家系选育而获得的品种[48];金边鲤原产于广西柳州市融水苗族自治县,晒江鲤原产于广西柳州市三江侗族自治县,两者均是具有地方特色的稻田养殖品种。因此,由以上信息可知,黑龙江鲤群体来源于黑龙江水系,太湖鲤、兴国红鲤群体来源于长江水系,金边鲤、晒江鲤群体来源于珠江水系,建鲤群体来源于长江水系与元江水系杂交,福瑞鲤群体来源于长江水系、元江水系及黄河水系。一般情况下,相同生态环境中的鲤亚科鱼类间存在频繁的基因交流现象,因此,杂交品种与杂交亲本相同水系的鲤群体间具有一定的同源性,进行聚类分析时会聚为一类,李建林等[37]对6个鲤鱼群体遗传差异的研究结果也支持该推测。而本研究中群体聚类分析和遗传结构结果显示,除珠江水系的两个稻田养殖品种金边鲤和晒江鲤群体遗传结构较为相似且聚为一类外,其余群体进行聚类的结果与溯源水系相关性较差。分析造成此类结果差异的原因:首先,隶属广西的两个稻田金边鲤和晒江鲤养殖群体起源地属于相同水系,在地理位置上比较接近,相距约100 km,两个群体间存在一定程度的基因交流。其次,金边鲤群体基础群体为野生群体,其基础群体数量大且繁殖代数少;晒江鲤群体繁殖亲本散布在各个养殖户,在繁殖季节到来时,养殖户将优质亲本共享,相互交换用于配对繁殖子代,因而大大降低了奠基者效应和遗传漂变发生的几率。最后,鱼类的杂交子代继承亲本遗传信息具有不均衡性[49],杂交种群经过几个世代的人工繁殖,遗传结构将会产生不定向的变化。本研究还发现,除金边鲤群体外,其余群体间存在不同程度的基因交流。推测鱼类品种经过人工选育、世代累进、封闭繁殖、遗传瓶颈效应、奠基者效应等因素,导致相同品种的鱼类在不同地区养殖后遗传结构发生明显改变,亲缘关系也发生相应的变化。综上所述,本研究使用不同地区、不同养殖模式及不同鲤鱼品种作为参照,可更好的对金边鲤种群的遗传信息进行合理的评估,7个鲤鱼群体遗传进化分析结果表明,金边鲤群体与野生和池塘养殖群体间遗传分化明显,而这些差异也造就了自身群体的特点(身型短小、肤黄腹圆、性情温顺、起捕率高、鱼肉品质高等)更适合稻田养殖模式,它可作为稻田鲤品种优质的育种材料。
3.4 小结
本研究通过利用野生、池塘和稻田养殖鲤鱼群体遗传多样性现状作为参照,可更好地对金边鲤群体遗传信息进行评估,研究表明,7个鲤鱼群体均呈现出较高的遗传多样性,其中稻田金边鲤群体遗传信息表明其均具有良好的育种潜力,可为稻田鲤品种遗传育种和改良提供优良的种质资源。然而该群体遗传多样性相较野生群体已呈现出下降趋势,推测养殖群体易受人为干涉影响,应引起有关人员的重视。另一方面,本研究中的兴国红鲤养殖群体遗传多样性衰退严重,提示鱼类原种种质资源应该要合理的保护和利用,并及时利用遗传分子标记对鱼类群体遗传结构进行监测,以补充优质亲本资源,防止人工养殖过程中奠基者效应和遗传瓶颈效应的发生。本研究旨在对金边鲤的遗传学信息背景进行初探,同时丰富稻田鲤种质资源数据库以及为稻田鲤新品种选育与遗传改良提供参考信息。
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
水产科学(2022年2期)2022-03-20
南京师范大学学报(工程技术版)(2021年2期)2021-10-20
种子(2021年3期)2021-04-12
环球人物(2020年13期)2020-07-13
孩子(2020年4期)2020-06-08
中国-东盟博览(旅游版)(2018年10期)2018-05-14
校园英语·下旬(2017年7期)2017-07-14
科技视界(2016年27期)2017-03-14
