
乙醇促进法夫酵母虾青素合成的机理及其代谢调控
2019-09-04 09:53沈宁燕陈艳红肖安风
中国食品学报 2019年8期
茹 毅 沈宁燕 倪 辉,2,3,4 陈艳红,2,3,4 肖安风,2,3,4*
(1 集美大学食品与生物工程学院 福建厦门361021 2 福建省食品微生物与酶工程重点实验室 福建厦门361021 3 福建省海洋功能食品工程技术研究中心 福建厦门361021 4 厦门市海洋功能食品重点实验室 福建厦门361021)
虾青素(Astaxanthin),又名3,3,-二羟基-β羟β′-胡萝卜素-4,4′-二酮,是一种具有超强抗氧化活性的类胡萝卜素[1,2]。虾青素可以有效抑制细胞的氧化损伤,降低机体的突变和癌变,同时还具有抗高血压,预防心血管疾病,增强免疫,防紫外线辐射等诸多功效[3]。虾青素在食品、饲料、保健品和医药等行业的应用与日俱增,如日本将虾青素与蓝莓提取物组配,以强化对视力的保护效果[4]。美国Cyanotech 公司推出的Derma Astin(黛玛)天然虾青素胶囊具有抗衰老功能[4]。作为饲料添加剂的虾青素促进观赏鱼类的着色[5]等。
现在虾青素的生产方法主要包括化学合成、天然提取以及生物合成。其中化学合成的虾青素由于稳定性、抗氧化活性以及安全性不如天然提取的虾青素,所以应用较少[6]。目前天然虾青素来源主要是藻类和酵母,藻类有单细胞绿藻雨生红球藻,酵母有法夫酵母。法夫酵母产虾青素的相关研究包括菌种改良[7-9],培养基优化[7-9]以及虾青素合成代谢调控[10-12]。Meyer 等[13]发现虾青素的合成是生长关联型,平衡期时菌体浓度迅速下降,而细胞内虾青素含量上升。Liu 等[14]研究发现在酵母培养前期加入适量的H2O2,虾青素的细胞产率和体积产率分别提高83%和65%。
研究表明虾青素的合成与法夫酵母的代谢途径的关键节点调控有关。Gu 等[15]发现添加0.2%的乙醇能有效提高虾青素的产量,可能是由于提高了虾青素合成过程的关键途径——甲羟戊酸合成途径中HMG-CoA 还原酶的活性。肖安风等[16]发现培养基中添加低浓度的乙酸钠、乳酸钠和柠檬酸(糖代谢产物),均能增强法夫酵母产虾青素的能力。本实验室前期研究了葡萄糖浓度、添加乙醇及溶氧状况对法夫酵母细胞生长、代谢副产物产生的影响及其与虾青素合成之间的关系,并据此阐释法夫酵母虾青素合成的机理[12]。在此基础上通过代谢通量分析方法,阐述乙醇促进虾青素的合成机理,然后利用代谢调控剂对法夫酵母虾青素合成的代谢调控节点进行发酵调控,从而达到促进法夫酵母合成虾青素的目的。
1 材料和方法
1.1 菌株
法夫酵母(Phaffia rhodozyma)JMU-MVP14 菌株:由出发菌株JMU-668 经甲基磺酸乙酯诱变,再用0.5%的双氧水结合紫外照射,产生自由基,从而淘汰低产菌株,最后选育得到菌株JMUMVP14[12]。
1.2 培养基
种子培养基:4°Bx 麦汁,pH 6.0。
发酵培养基(/L):葡萄糖0.5 g,(NH4)2SO45 g,MgSO40.5 g,KH2PO42 g 采 用121 ℃灭 菌15 min;微量成分ZnSO4·7H2O 12 mg,CaCl210 mg,FeSO4·7H2O 8 mg,H3BO32.67 mg,MnCl22.67 mg,Na2MoO4·2H2O 1.07 mg,CuSO4·5H2O 0.8 mg,Co-Cl20.8 mg,KI 0.27 mg,肌醇66.67 mg,泛酸钙2.67 mg,烟酸2.67 mg,VB62.67 mg,VB12.67 mg,对氨基苯甲酸0.53 mg,生物素0.13 mg,pH 6.0 mg,采用过滤除菌的方法。
1.3 法夫酵母发酵
取-70 ℃甘油管保存的菌种,在装有4°麦汁培养基的斜面上划线,22 ℃恒温培养5 d。挑取斜面上的单菌落,接入250 mL 的摇瓶中,培养2~3 d,为1 代种子。转接1 代种子到新配制的种子培养基中培养2~3 d,将培养好的2 代种子按5%接种量接入装有30 mL 发酵培养基的250 mL 摇瓶中,22 ℃,180 r/min 发酵5 d。
1.4 生物量的测定
干重法:将空的离心管于105 ℃干燥箱烘至恒重,称重为M;取一定体积的发酵液(V)于恒重后的离心管中,离心两次后得到菌体,于105 ℃干燥箱中烘干至恒重,称重为N,得到菌体干重为(N- M)/V[17]。
1.5 总类胡萝卜素含量测定
采用二甲基亚砜(DMSO)法破壁[18-19]。取5 mL发酵液,离心两次后得到菌体。将菌体55 ℃下预热5 min,加入2 mL 已预热至75 ℃的二甲基亚砜,充分振荡后加入5 mL 乙醇提取色素,离心后得到色素提取液,用乙醇定容10 mL。
采用紫外分光光度计检测[20]。配制不同浓度梯度的虾青素标准品,在波长474 nm 处检测,绘制出虾青素标准曲线。在相同条件下测定发酵样品,根据标准曲线求得样品中虾青素含量。
1.6 类胡萝卜素成分的测定
使用Aglient 1200 高效液相色谱仪(G1315 DAD 检测器)检测,Nova-Pak C18 柱(3.9 mm×150 mm,4 μm),液相分析时控制流速1 mL/min,柱温30 ℃,柱压0~20 MPa,进样量20 μL,检测波长474 nm[21]。
1.7 发酵液残糖的检测
采用3,5-二硝基水杨酸法(DNS)[22]测定残糖浓度。将1 mL 发酵上清液稀释至适宜浓度,加入0.6 mL DNS,沸水浴10 min 后立即冷却,用蒸馏水定容5 mL,使用紫外分光光度计在波长为520 nm 处测定糖浓度。
1.8 发酵液乙醇含量的检测
气相色谱法测定发酵液中的乙醇[17]。
1.9 代谢通量分析

图1 法夫酵母代谢网络Fig.1 The metabolic reaction network of Phaffiarhodozyma
根据文献[23]~[25]建立法夫酵母生物合成虾青素的简化代谢网络(图1)。该代谢网络主要是由糖酵解途径(EMP)、磷酸戊糖途径(HMP)、三磷酸循环(TCA)和代谢物积累等4 个部分组成。代谢流平衡模型的建立和代谢通量分布计算见文献[23]~[25]。
2 结果与讨论
2.1 代谢通量分析乙醇促进虾青素合成的机理
有研究表明法夫酵母由中心途径经异戊二烯合成虾青素的代谢途径[26],然而,对虾青素的合成机理及代谢调控机制研究不够深入。法夫酵母具有复杂的代谢网络,其合成虾青素由多个代谢网络节点所调控,每个节点的有益改造只能使虾青素的产量得到少量提高。虾青素产量的不断提高,实际是各代谢调控节点有益改造并不断累积的过程。要使法夫酵母的虾青素产量不断提高,就必须深入了解法夫酵母合成虾青素的代谢调控机理,这是对法夫酵母的代谢流进行理性改造,提高虾青素发酵产量的关键。
由于法夫酵母JMU-MVP14 细胞代谢不产生乙醇,因此配制葡萄糖质量浓度为20 g/L 的代谢通量分析培养基。在接种前,额外添加2 g/L 的乙醇作为试验组,以不加乙醇的培养条件作为对照组进行代谢通量分析,考察乙醇对整个法夫酵母JMU-MVP14 代谢网络的扰动规律。根据细胞的生长特性,分别在发酵对数期与稳定期取样,测定发酵参数后代入代谢通量的模型中进行计算。法夫酵母JMU-MVP14 的发酵曲线如图2所示。
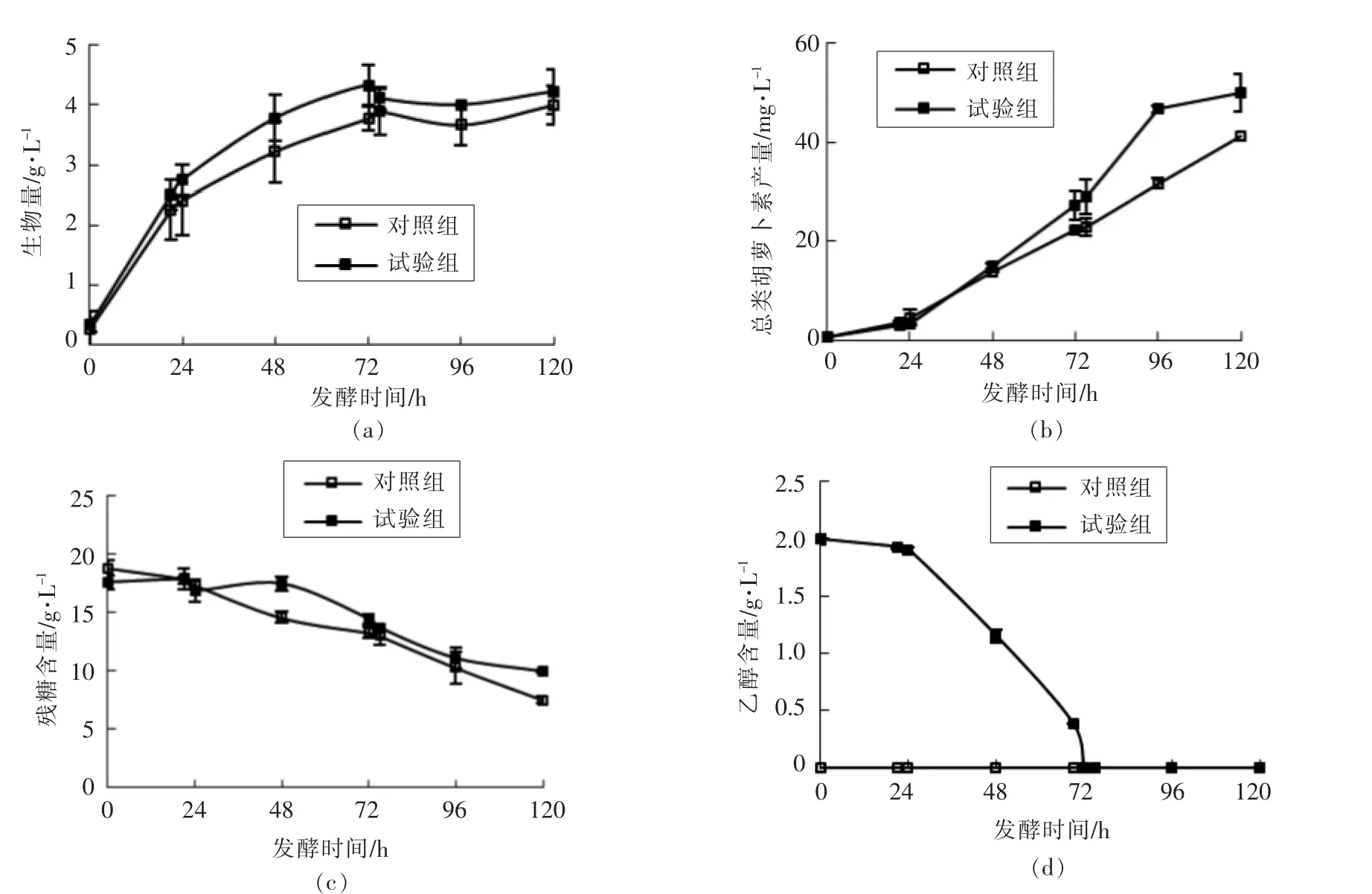
图2 法夫酵母JMU-MVP14 的发酵曲线Fig.2 Fermentation curve of Phaffiarhodozyma JMU-MVP14
从图2可以看出,在代谢通量分析培养基中额外添加乙醇,能显著提高法夫酵母JMU-MVP14菌株合成色素的能力。从生物量的变化曲线可知,法夫酵母JMU-MVP14 菌株在代谢通量培养基中生长快速,生物量发酵至72 h 时达到最大值。以这一时间点为分割,之前是细胞生长的对数期,之后是生长稳定期。当法夫酵母处于对数生长期时,能快速利用葡萄糖来合成新的细胞,而类胡萝卜素的合成较为缓慢。进入稳定期后,菌体量基本不增加,而胞内色素的合成却持续进行。从总类胡萝卜素的产量曲线可以看出,乙醇对虾青素产量的促进作用在生长稳定期较为显著。发酵至72 h,生物量达到最大值,而类胡萝卜素还在以对数的趋势积累。在此阶段进行代谢调控,将更有可能提高虾青素的产量。取72~75 h 时间段的样品进行代谢通量分析,得到图3。图中斜杠右边数据为代谢添加乙醇培养的试验组,斜杠左边数据为不添加乙醇培养的对照组。

图3 法夫酵母JMU-MVP14 菌株代谢通量分布图(72~75 h)Table 3 Metabolic flux distribution of Phaffiarhodozyma JMU-MVP14 in the period of 72~75 h
法夫酵母JMU-MVP14 菌株在生长的不同时期,各途径的代谢通量存在较大差异。由图1可知,r2和r3为进入磷酸戊糖途径的通量,主要产生5-磷酸核酮糖和NADPH,参与核酸的代谢并为细胞内大分子物质的合成提供还原力;r4为进入糖酵解途径的通量,主要产生丙酮酸及一些中间产物,参与其它途径的代谢;r9则可用来表示进入三羧酸循环的通量,能产生大量ATP 供细胞生长所利用,其中间产物α-酮戊二酸还可参与氨基酸的代谢;r8为乙酰辅酶A 的通量,可作为三羧酸循环和虾青素合成途径的前体物;r12为从乙酰辅酶A合成虾青素的通量。
Gu 等[15]研究发现,将乙醇添加到培养基中,能提高法夫酵母乙醇脱氢酶的活性。在乙醇脱氢酶的作用下,乙醇被氧化成乙醛,乙醛进一步转化成乙酰辅酶A 进入代谢网络中被法夫酵母所利用。从法夫酵母JMU-MVP14 菌株代谢通量分布图(72~75 h)对比图3可知,添加乙醇后法夫酵母JMU-MVP14 菌株代谢网络中流向虾青素合成途径的通量增加,是对照组通量的2.3 倍。而最终测定的虾青素产量比对照组提高了21%。添加乙醇后除了增大丙酮酸、乙酰辅酶A 代谢节点处的通量外,还增大了α-酮戊二酸处的通量,使法夫酵母JMU-MVP14 菌株将更多的碳源流向TCA 循环而获得大量ATP,同时α-酮戊二酸还可参与氨基酸的代谢合成蛋白质。
2.2 α-酮戊二酸代谢节点的调控
细胞代谢过程中对α-酮戊二酸的利用通常有两个方面:一是合成琥珀酰-CoA,参与TCA 循环;二是合成谷氨酸、脯氨酸、精氨酸等,参与蛋白质的代谢。而细胞内蛋白质的合成对虾青素的积累有显著的影响[27]。由上述试验可知,添加乙醇可以提高乙酰辅酶A 的代谢通量,同时使α-酮戊二酸增加,进而提高虾青素产量。结合代谢通量分析与关键代谢途径的通量对比,推测α-酮戊二酸影响虾青素的合成。为了验证推测,配制不同浓度的α-酮戊二酸,添加到法夫酵母的培养基中进行发酵,研究α-酮戊二酸对虾青素合成的影响。
培养环境中添加0.5 g/L 的α-酮戊二酸,有利于法夫酵母JMU-MVP14 细胞的生长,促进总类胡萝卜素的合成,如图4所示。然而,添加1 g/L 及以上质量浓度时对类胡萝卜素的合成不利。
通过高效液相分析法夫酵母JMU-MVP14 菌株的类胡萝卜素成分和含量,从表1可知,细胞内的主要色素是虾青素,占总色素的83%左右。除此之外,还有β-隐黄质和β-胡萝卜素生成。当培养液中添加α-酮戊二酸后明显提高了细胞内β-隐黄质的含量,由8.8%提至18%~28%,而虾青素与β-胡萝卜素的比例有所下降,这可能是由于α-酮戊二酸影响虾青素合成酶的活性,导致虾青素在总色素中的比例下降。

图4 不同α-酮戊二酸添加量对JMU-MVP14 发酵的影响Fig.4 The effect of α-ketoglutaric on fermentation of Phaffiarhodozyma JMU-MVP14

表1 α-酮戊二酸对法夫酵母JMU- MVP14 胞内类胡萝卜素比值的影响Table 1 The effect of α-ketoglutaric on endocellular carotenoid proportion of Phaffiarhodozyma JMU-MVP14
2.3 5-磷酸核酮糖代谢节点的调控
根据代谢通量分析,增强5-磷酸核酮糖代谢节点处的通量,即增强代谢网络中HMP 途径的碳流量,可能会提高虾青素的产量。在乙醇添加试验中,发现HMP 途径的通量值r2减小而TCA 循环的通量值r9增大,这与相关文献报道有所出入[25],其原因可能是乙醇会抑制6-磷酸葡萄糖脱氢(G6PDH)酶的活性。G6PDH 是HMP 途径的限速酶,若提高此酶的活性,则能有效提高HMP 途径的通量。Lan 等[28]研究表明,适宜浓度的谷氨酸能提高G6PDH 的活性。在发酵培养基中添加不同浓度的谷氨酸进行法夫酵母摇瓶发酵。
谷氨酸对法夫酵母JMU-MVP14 生产虾青素的促进效果更为显著。从图5可知,添加0.5~3 g/L的谷氨酸能促进虾青素的合成,且随着谷氨酸浓度的增大,得到的菌体量和色素产量越高。以3 g/L 谷氨酸添加量培养JMU-MVP14,最终收获6.93 g/L 的菌体量,相应总类胡萝卜素的产量为67.88 mg/L,是对照组的1.7 倍。这是由于法夫酵母合成1 个虾青素分子就需要消耗24 个NADPH,而胞内大量NADPH 的来源是HMP 途径。谷氨酸促进HMP 途径的通量后,将提供给虾青素合成更多的NADPH 及能量。
JMU-MVP14 菌株在生产虾青素的同时,还会合成β-胡萝卜素、β-隐黄质。如表2所示,通常情况下,JMU-MVP14 细胞内虾青素占总类胡萝卜素的83.05%。当其培养环境加入谷氨酸后,胞内类胡萝卜素的比例发生变化,β-隐黄质从7.79%提至25%左右,而虾青素和β-胡萝卜的比例有所下降。由此可知,不同培养条件影响法夫酵母胞内色素的比例。

图5 不同谷氨酸添加量对JMU-MVP14 发酵的影响Fig.5 The effect of glutamic on fermentation of Phaffiarhodozyma JMU-MVP14

表2 谷氨酸对法夫酵母JMU-MVP14 胞内类胡萝卜素比值的影响Table 2 The effect of glutamic on endocellular carotenoid proportion of Phaffia rhodozyma JMU-MVP14
3 结论
利用代谢通量分析的方法,阐释法夫酵母JMU-MVP14 生物合成虾青素的调控机理并对代谢流进行发酵调控。虾青素合成试验结果表明:添加2 g/L 乙醇,能促进丙酮酸、乙酰辅酶A 代谢节点的通量,从而增强流向虾青素的代谢通量,虾青素产量达到49.93 mg/L,与对照组相比提高了21%。根据代谢通量分析结果,发现添加乙醇使TCA 循环处的代谢通量值r9增大,使流向乙酰辅酶A 合成虾青素的通量变少。另外,添加α-酮戊二酸会抑制法夫酵母合成虾青素,当添加浓度较高时出现强烈的抑制作用。添加α-酮戊二酸后显著促进法夫酵母JMU-MVP14 菌株细胞内β-隐黄质的合成量,比空白对照组提高18%~28%。谷氨酸作为HMP 途径限速酶(G6PDH)的促进剂,添加后,法夫酵母JMU-MVP14 的总类胡萝卜素产量高达67.88 mg/L,是对照组的1.7 倍。
猜你喜欢
中南民族大学学报(自然科学版)(2022年4期)2022-07-01
中国饲料(2021年17期)2021-11-02
当代水产(2021年6期)2021-08-13
食品与发酵工业(2021年4期)2021-03-01
落叶果树(2021年6期)2021-02-12
三农资讯半月报(2020年6期)2020-04-10
妈妈宝宝(2018年4期)2018-11-29
健康博览(2018年9期)2018-01-04
中成药(2017年6期)2017-06-13
中国调味品(2017年2期)2017-03-20
