多胺代谢与自噬在增龄大鼠心脏中的变化及外源性精脒对衰老心脏自噬的影响
2019-08-20 01:40:00吴飞翔陈俞翰李邵琦王俊莹赵雅君
中国药理学通报 2019年8期
吴飞翔,陈俞翰,李邵琦,王 举,林 岩,王俊莹,张 昊,于 雪,赵雅君
(1. 哈尔滨医科大学基础医学院病理生理学教研室,黑龙江 哈尔滨 150081;2. 齐齐哈尔医学院基础医学院病理生理学教研室,黑龙江 齐齐哈尔 161006)
心脏衰老的机制复杂,近年来,自噬在心脏衰老中的作用备受关注[1]。自噬是在多种基因相互作用下完成的复杂细胞过程,是维持细胞内环境稳态的重要机制。每种自噬相关基因(ATG)负责调控自噬体启动与形成的不同阶段,其中ATG12-ATG5缀合物对自噬的中心事件自噬体的形成至关重要,ATG7是活化ATG8(即自噬标志性蛋白LC3)和ATG12的关键上游分子,ATG7的活化与ATG12-ATG5缀合物形成均是自噬的重要调控环节[2]。p16基因是一种抑癌基因,也是细胞衰老的重要调控基因。p16与pRb-cyclinD1-CDK4形成G1期检查站点,抑制细胞周期G1-S期的转化。导入p16基因的细胞可出现衰老表型[3]。目前,在多种组织器官衰老的模型中发现,自噬减少与疾病发展和寿命缩短密切相关,而增强细胞自噬可以延长寿命[4]。但是,自噬在心脏衰老中的作用报道较少,有待于进一步研究。
多胺(polyamine,PAs)包括精胺(spermine,SP)、精脒(spermidine,SPD)等,广泛存在于真核生物组织细胞内,具有促进细胞增殖、抗炎、抗氧化、抗凋亡、诱导自噬等广泛生物学作用[5]。细胞内多胺水平对维持细胞稳态非常重要,细胞内多胺水平受合成、分解及转移出胞等多种机制的精确调控。鸟氨酸脱羧酶(ornithine decarboxylase,ODC)为多胺合成代谢限速酶,精脒/精胺乙酰转移酶(spermidine/spermine N1-acetyltransferas,SSAT)为多胺分解代谢限速酶[5]。已有文献报道,多胺代谢失衡导致许多细胞过程受损,引起组织器官损伤;而提供外源性精脒对多种模式生物,包括小鼠、大鼠的衰老及衰老相关疾病具有保护作用[6]。但是年龄依赖的大鼠心脏多胺代谢与自噬的变化规律及二者的关系,目前还不清楚。本研究对此进行探讨,同时进一步观察外源性精脒对老龄大鼠心肌细胞自噬、氧化应激及细胞凋亡的影响。
1 材料与方法
1.1 实验动物Wistar大鼠,♂,清洁级,购自哈尔滨医科大学附属第二临床医学院动物实验中心,动物生产许可证号:SCXK(黑)2013-001。饲养条件:给予充足水和标准饲料,维持室温(20~22) ℃,并保持光照充足。青年组鼠为3月龄,老年组鼠为22~24月龄,精脒组鼠为腹腔注射精脒6周的22~24月龄大鼠。
1.2 试剂精脒(Sigma公司);抗GAPDH抗体(10494-1-AP)、抗p62/SQSTM1抗体(18420-1-AP)、抗p16抗体(10883-1-AP)、抗ATG5抗体(10181-2-AP),均购自Proteintech公司;抗LC3-Ⅰ/Ⅱ抗体(#4108)、抗ATG7抗体(#8558)购自Cell Signaling Technology公司;抗ODC抗体(sc-33539)、抗SSAT抗体 (sc-67159)购自Santa Cruz公司;BCA蛋白浓度测定试剂盒(P0006C)、超氧化物阴离子荧光探针二氢乙锭(dihydroethidium,DHE)试剂盒购自上海碧云天生物技术研究所;TUNEL试剂盒(11684817910),购自Roche公司。
1.3 仪器倒置显微镜(日本Nikon公司);H-600型透射电子显微镜(日本日立);MCO-17AI CO2培养箱(日本SANYO);电泳仪、电泳槽(美国Bio-Rad);FluorChemR化学发光仪(美国Thermo公司);S-1300-U净化工作台(苏州安泰空气技术公司);Fluoview FV1000荧光显微镜(日本奥林巴斯光学有限公司)。
1.4 动物实验随机取3、6、12、24月龄Wistar ♂大鼠(n=12),应用水合氯醛(300 mg·kg-1)麻醉大鼠,同时进行肝素化处理(1 000 IU·kg-1),大鼠仰卧位固定于手术台上,胸前区备皮,开胸后迅速取出心脏,冷PBS冲洗出心脏残余血液,拭干心脏并称重,取左心室肌组织用于免疫印迹分析。另一组实验,随机取3月龄Wistar ♂大鼠为青年组(Young组,n=6);24月龄♂大鼠为老年组(Old组,n=6);24月龄♂大鼠每天腹腔注射精脒(10 mg·kg-1),连续6周,设为精脒给药组(SPD组,n=6)。大鼠麻醉后,摘取心脏称重用于后续实验。
1.5 免疫印迹法检测蛋白表达取各组大鼠左心室肌组织,液氮中研磨后,加入含1% PMSF的裂解液,制成心肌组织匀浆,4 ℃、1 500×g离心15 min,取上清,BCA法测定蛋白含量。经SDS-PAGE分离目的蛋白,将蛋白转移到PVDF膜上,脱脂牛奶室温封闭2 h,孵育一抗4 ℃冰箱过夜;TBST洗膜3次后,加入二抗室温孵育2 h;TBST清洗3次,加入ECL发光试剂显色成像,分析条带密度。
1.6 HE染色取大鼠左心室肌组织(厚度小于0.5 cm)在4%多聚甲醛溶液中固定24 h后,石蜡包埋,依次脱蜡、水化、苏木精与伊红(HE)染色、脱水、透明、封片,制成HE染色切片。光学显微镜下观察心肌形态结构。随机选取8个视野,采用Image-Pro Plus软件进行心肌细胞横截面积测量。
1.7 TUNEL分析取上述方法制备的石蜡包埋的心肌组织切片,采用TUNEL法检测心肌细胞凋亡。按试剂盒说明书操作,光镜下,凋亡的心肌细胞核染成棕褐色,每张切片随机取5个以上的高倍视野,计数不少于200个心肌细胞核,计算凋亡指数(apoptosis index,AI)/%=阳性细胞数/(阳性细胞核数+阴性细胞数)×100%。
1.8 DHE荧光探针测定心肌细胞ROS生成取制备好的心肌组织冰冻切片,加入DHE(5 μmol·L-1)工作液到待染心肌冰冻切片上,37 ℃避光孵育30 min,用PBS洗涤细胞3次,在荧光显微镜下观察ROS的荧光强度。
1.9 透射电子显微镜观察心肌超微结构及自噬小体形成取大鼠左心室心肌组织1.0 mm×1.0 mm×1.0 mm,置于2.5%戊二醛磷酸缓冲液中,4 ℃固定24 h以上。常规脱水,透明,包埋并染色,制成50~70 nm超薄切片,在透射电镜下观察心肌超微结构变化及心肌细胞内自噬小体的生成,并拍照。

2 结果
2.1 大鼠心肌ODC、SSAT、LC3-Ⅱ/LC3-Ⅰ和p62蛋白表达的增龄性变化Western blot检测3、6、12、24月龄大鼠左心室肌组织中心肌多胺合成代谢关键酶ODC与多胺分解代谢关键酶SSAT的蛋白表达,以及自噬相关蛋白p62、LC3-Ⅱ/LC3-Ⅰ蛋白表达。Fig 1结果显示,ODC呈年龄依赖的表达下降,SSAT呈年龄依赖的表达增加。与3月龄比较,24月龄大鼠心肌组织ODC蛋白表达明显下降(P<0.05),SSAT蛋白表达明显增多(P<0.05)。LC3-Ⅱ/LC3-Ⅰ表达比值也呈年龄依赖地下降,p62表达呈年龄依赖性增加。与3月龄比较,24月龄心肌组织LC3-Ⅱ/LC3-Ⅰ比值明显下降(P<0.01),p62蛋白表达明显增多(P<0.01)。
2.2 年龄依赖的大鼠心肌多胺代谢变化与细胞自噬水平变化的相关性分析如Fig 2所示,年龄依赖的(3、6、12、24月龄)大鼠心肌ODC表达与LC3-Ⅱ/LC3-Ⅰ比值呈明显正相关(r2= 0.338 4,P<0.05),与p62表达呈明显负相关 (r2= 0.386 6,P<0.05);SSAT蛋白表达与LC3-Ⅱ/LC3-Ⅰ比值呈明显负相关(r2= 0.344 4,P<0.05),与p62表达呈明显正相关(r2= 0.388 6,P<0.05)。
2.3 外源性多胺对老龄大鼠心肌细胞自噬的影响透射电镜检测心肌超微结构变化及心肌细胞自噬小体。Fig 3结果显示,青年组心肌肌丝排列整齐、肌节结构清晰,肌丝间线粒体排列紧密,嵴清晰、膜结构完整、基质致密,并可见自噬小体,未见脂褐素颗粒沉积。老年组大鼠心肌肌丝排列较整齐,线粒体基质密度下降、部分线粒体可见肿胀甚至呈空泡变性,可见脂褐素颗粒沉积,未见双层膜的自噬体;精脒干预组肌丝排列整齐,线粒体基质致密,脂褐素颗粒少见,自噬小体形成增多。大鼠心肌组织中自噬相关蛋白LC3-Ⅱ/LC3-Ⅰ、p62、ATG5和ATG7表达结果显示(Fig 4),与青年组相比,老年组心肌p62蛋白表达明显增加(P<0.05),LC3-Ⅱ/LC3-Ⅰ比值及ATG5、ATG7蛋白表达明显减小(P<0.05);给予外源性精脒处理的老龄大鼠心肌(SPD组)p62蛋白表达明显减少(P<0.05),LC3Ⅱ/Ⅰ、ATG5和ATG7蛋白表达明显增加(P<0.05)。
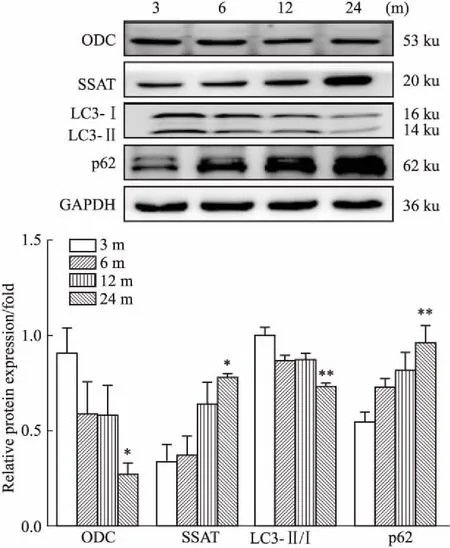
Fig 1 Expression of ODC, SSAT, LC3-Ⅱ/LC3-Ⅰ and p62 protein in cardiac tissues from 3-, 6-, 12-
*P<0.05,**P<0.01vs3 months
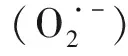
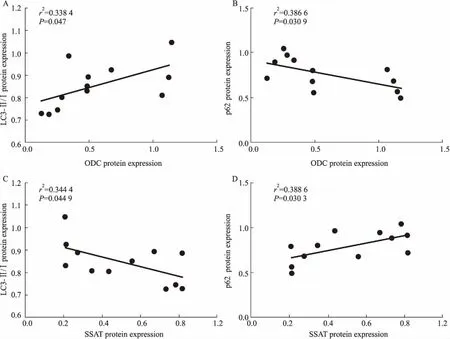
Fig 2 Relationship between expression of ODC and LC3-Ⅱ/LC3-Ⅰ (A) and ODC and p62 (B); and expression of SSAT and LC3-Ⅱ/LC3-Ⅰ ratio (C), and SSAT and p62 (D) in cardiac tissues from 3-, 6-, 12- and 24-month-old rats by scatter plots (n=12)

Fig 3 Effect of SPD on ultrastructure and autophagosome in myocardium of aged rats (×10 000)
The left ventricular ultrastructure was observed by transmission electron microscopy.
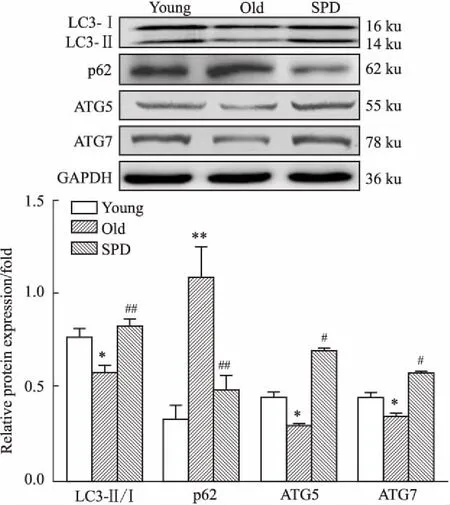
Fig 4 Effect of SPD on autophagy in myocardium in aged rats n=6)
*P<0.05,**P<0.01vsyoung group;#P<0.05,##P<0.01vsold group
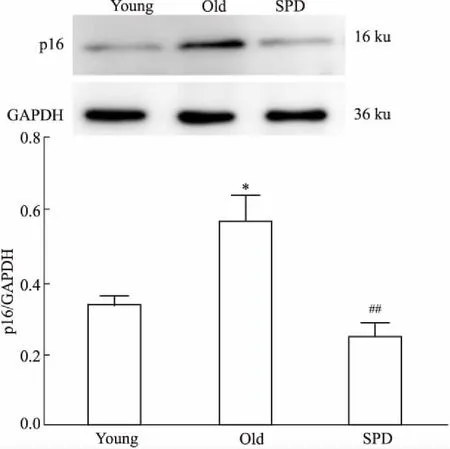
Fig 5 Expression of p16 protein measured by Western blot n=6)
*P<0.05vsyoung group;##P<0.01vsold group
3 讨论
本研究结果显示,随着年龄增加,多胺合成代谢关键酶ODC表达下调,多胺分解代谢关键酶SSAT表达上调,这将会导致心肌多胺总水平降低。细胞内的多胺具有促进细胞增殖、分化、调控细胞内钙、抗炎、抗氧化等广泛的生物学作用,对细胞稳态的维持非常重要[5]。我们前期研究发现,适度运动训练通过上调老龄大鼠心肌多胺合成代谢,维持心肌多胺总水平,恢复了老年大鼠心肌对缺血预适应刺激的敏感性[6]。因此,我们推测,由多胺代谢失衡导致的心肌多胺池耗损可能参与了心脏衰老过程。Nishimura等[7]检测3、10和26周龄的♀小鼠14种不同组织中的多胺含量,发现14种组织中,有11种组织中的精脒水平降低。研究显示,随着年龄增加,组织器官多胺水平降低,ODC活性也呈相同下降趋势,相反,增殖旺盛的组织细胞ODC活性增加,多胺水平增加[8]。因此,多胺可能在心脏衰老及衰老相关心血管疾病中发挥重要作用。
微管相关蛋白轻链1轻链3(microtubule-associated protein 1 light chain 3,LC3)是酵母ATG8蛋白的哺乳动物同源体,有I和II两种形式。LC3-Ⅰ定位于胞质,LC3-Ⅱ定位于自噬体的双层膜上。自噬发生时,自噬体与溶酶体融合,LC3-Ⅱ被溶酶体中的水解酶降解,因此,LC3-Ⅱ含量或LC3-Ⅱ/LC3-Ⅰ比值能够反映自噬活性。p62作为适配子蛋白,在细胞内诱导镶嵌有LC3的自噬体到溶酶体,将其吞噬并清除,与底物结合的p62也被蛋白水解酶降解,p62水平升高通常被认为是自噬活性受到抑制的标志[1]。本研究观察到,3、6、12、24月大鼠心肌LC3-Ⅱ/LC3-Ⅰ比值呈年龄依赖性下降,p62蛋白表达呈年龄依赖性的增加,说明随着年龄增加,大鼠心肌自噬水平下调。我们同时观察到,3、6、12、24月大鼠心肌的多胺代谢合成代谢关键酶ODC蛋白表达与LC3-Ⅱ/LC3-Ⅰ比值呈正相关,与p62蛋白表达呈负相关;多胺分解代谢关键酶SSAT表达与LC3-Ⅱ/LC3-Ⅰ比值呈负相关,与p62蛋白表达呈正相关。近期已有研究证实,心肌细胞依赖于包括自噬的线粒体质量控制机制维持细胞活力, 自噬能力下降是心脏衰老的重要机制之一[1]。本研究也证实,衰老大鼠心肌自噬水平随着年龄增加而降低,同时, 衰老心肌自噬功能下降可能与多胺代谢的增龄性变化存在内在联系,二者可能相互作用,共同促进了心脏老化进程。
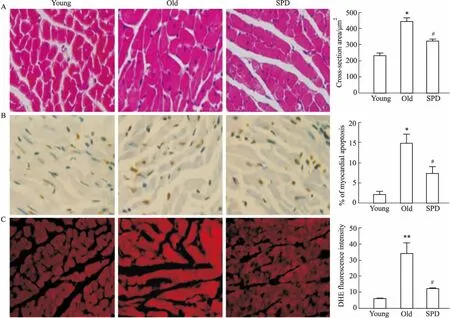

A: Representative left ventricle tissue sections stained by HE (×400), and quantification of the cross-section of cardiomyocytes; B: Nuclei with brown staining indicate TUNEL-positive cells (×400), and apoptosis rate in myocardial cells; C: ROS production was detected by DHE fluorescence staining (×400), and ROS levels in myocardial cells.*P<0.05,**P<0.01vsyoung group,#P<0.05vsold group.
我们给予老年24月龄大鼠补充外源性精脒处理6周,发现精脒能明显抑制衰老诱导的心肌LC3-Ⅱ/LC3-Ⅰ、ATG5及ATG7蛋白表达下调,以及p62表达上调,增加衰老心肌细胞自噬小体形成,提示精脒能够促进衰老大鼠心肌细胞自噬。此外,研究发现,精脒处理能明显抑制24月龄大鼠心脏p16蛋白表达,降低衰老诱导的心肌细胞横截面积增大、心肌细胞凋亡及心肌ROS生成,提示精脒具有延缓大鼠心脏的老化作用。
目前广泛认可的观点是氧化应激及线粒体功能障碍是心脏衰老的重要机制。线粒体是产生ROS的主要场所,线粒体功能障碍导致ROS产生增加,过量产生的ROS破坏核及线粒体DNA、蛋白质、脂质等,导致线粒体受损,又进一步促进ROS的生成,形成的恶性循环导致细胞凋亡[9]。衰老细胞中,过量生成的ROS也参与调控自噬相关蛋白,导致心脏的选择性自噬功能不足,促进心脏衰老[10]。此外,在衰老的心脏,残余健康的心肌细胞会发生代偿性肥大,以维持相对正常的心脏功能[10]。目前,无论在植物还是在动物模型中都有报道精脒与精胺具有广谱的抗氧化功能[11]。我们前期研究也显示,外源性多胺能通过增加肝脏与心脏的抗氧化能力,延缓自然衰老大鼠的老化[12]。近年来学者们在多种属、多器官的研究证实,精脒通过诱导自噬,发挥抗衰老及衰老相关疾病的作用。Eisenberg等[13]证实,精脒通过诱导自噬,延长酵母、果蝇和蠕虫的寿命;喂食精脒的小鼠,寿命明显延长,精脒通过促进小鼠心肌细胞自噬及线粒体自噬,增强了衰老心肌线粒体呼吸功能,进而抑制老年小鼠心脏舒张功能的下降。我们前期应用蛋白质组学的研究发现,精胺、精脒通过影响老年大鼠心肌参与免疫反应、脂质代谢、凝血反应和谷胱甘肽代谢通路上的蛋白质分子,抑制老年大鼠心脏的病理改变[14]。因此,我们有理由认为本研究中外源性精脒可能是通过抗氧化、诱导自噬,发挥抗大鼠心脏老化作用。
总之,本研究证明,随着年龄增加,大鼠心肌多胺合成代谢下调、分解代谢上调、细胞自噬能力下降;多胺分解代谢与细胞自噬呈明显的负相关;外源性多胺可通过抗氧化、诱导自噬,延缓大鼠心脏老化。本研究为开发新的抗心脏衰老药物,进而降低衰老相关心血管疾病发生率及死亡率提供了理论基础。
(致谢: 本实验在哈尔滨医科大学基础医学院病理生理学教研室实验室完成,对本实验做出贡献的所有人员表示衷心的感谢!)
猜你喜欢
世界科学技术-中医药现代化(2022年2期)2022-05-25 13:16:04
世界科学技术-中医药现代化(2021年7期)2021-11-04 08:10:24
科学生活(2019年7期)2020-01-01 08:28:02
国际呼吸杂志(2019年8期)2019-04-29 09:15:10
养殖与饲料(2019年10期)2019-02-25 14:52:37
山东畜牧兽医(2018年3期)2018-04-26 09:10:34
中成药(2017年9期)2017-12-19 13:34:56
海南医学(2016年8期)2016-06-08 05:43:00
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:04
中国病理生理杂志(2015年8期)2015-12-21 12:38:08