
HIF-1α-Snail轴介导不同氧环境下结肠癌EMT-MET转化
2019-07-16 09:46:22雷星,陈熹,张迪,单涛
安徽医科大学学报 2019年6期
雷 星,陈 熹,张 迪,单 涛
结肠癌是常见消化系统肿瘤之一,发病率列世界第三,病死率列世界第四[1]。全球范围内结肠癌的发病率和病死率呈现上升的势头[2]。目前,早期治疗结肠癌是手术为主、化疗为辅的综合疗法,但其易复发和易转移的特点常常导致治疗失败[3],结直肠癌仍严重影响我国人民生命健康[3]。因此,寻找治疗干预的新靶点具有重要意义。
上皮-间质转化(epithelial-mesenchymal transition, EMT)指上皮到间质细胞的转化,它赋予细胞转移和入侵的能力,包括干细胞特征、减少凋亡与衰老和促进免疫抑制,不仅在发育过程中起着关键的作用,而且还参与组织愈合、器官纤维化和癌症发生等过程[4]。上皮-间质转化又是一个动态的可逆过程[5],Thiery et al[6]总结了肿瘤两步转移理论,认为原发上皮肿瘤依靠EMT程序来进行侵润和全身播散,播散的瘤细胞到达远处组织时,依靠逆转的过程间质表皮化(mesenchymal-epithelial transition, MET)最终定植形成转移灶。既往研究证实在结肠肿瘤中原发部位(低氧)及归巢部位(常氧或适度高氧)氧浓度差异参与EMT-MET两步转移过程,而目前EMT-MET过程中时空调控关键环节的确切机制尚未阐明。研究证明,TGF-β、Wnt/β-catenin、Notch、Hedgehog、IL-6/STAT3以及HIF-1α等信号通路的激活会调控下游Snail1、Snail2、Twist1、Twist2、ZEB1和ZEB2等转录因子,开启EMT进程[7]。而原发部位(低氧)及归巢部位(常氧或适度高氧)氧浓度差异是否可通过HIF-1α-Snail通路,是否可用来解释肿瘤两步转移复原机制还未见完全明确。该研究在体外模拟原发灶低氧环境,归巢部位常氧或适度高氧环境,运用激光共聚焦、qRT-PCR、Westernblot等方法,观察HIF-1α-Snail通路是否介导EMT、MET形态变化,探索结肠癌发生转移机制,为治疗研究提供新依据。
1 材料与方法
1.1 主要试剂二甲基亚砜(DMSO,美国Sigma公司);RIPA裂解液试剂盒(上海碧云天公司);DMEM培养液、胎牛血清(美国HyClone公司);抗E-cadherin抗体(sc92614)、vimentin抗体(sc80975)、HIF-1α抗体(cat. no. sc13515)、Snail抗体(cat. no. sc393172)和β-actin抗体(cat. no. sc517582)购自美国Santa Cruz 生物公司。
1.2 细胞培养和处理COLO 205(人结肠癌细胞系)购自美国ATCC公司。胎牛血清、100 U/ml青霉素、100 μg/ml链霉素的DMEM培养基用于培养细胞。细胞生长达到80%融合时,开始给与干预,分为:低氧组(氧浓度5%),常氧组(氧浓度21%),适度高氧组(氧浓度30%)。其中,常氧组直接置于普通CO2培养箱中培养,低氧及高氧组则置于自制培养器皿中(10 cm×15 cm×25 cm),持续通入含有5%、30%O2和5%CO2的混合气体。培养 48 h后更换培养基为3 ml无血清基础培养基,置于普通CO2培养箱中培养24 h后收集细胞,进行下一步试验。
1.3 细胞免疫荧光细胞培养,常规铺片,滴加PBS(0.01 mol/L,pH 7.4)于已知抗原标本片,润洗10 min,使标本片具有一定湿度。滴加PBS(0.01 mol/L,pH 7.4)适当稀释抗体后,覆盖已知抗原标本片。玻片置于有盖搪瓷盒内(37 ℃、30 min)。随后取出置于玻片架上,先用PBS(0.01 mol/L,pH 7.4)冲洗1~2次,然后按顺序PBS(0.01 mol/L,pH 7.4)三缸浸泡,每缸5 min,不时振荡。取出玻片,用滤纸吸去多余水分,滴加1滴荧光标记的抗人球蛋白抗体。将玻片平放在盖搪瓷盒内(37 ℃、30 min)。取出覆以盖玻片。荧光显微镜下观察。
1.4 RT-PCR细胞系COLO 205总RNA应用TRIzol试剂进行提取。使用2 μg RNA合成第一条cDNA链(RevertAid)。PCR引物设计:E-cadherin上游引物:5′-CAATGGTGTCCATGTGAACA-3′,下游引物:5′-CCTCCTACCCTCCTGTTCG-3′;vimentin上游引物: 5′-CGCTTCGCCAACTACAT-3′,下游引物:5′-AGGGCATCCACTTCACAG-3′;Snail上游引物:5′-CGCGCTCTTTCCTCGTCAG-3′,下游引物:5′-TCCCAGATGAGCATTGGCAG-3′;β-actin上游引物:5′-ATCGTGCGTGACATTAAGGAGAAG-3′,下游引物:5′-AGGAAGGAAGGCTGGAAGAGTG-3′。PCR反应条件:变性(94 ℃、3 min),再进行35循环(94 ℃、30 s,55 ℃、30 s,72 ℃、35 s),退火(72 ℃、5 min)。内参使用β-actin。PCR产物置于1.5%琼脂糖凝胶电泳后于紫外光下观察。
1.5 Western blot检测蛋白取5×105个对数生长期细胞加入预冷细胞裂解缓冲液(0.5 ml)冰上孵育(30 min)。离心取上清液,测定蛋白含量(BCA法),凝胶电泳分离蛋白(10% SDS-PAGE),目的蛋白半干转到硝化纤维膜上。后用含5%脱脂牛奶TBST进行封闭1 h,一抗孵育过夜(4 ℃),辣根过氧化物酶(HRP,1 ∶1 500)结合的二抗(Santa Cruz公司)室温孵育2 h,用增强型化学发光试剂盒(Amersham Pharmacia Biotech, Piscataway,NJ)显色,照相并分析结果。
1.6 统计学处理使用SPSS 16.0 软件进行统计学分析,采用One-way ANOVA 方差分析法,首先利用Levene方法进行方差齐性检验,确定方差齐性且整体比较组间差异有统计学意义后进一步作多重比较,多重比较采用LSD法。以P<0.05为差异有统计学意义。
2 结果
2.1 不同氧浓度对结肠癌Snail mRNA及蛋白表达的影响为了明确低氧是否影响癌细胞Snail表达,Western blot的方法检测了Snail在mRNA及蛋白水平的表达情况。RT-PCR结果显示低氧组Snail mRNA水平较常氧/高氧组明显升高(F=55.21,P<0.05)。Western blot结果显示低氧组Snail的蛋白表达水平则较常氧/高氧组显著上调(F=129.75,P<0.05)(图1)。以上结果进一步证实了低氧上调Snail表达可能是诱导EMT发生的关键转录点。相反,归巢环境下常氧或高氧状态下向MET转变。
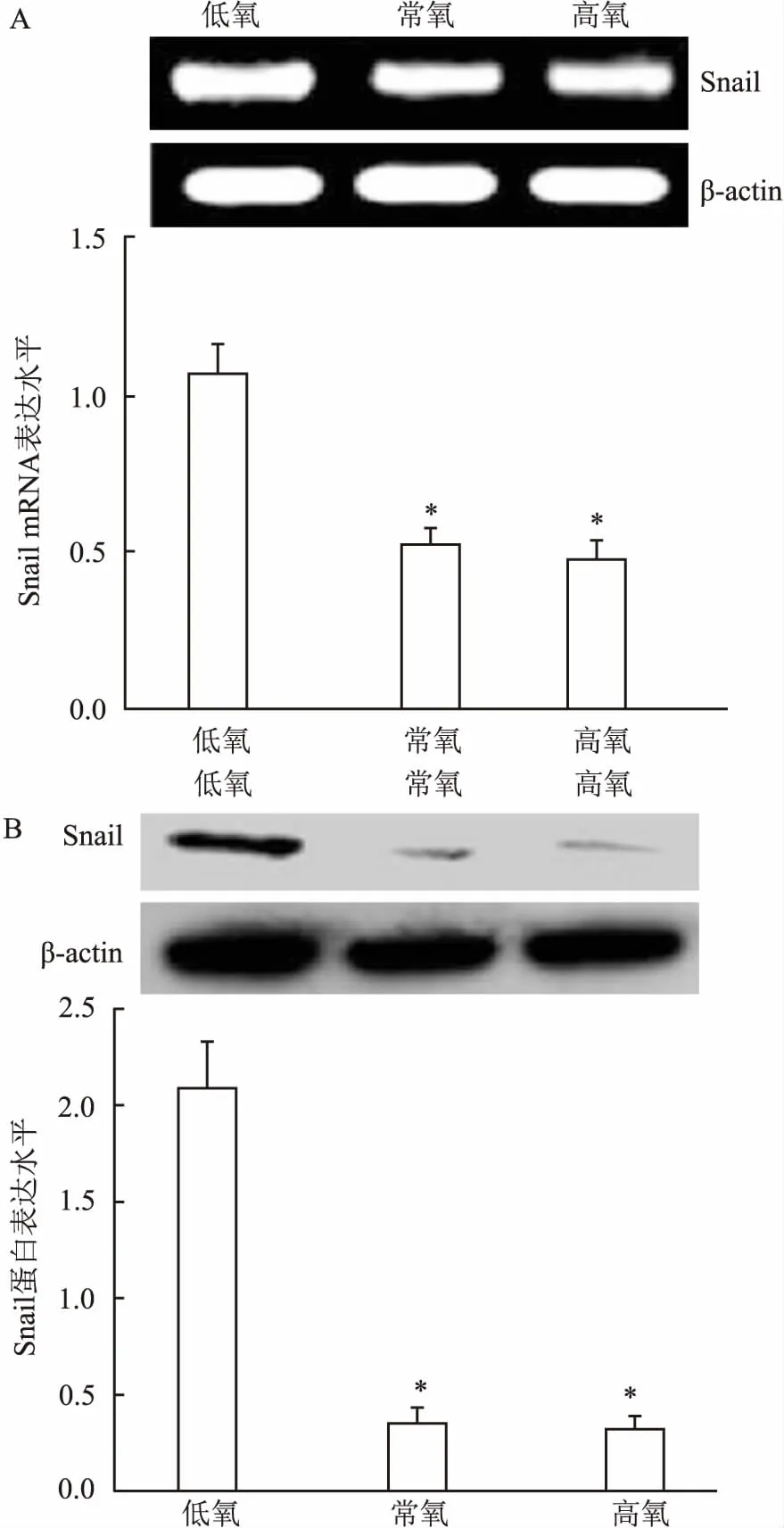
图1 RT-PCR及Western blot结果
A:RT-PC检测Snail mRNA水平;B. Western blot检测Snail蛋白水平;与低氧组比较:*P<0.05
2.2 低氧通过HIF-1α通路调控结肠癌Snail mRNA及蛋白的表达Western blot检测其表达情况,结果显示低氧组HIF-1α的蛋白表达水平较常氧/高氧组显著上调(F=126.59,P<0.05)(图2B)。以上结果进一步证实了低氧下结肠癌细胞中HIF-1α通路处于活化状态。为了验证低氧下结肠癌细胞中HIF-1α通路与结肠癌细胞EMT有关,HIF-1α特异性阻断剂预处理细胞24 h后再用不同氧浓度处理,检测Snail蛋白的表达。结果显示,加入阻断剂组Snail蛋白的表达与常氧组(P=0.23)/高氧组无差别(F=0.60,P=0.58)。相应的E-cadherin、Vimentin表达较常氧/高氧组无差别。以上结果证实了结肠癌细胞中HIF-1α-Snail调控轴是低氧诱导EMT发生关键机制,相应地常氧/高氧环境下HIF-1α-Snail调控轴失活可诱导MET转变。
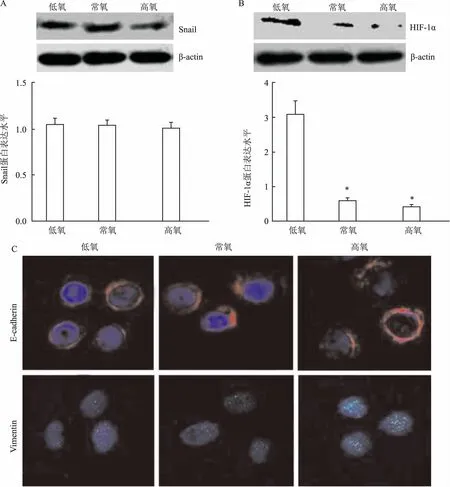
图2 HIF-1α通路调控结肠癌Snail mRNA及蛋白的表达
A: Western blot结果显示HIF-1α的蛋白表达水平;B: 阻断HIF-1α,Snail蛋白的表达情况比较(组间比较:P>0.05);C:阻断HIF-1α,E-cadherin/Vimentin各组表达较常氧/高氧组无差别;与低氧组比较:*P<0.05
3 讨论
EMT给了癌细胞"双腿"让它们能够脱离原位肿瘤,并通过促进血管生成和内渗帮助肿瘤细胞进入血液循环,形成循环肿瘤细胞(circulating tumor cells,CTCs)[5]。肿瘤细胞发生EMT,除了可以促进CTCs产生,还可以促进CTCs存活,CTCs定植通过MET形成肿瘤转移灶[5]。但目前EMT-MET这一时空调控关键环节的确切机制尚未阐明。此研究证实肿瘤原发处低氧微环境可通过HIF-1α-Snail调控轴调控细胞EMT发生,而常氧/高氧环境下则发生相反的现象。此结果证实假说,即已习惯乏营养环境下生存的肿瘤细胞进入归巢富氧微环境后,失去低氧诱导,迫使肿瘤细胞发生EMT-MET转换,形成新的转移灶,一旦肿瘤在空间上生长到一定体积后,又建立新的乏氧微环境,再次形成新的恶性循环,以此方式不停的促进肿瘤进展。
缺氧是实体肿瘤在发生、发展的过程中,局部微环境最显著特征[8]。胃癌中已经证实存在明显的缺氧状态,胃癌的这种缺氧环境与其高侵袭特征密切相关[9]。因此低氧的肿瘤组织内癌细胞的生物学行为更趋向恶性,形成更具侵袭性的类型,并表现出对治疗的抵抗性。低氧浓度的确可以影响间质-上皮可塑性,介导EMT发生[9]。Huang et al[10]研究表明缺氧依靠HIF-1α正向调节Slug的表达,通过EMT程序获得高侵袭转移能力。本研究结果和上述结果一致,也再次证实了此结论。并在此基础上本试验继续在富氧环境下观察到MET现象。提示氧浓度的差异在肿瘤转移的不同阶段,均对EMT和MET之间转换过程产生重要影响。这种原发灶低氧环境和归巢部位富氧环境的差异有可能是EMT-MET转换的新视角,但更深入的机制研究仍需进一步验证。
EMT赋予肿瘤细胞一些新特性,主要有增加细胞迁移侵袭,使其突破基膜,促进肿瘤细胞进入血液循环,形成CTCs。Tsuji et al[11]和Thiery et al[6]证实只有发生EMT的细胞侵入相邻组织和血管,表明发生EMT是产生CTCs的重要前提。转移到远部位CTCs适应周围基质环境后,还会再经历一个MET转换,重新获得增殖特性,进而形成转移灶[12]。研究这种可逆并且瞬时的EMT-MET转换将是一个重要方向,无疑有大量未知问题亟待解决。深入研究CTCs-EMT产生和转化的机制及其生物学意义必将有助于揭开肿瘤发生、发展和转移之谜,并且可为制定新的肿瘤靶向治疗策略提供依据。
猜你喜欢
食品安全导刊(2022年2期)2022-03-18 00:35:00
食品工业(2021年12期)2021-12-31 02:57:08
天津医科大学学报(2021年4期)2021-08-21 02:14:50
天津医科大学学报(2021年3期)2021-07-21 09:03:42
中国医药导报(2018年36期)2018-03-04 06:50:42
西南国防医药(2016年7期)2016-12-01 06:01:15
国外医药(抗生素分册)(2016年5期)2016-07-12 14:25:34
中国卫生标准管理(2015年1期)2016-01-14 03:41:26
实验动物与比较医学(2014年5期)2014-02-28 14:53:12
河南医学研究(2014年3期)2014-02-27 14:51:48
