
重组人促红细胞生成素对肝细胞模拟缺血/再灌注损伤的保护作用
2019-06-27 06:44:10李龙温陈解维敏刘文玉刘波陶开山窦科峰慕喜喜神木市医院肝胆外科陕西神木79300空军军医大学西京医院肝胆外科陕西西安7003西安市中心医院普通外科陕西西安70003
实用器官移植电子杂志 2019年3期
李龙,温陈,解维敏,刘文玉,刘波,陶开山,窦科峰,慕喜喜,3(.神木市医院肝胆外科,陕西 神木 79300;.空军军医大学西京医院肝胆外科,陕西西安 7003;3.西安市中心医院普通外科,陕西 西安 70003)
由于移植外科技术的成熟以及新型免疫抑制药物的开发,越来越多的终末期肝功能衰竭患者选择进行肝移植来挽救生命,由此造成严重的供肝短缺,并引起边缘供肝的使用逐渐增加,但是此类供体肝脏对缺血/再灌注损伤(ischemia reperfusion injury,IRI)十分敏感,常常预后不良[1-2]。尽管有学者已开始尝试无血流阻断肝移植,并取得初步成果[3-4],但IRI仍是肝移植手术中无法避免的重要环节,严重影响移植肝的预后[5]。在移植肝IRI过程中伴随着活性氧自由基的释放,由此引发氧化损伤[6],常常导致细胞凋亡、坏死,包括引发细胞过度自噬死亡[7]。自噬(autophagy)是细胞利用胞内溶酶体降解受损、变性、衰老的蛋白质,实现物质和能量的回收利用,从而维持细胞稳态,但自噬活性过高会诱发细胞Ⅱ型程序性死亡[8]。促红细胞生成素(erythropoietin,EPO)主要由肾脏分泌促进红细胞生成来应对缺氧。有研究表明,EPO还具有抗炎、抗氧化、抗凋亡等作用,有学者认为其可能是一种细胞保护因子[9]。重组人促红细胞生成素(recombinant human erythropoietin,rhEPO)是通过DNA重组生产的EPO,其序列和生物活性均与内源性EPO相同。本研究拟采用H2O2建立肝细胞缺氧/复氧培养模型,探索rhEPO处理对肝细胞模拟IRI的保护作用,以及与肝细胞自噬活性的关系,揭示其可能涉及的分子机制。
1 材料与方法
1.1 材料:大鼠肝细胞株BRL3A购于上海中科院细胞库。rhEPO购自克隆生物高技术有限公司。30% H2O2试剂购自国药集团化学试剂有限公司。吖啶橙染色试剂盒购自Sigma。CCK-8检测试剂盒购自碧云天生物技术研究所。Western blot检测抗体:anti-LC3B、anti-p62、anti-p-p85(1∶1 000)、anti-p85(1∶ 500)购自Abcam,anti-p-AKT(1∶500)、anti-AKT (1∶1 000)购自CST,anti-βactin (1∶300)购自Abmart,HRP标记的二抗购自碧云天生物技术有限公司。PrimeScript RT Master Mix试剂盒购自Takara。BCA蛋白检测试剂盒购自碧云天生物技术有限公司。天冬氨酸转氨酶 (aspartate aminotransferase,AST)和丙氨酸转氨酶(alanine aminotransferase,ALT)测定试剂盒购自Sigma。
1.2 细胞培养与分组:取指数生长期的BRL3A细胞,按照2000个 (100 μl·孔) 的密度种植于96孔板,加入H2O2(终浓度为200 μmol/L)刺激细胞2 h,取3代培养的BRL3A细胞随机分为4组:① 正常对照组;② H2O2处理组;③ H2O2+10 U/ml rhEPO处理组;④ H2O2+ 20 U/ml rhEPO处理组,每组分为6个复孔。
1.3 细胞存活率的检测:将各组细胞继续培养4 h后,采用CCK-8测定细胞存活率,每孔(200 μl体积溶液)加入10 μl的CCK-8溶液,然后用加入了相应体积的细胞培养液以及CCK-8溶液,不含细胞的孔作空白对照。放入培养箱内继续孵育约1 h左右,用酶标仪检测,在450 nm测定吸光度值。各孔OD值为实际OD值减去空白对照组OD值,每组复孔去掉最大值和最小值 (n=6孔),取均数。重复实验(n=3次),根据结果绘制细胞存活曲线。
1.4 转氨酶释放测定:取各组培养细胞的上清液,按照试剂盒说明书的方法,化学比色法测定培养液中AST、ALT的含量。
1.5 吖啶橙染色检测肝细胞自噬小体形成:24孔板培养孔中加入细胞爬坡玻璃片(预先消毒处理),按上述实验分组,每组3个复孔。将各组细胞继续rhEPO处理4 h后,去除培养基,无菌PBS洗3遍,然后向各孔中加入吖啶橙荧光染色液,保持吖啶橙染液终浓度20 μg/ml,进行室温孵育约15 min。移除染色液后,用无菌的PBS清洗约4遍,置于倒置荧光显微镜下,观察染色结果。
1.6 Western Blot检测:样品中加入蛋白酶抑制剂后,在冰上裂解细胞,提取蛋白99℃变性10 min。BCA蛋白检测试剂盒检测蛋白浓度。制作10%的SDS-PAGE分离胶,蛋白上样后电泳,110 V,120 min。转膜,220 mA,40 min。5%牛奶封闭后,加入一抗4℃孵育过夜。用辣根过氧化物酶标记的二抗,室温孵育1 h。最后用ChemiDocTM XRS +和Image Lab TM software显影,并计算条带灰度值。
2 结 果
2.1 rhEPO可以逆转H2O2导致的BRL3A细胞存活率下降(表1、图1):H2O2处理后的BRL3A细胞存活率明显下降(95.36±3.12%比57.82±2.54%,P<0.05),经过不同浓度的rhEPO处理后,BRL3A细胞存活率有不同程度升高,差异有统计学意义(P<0.05)。
表1 BRL3A细胞存活率(±s)

表1 BRL3A细胞存活率(±s)
注:与对照组比较,aP<0.05;与200 μmol/L H2O2处理组比较,bP<0.05
组别 例数(例) 存活率(%)正常对照组 3 95.36±3.12 H2O2处理组 3 57.82±2.54a H2O2 +10 U/ml rhEPO处理组 3 72.55±2.79b H2O2 +20 U/ml rhEPO处理组 3 81.63±3.45b

图1 CCK-8测定各组BRL3A细胞存活率
2.2 各组BRL3A细胞上清中转氨酶的含量(表2、图2):H2O2处理后的BRL3A细胞上清中AST和ALT的释放量明显增加〔AST(U/L):574±21.34比75±11.63;ALT(U/L):378±16.42 比 66±10.21,P<0.05〕;经过不同浓度的rhEPO预处理,AST和ALT的释放量明显下降,与200 μmol/L H2O2处理组比较,差异有统计学意义(P<0.05)。
表2 各组细胞培养上清中转氨酶比较(IU/L,±s)
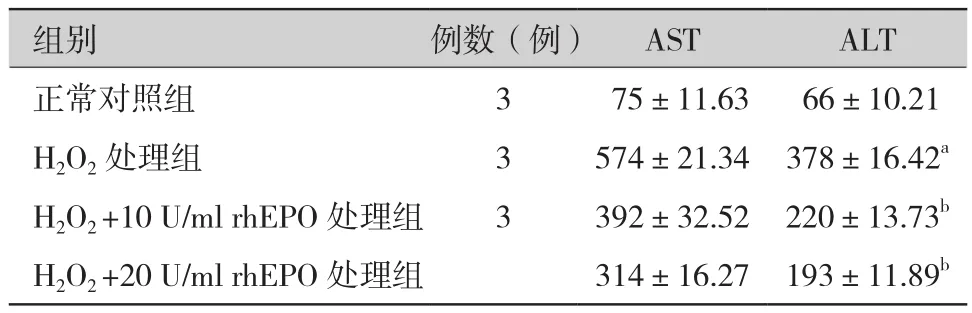
表2 各组细胞培养上清中转氨酶比较(IU/L,±s)
注:与对照组比较,aP<0.05;与200 μmol/L H2O2 处理组比较,bP<0.05
组别 例数(例) AST ALT正常对照组 3 75±11.63 66±10.21 H2O2处理组 3 574±21.34 378±16.42a H2O2 +10 U/ml rhEPO处理组 3 392±32.52 220±13.73b H2O2 +20 U/ml rhEPO处理组 314±16.27 193±11.89b

图2 化学比色法检测培养上清中AST和ALT含量
2.3 rhEPO能明显抑制H2O2导致的BRL3A细胞致死性自噬活性,使胞内自噬小体明显减少(图3):为测定各组BRL3A细胞的自噬活性,本研究中采用吖啶橙染色。正常对照组BRL3A细胞自噬活性水平较低,经H2O2处理后,BRL3A细胞自噬活性明显升高,胞内酸性小体增加,发生自噬性死亡。给予不同浓度的rhEPO处理后,BRL3A细胞自噬活性得到明显抑制,其中20 U/ml rhEPO处理组的胞内酸性小体的数量明显减少,和H2O2组比较,差异具有统计学意义(P<0.05)。
2.4 rhEPO处理能明显抑制LC3-Ⅱ蛋白表达,同时p62蛋白增多(图4):LC3B蛋白分子两个亚型,两者的比例可以反映细胞自噬的活性强弱,当自噬增强时,形成酸性自噬小体,使LC3-Ⅰ向LC3-Ⅱ转变,同时,随着细胞自噬活性增强,自噬相关蛋白p62会成比例的降解。与对照组相比,H2O2处理可引起BRL3A细胞自噬活性增加,LC3-Ⅰ向LC3-Ⅱ转变,同时,自噬相关蛋白p62被大量降解,差异具有统计学意义(P<0.05)。给予rhEPO处理后,特别是在20 U/ml rhEPO浓度下,BRL3A细胞自噬活性被明显抑制,与H2O2处理组比较,LC3-Ⅱ/LC3-Ⅰ的比例显著下降,同时伴随有p62蛋白的重新堆积,提示BRL3A细胞致死性的自噬行为被逆转。

图3 吖啶橙染色检测BRL3A细胞胞内自噬小体的形成

图4 Western-Blot检测各组培养BRL3A细胞中自噬相关蛋白LC3-Ⅱ和p62的表达变化
2.5 rhEPO处理能显著增加p-p85和p-AKT表达,活化PI3K/AKT信号通路(图5):检测p85和AKT的磷酸化蛋白p-p-p85和p-AKT的表达量,观察信号通路PI3K/AKT的活化水平。与正常对照组相比较,H2O2处理可引起BRL3A细胞p-p85和p-AKT的表达量下降。给予rhEPO处理后,磷酸化蛋白p-p85和p-AKT的表达又出现增多,这和BRL3A的自噬活性变化水平相一致。
3 讨 论
多种肝脏外科操作中,如肝切除、肝移植等,肝脏组织都将不可避免的面临组织IRI[10]。然而,迄今为止,其具体的损伤机制尚未完全阐明,在临床上也始终缺乏有效的干预手段。研究证实,无氧代谢与酸中毒、钙离子超载、氧自由基的生成、细胞凋亡、内皮素、肝脏微循环衰竭、补体系统、热休克蛋白等均参与其中[11]。

图5 Western Blot检测各组培养BRL3A细胞中PI3K/AKT信号通路蛋白p-p85和p-AKT的表达变化
缺氧/复氧模型是体外从细胞水平研究IRI的理想模型[12],且有研究证实利用H2O2可成功建立细胞缺氧/复氧损伤模型[13]。本研究通过建立肝细胞模拟缺血/再灌注损伤的体外培养模型,利用H2O2释放的活性氧自由基,造成大鼠BRL3A肝细胞的氧化应激损伤,然后给予rhEPO观察其对细胞损伤的防御或修复效应,结果显示,其能有效提升氧化应激环境下肝细胞的生存能力。既往研究发现EPO不仅能够促进红细胞生成,提高机体的携氧能力,改善组织器官缺氧,同时还具备许多特殊的生理功能,如抗炎[14-15]、抗凋亡[16]、促进血管生成[17]等。我们的结果说明,EPO具有显著的抗氧化损伤功能,且这种保护作用与rhEPO浓度呈正相关。
值得注意的是,近年来,许多学者还发现肝脏IRI过程中还存在着特殊的细胞生理活动,即自噬现象[18]。生理状况下,肝脏组织细胞保持着低水平的自噬活性,通过物质和能量循环来维持细胞的稳态[19]。但在一些应激条件下,这种自噬活性会被放大,打破这种稳态平衡,形成细胞Ⅱ型程序性死亡,称之为自噬性死亡[20-21]。本研究中在给予H2O2刺激后,吖啶橙染色可观察到肝细胞胞质中出现大量的酸性自噬小体,同时,可检测到微管相关蛋白1轻链3(microtubuleassociated protein 1 light chain 3,LC3/Atg8)亚型LC3-Ⅰ向LC3-Ⅱ的转变,且p62大量降解[22],提示H2O2加强了肝细胞的自噬活性,结合细胞存活率下降,我们推测这种过度强化的自噬现象即为致死性细胞自噬。自噬涉及多种信号调节通路,如PI3K/AKT/mTOR[23]、MAPK/ERK1/2[24]、p53/Genotoxic Stress[25]、AMPK[26]等,其中最为经典的即为PI3K/AKT/mTOR信号通路,PI3K/AKT的磷酸化可促进mTOR的活化,对细胞自噬产生明显的抑制作用,反之,即可启动细胞自噬[27-28]。研究证实,mTOR作为该通路的关键效应分子,激活的mTOR,即p-mTOR可以抑制以及破坏自噬体的形成,反馈调节细胞的自噬水平[28-29]。本研究中 H2O2刺激细胞后,p-p85和p-AKT的表达明显下降,说明PI3K/AKT的磷酸化程度变低,提示PI3K/AKT信号被抑制,细胞自噬过程被加强,即形成过度激活现象,导致肝细胞死亡。给予rhEPO处理后,p-p85和p-AKT的表达回升,PI3K/AKT的磷酸化导致mTOR被激活,自噬效应得到明显的抑制。本研究的不足之处在于没有进一步做mTOR的蛋白水平检测,以进一步验证我们的猜想,后续需要进一步检测磷酸化mTOR,完善相关的分子机制。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:52:18
建材发展导向(2021年11期)2021-07-28 06:57:22
当代水产(2020年10期)2020-03-17 07:02:48
当代水产(2019年8期)2019-10-12 08:57:26
癌变·畸变·突变(2016年3期)2016-02-27 06:15:36
法医学杂志(2015年4期)2016-01-06 12:36:36
法医学杂志(2015年4期)2016-01-06 12:36:36
哈尔滨医药(2015年4期)2015-12-01 03:57:54
中国海洋大学学报(自然科学版)(2014年8期)2014-02-28 12:21:24
河南医学研究(2014年5期)2014-02-27 14:52:51
