
增水和施肥对苜蓿-冬小麦轮作系统冬小麦叶片光合速率和叶绿素含量的影响
2019-06-24 08:43:22张晓琳陈梦恬翟鹏辉赵祥董宽虎
山西农业大学学报(自然科学版) 2019年4期
张晓琳,陈梦恬,翟鹏辉,赵祥,董宽虎*
(1.山西农业大学 动物科技学院,山西 太谷 030801;2.兰州大学 生命科学学院草地农业生态系统国家重点实验室,甘肃 兰州 730000)
黄土高原是我国高原的重要组成部分,是全球占地面积最广的黄土区域,该地区矿物质养分含量丰富,利于农作物的生长。由于水蚀和风蚀等外界作用,黄土高原面临着严重的水土流失和土壤肥力下降等问题[1]。苜蓿属于豆科牧草,营养丰富,具有生物固氮作用能提高土壤肥力,且具有庞大的根系能保持土壤水分,是温带地区重要的牧草资源。将苜蓿引入小麦的作物体系形成草田轮作系统,在生产实践过程中表现出强大的优势[2]。苜蓿-小麦轮作系统是黄土高原地区重要的草田轮作模式,然而水分和养分是限制该地区作物生长的重要因素,影响作物与环境的相互作用,最终影响该生态系统的碳循环过程。因此,研究增水和施肥对叶片光合速率和叶绿素含量影响,对探讨草田轮作系统的碳循环过程以及该系统的管理和可持续发展具有重要意义。
目前,国内外开展了一些关于冬小麦叶片光合作用速率和叶绿素含量的研究,大多数研究发现,增水和施氮磷肥显著增加冬小麦旗叶的叶绿素含量,从而促进冬小麦抽穗期到灌浆期的净光合速率,提高叶片光合能力[3~6],进而增加小麦产量。但也有结果发现,施肥在不灌水条件下抑制净光合速率,而在灌水条件下促进净光合速率,改善花后旗叶光合性能[7],因此增水和施肥对冬小麦光合速率的影响可能存在交互作用。但有关增水和施肥对冬小麦的研究,主要集中在特定物候期叶片光合作用速率和叶绿素含量的响应,对整个生长季的动态变化研究很少。
为研究苜蓿-小麦草田轮作系统冬小麦叶片光合作用速率和叶绿素含量是如何响应增水和施肥的,本研究利用山西农业大学已经建立3年的紫花苜蓿实验平台,种植冬小麦,并以此为研究对象,探讨以下问题:(1)增水处理下,紫花苜蓿-冬小麦草田轮作系统中冬小麦叶片光合作用速率和叶绿素含量是如何响应的?(2)施肥处理下,紫花苜蓿-冬小麦草田轮作系统中冬小麦叶片光合作用速率和叶绿素含量是如何响应的?(3)增水和施肥对紫花苜蓿-冬小麦草田轮作系统中冬小麦叶片光合作用速率和叶绿素含量是否存在交互作用?是相互促进,还是相互抵消?
1 材料与方法
1.1 样地概况
研究地点位于山西省晋中市太谷县山西农业大学动物科技学院试验站内(112°35′E,37°25′N),在地貌上属于黄土高原,海拔约800 m,多年平均降水量为458 mm,降水多集中在7~9月份。
1.2 试验设计
本试验于2017—2018年进行,试验地在2014年春季播种紫花苜蓿,撒播。紫花苜蓿翻耕前0~10 cm土壤中有机碳含量为23.96 g·kg-1,土壤磷含量3.29 g·kg-1,土壤pH值7.99。2017年进行紫花苜蓿与冬小麦轮作试验,2017年9月28号人工播种冬小麦,冬小麦品种为中麦175号,由山西农业大学农学院提供,播种量为187.5 kg·hm-2,行间距为15 cm,深度2~3 cm。冬小麦生长阶段人工去除田间杂草。
本试验采用完全随机区组设计,共4个处理,即对照(无增水施肥,CK)、增水 (W)、施肥 (F)、增水施肥 (W+F),每个处理4个重复,共16个样方。各样方面积为16 m2(4 m × 4 m),样方之间过道宽度不低于2 m。在施肥和增水施肥样方中施入底肥,施肥时间为播种前一周,施用量为尿素225 kg N·hm-2和磷酸二氢铵150 kg P2O5·hm-2。在增水和增水施肥样方中,按照冬小麦的需水量情况进行5次适量补水:(1)越冬水(2017.12.6):灌水量900 m3·hm-2,(2)返青起身水(2018.3.4):500 m3·hm-2,(3)拔节孕穗水(2018.4.17):900 m3·hm-2,(4)抽穗扬花水(2018.5.1):900 m3·hm-2,(5)灌浆麦黄水(2018.5.14):675 m3·hm-2。
1.3 光合生理特性和叶绿素含量的测定
在各试验小区里划定1 m2(1 m×1 m)样方,用于测定冬小麦叶片光合生理特性和叶绿素含量,并选择长势良好具有代表性的5株冬小麦进行标记,每次测定时选择植株上第一片完整的完全伸展的叶片。使用便携式光合作用测量系统 LI-COR 6400 (Li-Cor Inc., Lincoln, NE, USA)测定冬小麦叶片光合作用,测定时需连接红蓝光源探头,设定光照强度为2 000 μmol·m-2·s-1,得到叶片净光合速率,叶片气孔导度,叶片蒸腾速率以及叶面饱和蒸汽压差。利用便携式叶绿素仪 SPAD-502 Puls (Konica Minolta Sensing, INC.) 监测叶片叶绿素含量,选择与测定光合速率相同位置的叶片,每片叶测定5个点,计算其平均值即为该叶片的叶绿素含量,每个样方选取5株小麦。测定时间分别为(1)2017年:10月29-12月2日,即冬小麦出苗以后到进入越冬期初期;(2)2018年:3月14日-6月4日,即冬小麦从越冬期末期至成熟期。测定频次为每周1次。
1.4 统计方法
首先测定每片冬小麦叶片的叶绿素含量,并计算小区内5株冬小麦叶绿素含量的平均值,即为该样方的叶片叶绿素含量。本研究采用三因素重复测定方差分析法分析时间、增水、施肥及其交互作用对2017年和2018年冬小麦叶片净光合速率,气孔导度,蒸腾速率、叶面饱和蒸汽压差和叶绿素含量的影响。显著性水平为P<0.05。统计分析过程应用 SPSS 19.0 完成,所有图形的制作使用SigmaPlot 10.0完成。
2 结果与分析
2.1 增水和施肥对冬小麦叶片光合生理特性的影响
本研究结果显示,2017年,增水和施肥及其交互作用对冬小麦叶片净光合速率、气孔导度、蒸腾速率以及叶面饱和蒸气压差的影响作用不显著,但以上指标随时间变化显著。2018年,增水分别显著提高光合速率20.3%、气孔导度43.8%和蒸腾速率31.4%,但显著降低叶面饱和蒸气压差6.5%;施肥仅显著降低气孔导度的8.2%,对以上其它指标作用均不显著;增水施肥对以上各指标均不存在交互作用,但以上各指标仅随时间变化显著。(图1和图2,表1和表2)
表1三因素重复测定方差分析法分析测定时间(D)、增水(W)、施肥(F)及其交互作用对2017年冬小麦叶片光合生理特性和叶绿素含量的影响结果(P值)
Table1Theresults(Pvalues)ofrepeated-measuresANOVAsofwateraddition(W),fertilizeraddition(F)andtheirinteractionsonphotosyntheticphysiologicalcharacteristicsandleafchlorophyllcontent(SPADvalue)ofwinterwheatin2017
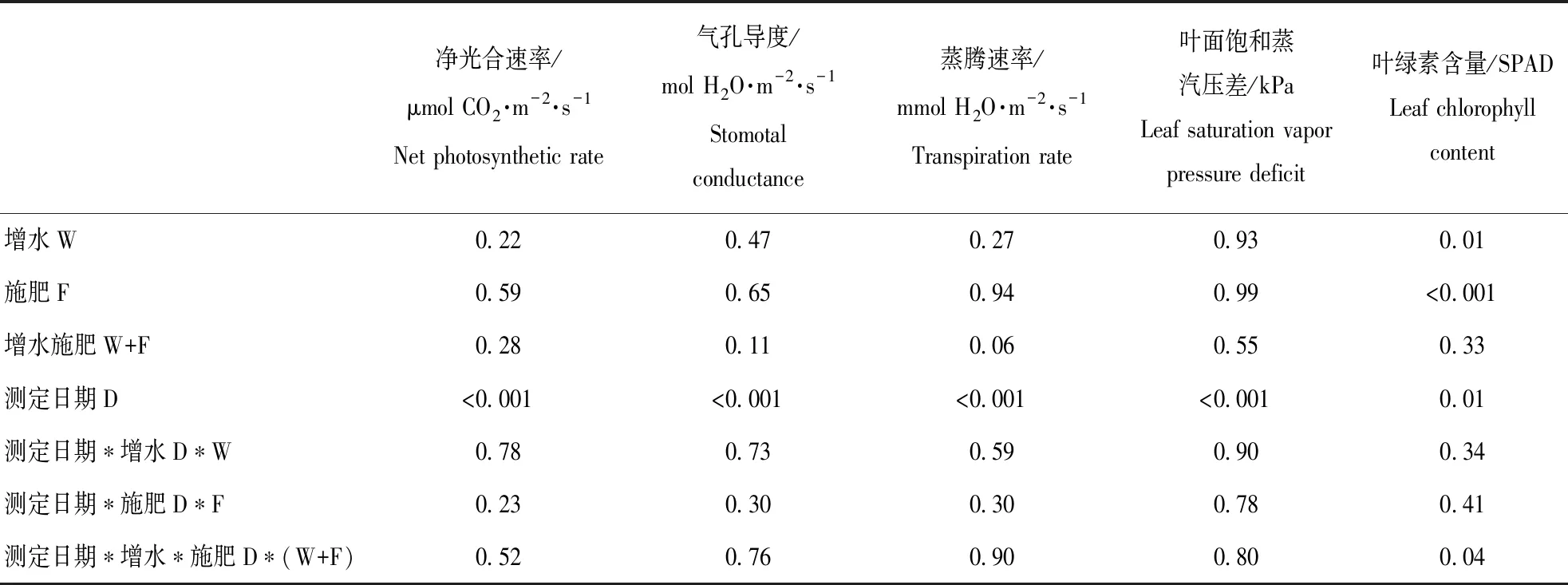
净光合速率/μmol CO2·m-2·s-1Net photosynthetic rate气孔导度/mol H2O·m-2·s-1Stomotal conductance蒸腾速率/mmol H2O·m-2·s-1Transpiration rate叶面饱和蒸汽压差/kPaLeaf saturation vapor pressure deficit叶绿素含量/SPADLeaf chlorophyll content增水W0.220.470.270.930.01施肥F0.590.650.940.99<0.001增水施肥W+F0.280.110.060.550.33测定日期D<0.001<0.001<0.001<0.0010.01测定日期∗增水D∗W0.780.730.590.900.34测定日期∗施肥D∗F0.230.300.300.780.41测定日期∗增水∗施肥D∗(W+F)0.520.760.900.800.04
2.2 增水和施肥对冬小麦叶片叶绿素含量的影响
三因素重复测定方差分析结果显示,2017年增水和施肥分别显著提高冬小麦叶片叶绿素含量的2.7%和8.2%,但增水和施肥对冬小麦叶片叶绿素含量的影响不存在交互作用(图3,表1)。随着时间的变化,冬小麦叶绿素含量变化显著,增水和施肥对冬小麦叶片叶绿素含量的影响不显著,但其交互作用对冬小麦叶片叶绿素含量的影响作用显著(表1)。2018年增水和施肥显著提高冬小麦叶绿素含的2.3%和2.0%,但两者对冬小麦叶片叶绿素含量的影响不存在交互作用(图3,表2)。随着时间的变化,冬小麦叶绿素含量变化和施肥下叶绿素含量变化显著,但增水、增水施肥对冬小麦叶绿素含量的作用均不显著(表2)。
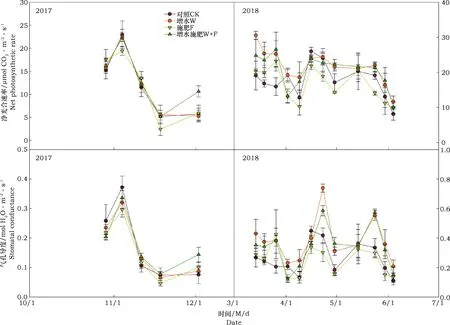
图1 2017年和2018年冬小麦叶片净光合速率和气孔导度动态Fig.1 Dynamics of net photosynthetic rate and stomotal conductance on winter wheat in 2017 and 2018

图2 2017年和2018冬小麦叶片蒸腾速率和叶面饱和蒸气压差动态Fig.2 Dynamics of transpiration rate and leaf saturation vapor pressure deficit on winter wheat in 2017 and 2018
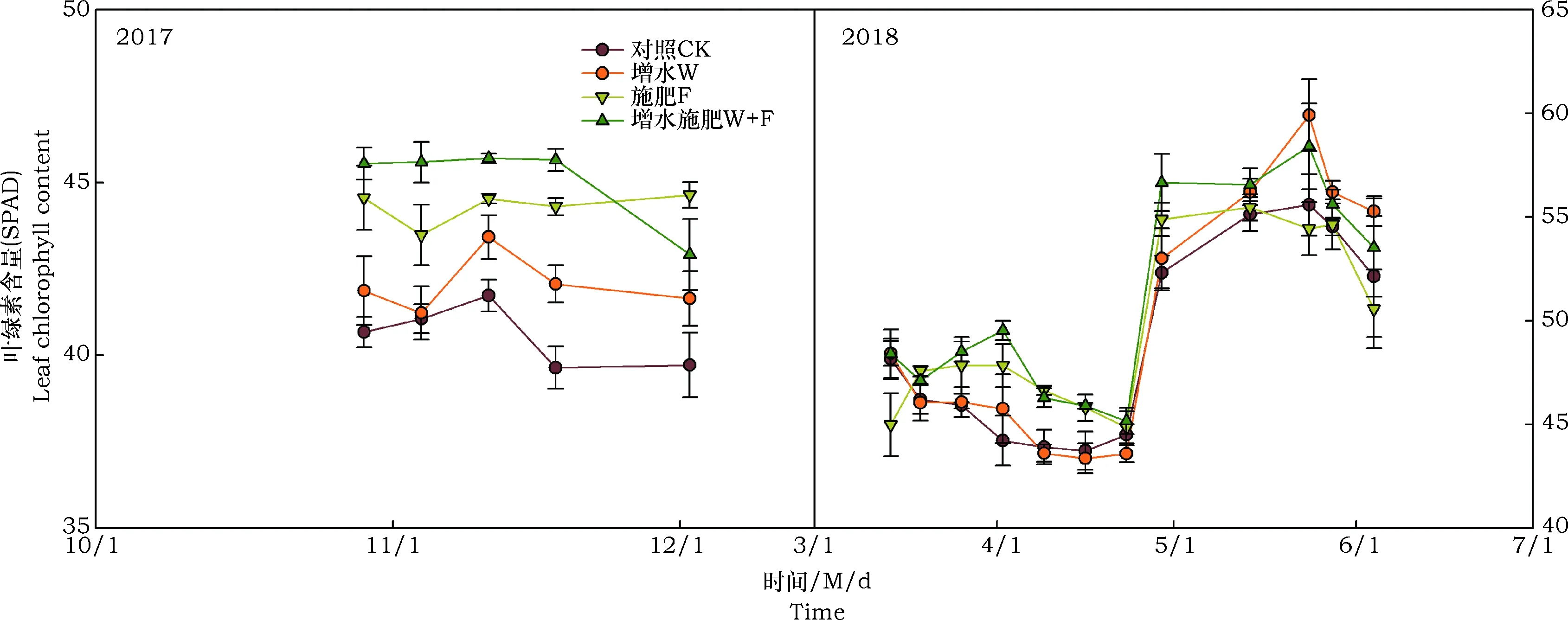
图3 2017年和2018年冬小麦叶片叶绿素含量动态Fig.3 Dynamics of leaf chlorophyll content of winter wheat in 2017 and 2018
表2三因素重复测定方差分析法分析测定时间(D)、增水(W)、施肥(F)及其交互作用对2018年冬小麦叶片光合生理特性和叶绿素含量的影响结果(P值)
Table2Theresults(FandPvalues)ofrepeated-measuresANOVAsofwateraddition(W),fertilizeraddition(F)andtheirinteractionsonphotosyntheticphysiologicalcharacteristicsandleafchlorophyllcontent(SPADvalue)ofwinterwheatin2018
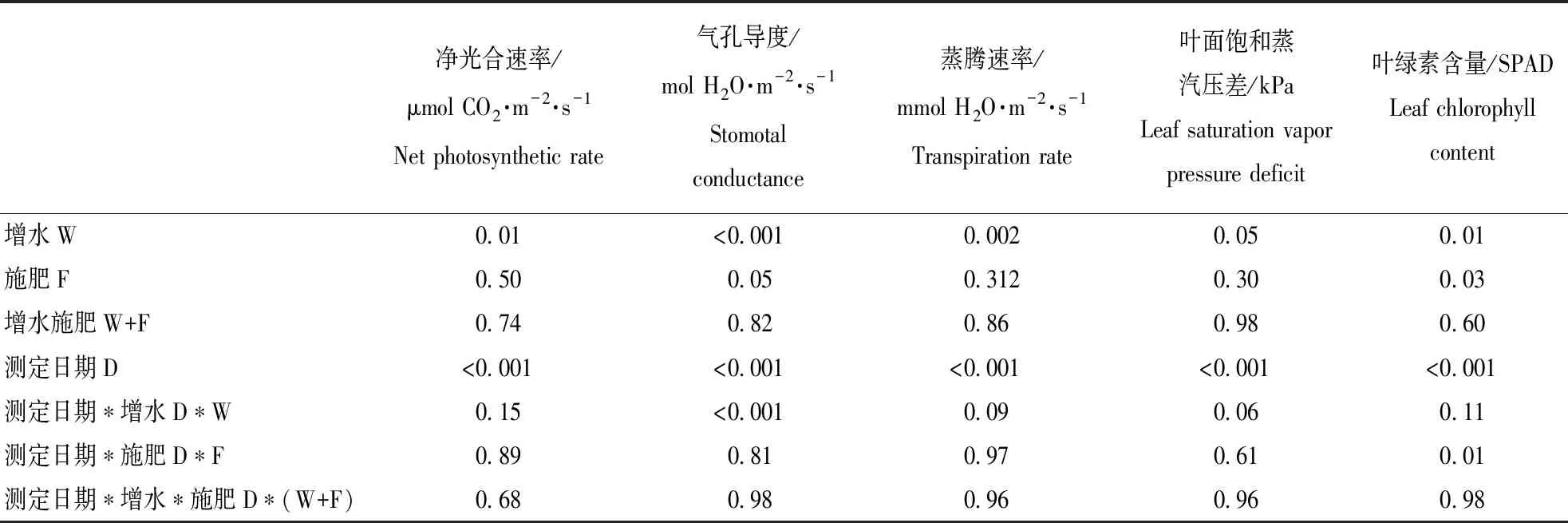
净光合速率/μmol CO2·m-2·s-1Net photosynthetic rate气孔导度/mol H2O·m-2·s-1Stomotal conductance蒸腾速率/mmol H2O·m-2·s-1Transpiration rate叶面饱和蒸汽压差/kPaLeaf saturation vapor pressure deficit叶绿素含量/SPADLeaf chlorophyll content增水W0.01<0.0010.0020.050.01施肥F0.500.050.3120.300.03增水施肥W+F0.740.820.860.980.60测定日期D<0.001<0.001<0.001<0.001<0.001测定日期∗增水D∗W0.15<0.0010.090.060.11测定日期∗施肥D∗F0.890.810.970.610.01测定日期∗增水∗施肥D∗(W+F)0.680.980.960.960.98
2.3 冬小麦叶片净光合速率与相关指标的关系
线性回归分析结果显示,随着冬小麦叶片气孔导度增加其净光合速率增加,2017年和2018年气孔导度对净光合速率变化的解释度分别为91%和78%(图4a)。2017年随着冬小麦叶片蒸腾速率增加净光合速率快速增加,但2018年增加速率相对缓慢,对净光合速率变化的解释度分别为89%和14%(图4b)。随着冬小麦叶面饱和蒸气压差增加净光合速率降低,2017年和2018年对净光合速率变化的解释度分别为76%和44%(图4c)。随着冬小麦叶片叶绿素含量增加净光合速率呈线性增加,2017年和2018年对净光合速率变化的解释度分别为20%和39%(图4d)。
3 讨论
3.1 增水对冬小麦叶片光合作用和叶绿素含量的影响
水分是影响陆地生态系统植物生长发育过程的重要因素,通过改变植物叶片光合生理特性和叶绿素含量,从而影响叶片水平碳循环过程,最终影响生态系统碳循环[8, 9]。有研究发现叶片气孔导度、蒸腾速率、叶面饱和蒸气压差是影响叶片光合速率的重要因素,气孔导度是影响进入叶片水汽平衡的关键因素,蒸腾速率影响植物对水分和养分的吸收,叶面饱和蒸气压差是叶片和大气之间的水蒸气压差值,是水蒸气从叶片向空气扩散的动力[10]。另外,增水提高植物对养分包括氮元素和磷元素的利用效率,促进叶绿素合成,从而提高了叶绿素含量,影响叶片光合速率[11]。本研究发现增水促进2018年叶片光合速率,主要是由于一方面增水提高叶片的气孔导度提高CO2的摄入量,另一方面增水提高蒸腾速率和降低叶面饱和蒸气压差促进植物对水分和养分的吸收。但增水对叶片叶绿素含量的改变比例在2017年高于2018年,说明可能随着时间的增加叶片叶绿素含量对水分响应敏感性降低,由于其与叶片光合速率的响应不一致,说明叶片净光合速率较叶绿素含量对水分的响应可能存在迟滞效应。因此,本研究结果表明水分对冬小麦叶片碳循环过程的调控主要通过调节其气孔导度、蒸腾速率和叶面饱和蒸气压差等光合生理特性指标实现。

图4 2017年和2018年冬小麦叶片净光合速率与气孔导度Fig.4 Relationships of net photosynthetic rate with stomotal conductance
3.2 施肥对冬小麦叶片净光合速率和叶绿素含量的影响
养分是限制作物生长的重要因素,研究表明大多数陆地生态系统处于氮元素和磷元素缺乏的状态[12]。施肥提高土壤中氮元素和磷元素含量,促进叶片氮含量、蛋白质、光合酶以及相关光合色素合成,从而提高植物光合作用能力。本研究显示施肥提高2017年和2018年叶绿素含量,改变量在2017年高于2018年,说明随着时间增加施肥对叶绿素含量影响减弱。但由于2018年施肥降低气孔导度,导致叶片净光合速率响应不显著,说明施肥通过调节叶片气孔导度以及叶绿素含量调控叶片净光合速率,而叶片净光合速率较叶绿素含量对施肥的响应可能存在迟滞效应。另外,施肥在整个生长季对冬小麦叶片光合速率影响不显著,主要是由于紫花苜蓿属于豆科植物,具有生物固氮作用,能提高土壤肥力,冬小麦可以利用周围土壤中苜蓿固定的氮素,使得施肥效果不显著。
短期内增水和施肥能促进小麦叶绿素含量增高,但对小麦越冬期后期到成熟期叶绿素含量作用变弱,一方面说明冬小麦叶片叶绿素含量对增水和施肥的响应在不同生长阶段不同,在出苗期、分蘖期、返青期到灌浆期植物需要更多的水分和养分,而在越冬期和成熟期作物基本不需要水分和养分的输入,另一方面也证明紫花苜蓿固定的氮素降低冬小麦对肥料的需求。增水提高冬小麦叶片净光合速率但施肥作用不显著,主要是由于在紫花苜蓿-冬小麦的草田轮作系统下,紫花苜蓿不但具有生物固氮作用增强土壤肥力,而且具有庞大的根系能保持土壤水分,缓解冬小麦对水肥的需求。同时说明在紫花苜蓿-冬小麦的草田轮作系统下,对该草田轮作系统来讲水分是最主要的限制因子,养分是次要因子。前人的研究也发现,苜蓿-小麦轮作系统,在生产实践过程中优势突出,在降低肥料输入的同时提高农民收入[13, 14]。因此,将苜蓿引入小麦的作物体系形成草田轮作系统,能肥沃土壤,减少肥料在农作物生产中的施用。另外,建立苜蓿-小麦的草田轮作系统在的能够有效地改善环境,最终实现生态和经济上的可持续发展。
4 结论
叶片净光合速率和叶绿素含量是植物叶片水平最重要的功能性状特征,准确的反应叶片水平碳循环过程对增水和施肥的响应。紫花苜蓿-冬小麦轮作体系研究结果发现增水和施肥能促进小麦叶绿素含量,但随着时间增加影响减弱。增水提高冬小麦叶片光合速率,主要是由于水分提高了气孔导度和蒸腾速率,降低了叶面饱和蒸气压差。施肥在整个生长季对冬小麦叶片光合速率作用不显著,一方面是由于施肥促进叶绿素含量提高,但降低植物气孔导度抑制光合作用原材料进入植物体,另一方面主要是因为紫花苜蓿的生物固氮作用能提高土壤肥力,降低作物对外源氮素的需求。因此,建立苜蓿-小麦的草田轮作系统具有很高的生态和经济效益,能更好的达到可持续发展的目的。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
节水灌溉(2021年5期)2021-05-26 06:49:38
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
中国农业科技导报(2020年12期)2020-03-15 10:50:08
新课程·下旬(2017年7期)2017-08-14 11:12:50
西北林学院学报(2015年5期)2015-01-03 05:12:20
赤峰学院学报·自然科学版(2012年19期)2012-10-14 02:53:58
植物营养与肥料学报(2011年2期)2011-10-26 03:52:46
