
Effects of palbociclib on oral squamous cell carcinoma and the role of PIK3CA in conferring resistance
2019-06-18 07:06NurSyafinazZainalBernardKokBangLeeZhengWeiWongIuanSheauChinPeiSanYeeChaiPheiGanKeinSeongMunZainalAriffAbdulRahmanSilvioGutkindVyomeshPatelSokChingCheong
Cancer Biology & Medicine 2019年2期
Nur Syafinaz Zainal, Bernard Kok Bang Lee, Zheng Wei Wong, Iuan Sheau Chin, Pei San Yee, Chai Phei Gan, Kein Seong Mun, Zainal Ariff Abdul Rahman, J. Silvio Gutkind, Vyomesh Patel, Sok Ching Cheong
1Head and Neck Team, Cancer Research Malaysia, Selangor 47500, Malaysia;
2Department of Oral & Maxillofacial Clinical Sciences, Faculty of Dentistry, University of Malaya, Kuala Lumpur 50603, Malaysia;
3Department of Pathology, Faculty of Medicine, University of Malaya, Kuala Lumpur 50603, Malaysia;
4Oral Cancer Research and Co-ordinating Centre (OCRCC),Faculty of Dentistry, University of Malaya, Kuala Lumpur 50603, Malaysia;
5Department of Pharmacology, University of California, San Diego 92093-5004, CA, USA
ABSTRACT Objective:Lack of effective therapies remains a problem in the treatment of oral squamous cell carcinoma (OSCC), especially in patients with advanced tumors. OSCC development is driven by multiple aberrancies within the cell cycle pathway, including amplification of cyclin D1 and loss of p16. Hence, cell cycle inhibitors of the CDK4/6-cyclin D axis are appealing targets for OSCC treatment. Here, we determined the potency of palbociclib and identified genetic features that are associated with the response of palbociclib in OSCC.Methods:The effect of palbociclib was evaluated in a panel of well-characterized OSCC cell lines by cell proliferation assays and further confirmed by in vivo evaluation in xenograft models. PIK3CA-mutant isogenic cell lines were used to investigate the effect of PIK3CA mutation towards palbociclib response.Results:We demonstrated that 80% of OSCC cell lines are sensitive to palbociclib at sub-micromolar concentrations.Consistently, palbociclib was effective in controlling tumor growth in mice. We identified that palbociclib-resistant cells harbored mutations in PIK3CA. Using isogenic cell lines, we showed that PIK3CA mutant cells are less responsive to palbociclib as compared to wild-type cells with concurrent upregulation of CDK2 and cyclin E1 protein levels. We further demonstrated that the combination of a PI3K/mTOR inhibitor (PF-04691502) and palbociclib completely controlled tumor growth in mice.Conclusions:This study demonstrated the potency of palbociclib in OSCC models and provides a rationale for the inclusion of PIK3CA testing in the clinical evaluation of CDK4/6 inhibitors and suggests combination approaches for further clinical studies.
KEYWORDS OSCC; palbociclib; CDK4/6 inhibitors; PIK3CA
Introduction
Cancers that begin in the squamous cells lining the mucosal surfaces of the head and neck region, such as the oral cavity,pharynx, larynx, salivary glands, and nasal cavity are collectively termed as head and neck squamous cell carcinoma (HNSCC). In 2012, oral squamous cell carcinoma(OSCC), the most predominant type of HNSCC, accounted for an estimated 300,400 new cases worldwide and was the 8thmost common cancer in developing countries preceding leukemia and non-Hodgkin lymphoma1. The standard treatments for OSCC used in the past decades include surgery, radiation therapy, and chemotherapy. Despite advances in these treatment modalities to improve survival and locoregional control, the 5-year survival rate remains below 60%2. Moreover, about 50% of patients with OSCC present late at stage 3 and 43, resulting in a significant drop in their survival rates4. In the past decade, the use of targeted therapy in OSCC was previously limited to cetuximab, a humanized antibody inhibiting the epidermal growth factor receptor. However, the clinical benefit of cetuximab with chemotherapy is moderate, with only 2.7 months improvement in overall survival as compared to chemotherapy alone5. Cetuximab treatment is challenged by primary and acquired resistance in patients with OSCC6.More recently, the immune checkpoint inhibitors,pembrolizumab and nivolumab, were approved for the treatment of OSCC, but the efficacy of immunotherapy is only seen in less than 20% of patients7. It is apparent that the treatment options for patients with OSCC are currently limited, especially, for patients with advanced disease and who are refractory to chemotherapy,underscoring the unmet need to increase effective treatment options for such patients.
As OSCC is largely driven by several molecular changes affecting the cell cycle, such as amplification of cyclin D1 and loss of p168, we investigated if the cell cycle inhibitor,palbociclib, would be effective in treating OSCC. Palbociclib is a specific CDK4/6 inhibitor and the first of its class to be approved for breast cancer in combination with letrozole9and fulvestrant10. In OSCC, the use of palbociclib in combination with cetuximab is still under clinical investigation11and preclinical data evaluating palbociclib are currently limited. Here, we demonstrate the effect of palbociclib on a panel of well-characterized OSCC cell lines and subcutaneous xenograft models. Using genomic information of these well-characterized models,we seek to identify molecular aberrancies that could modulate the response of OSCC to palbociclib, thereby defining new precision therapeutic options for this aggressive malignancy.
Materials and methods
Chemical reagents
Palbociclib was purchased from LC Laboratories (MA, USA).Ribociclib, abemaciclib, and PF-04691502 were purchased from Selleckchem (TX, USA). For in vitro use, all reagents were dissolved in 10 mM stocks (palbociclib and PF-04691502 in DMSO, ribociclib and abemaciclib in water) and kept as small aliquots in -20°C until further use. For in vivo experiments, palbociclib and PF-04691502 were dissolved in 133 mg/mL and 25 mg/mL stocks, respectively, in DMSO and stored at -20°C until further use.
Cell lines
The ORL series used in this study was established from patients with oral cancer as previously reported12,13. All ORL cell lines were cultured in Dulbecco’s modified Eagle’s medium/Nutrient mixture F12-Ham’s medium (DMEM/F12; Hyclone, UT, USA) supplemented with 10% (v/v) heatinactivated fetal bovine serum (FBS; Gibco, Auckland, NZ)and 500 ng/mL of hydrocortisone (Sigma-Aldrich, MO,USA). Generation of CAL27/PIK3CAH1047R+was described previously14. CAL27, CAL27/PIK3CAH1047R+and MCF-7 cells were grown in DMEM (Gibco, Auckland, NZ) supplemented with 10% (v/v) heat-inactivated FBS. All cell lines have been authenticated to tissues and/or blood from corresponding patients as described previously12. Cells were cultured at 37 °C in a 5% CO2humidified atmosphere.
Crystal violet cytostatic assay
The cytostatic effect of palbociclib on a panel of OSCC lines was determined by crystal violet cytostatic assay. Briefly, 8 ×103cells were seeded in triplicates in 12-well plates and following overnight incubation, cells were exposed to palbociclib at concentrations ranging from 0.008-2 μM or vehicle control [0.5% (v/v) DMSO]. After 72 h of incubation,crystal violet solution consisting of 2% (w/v) crystal violet(Sigma Aldrich, MO, USA), 10% (v/v) formaldehyde (Merck Millipore, MA, USA), and distilled water was added into each well and the plates were further incubated for an additional 10 min with agitation. The crystal violet solution was then removed from each well and cells were washed 2 times with water and dried overnight. The following day, the resulting crystals were dissolved in 500 μL of DMSO and the absorbance was measured at 590 nm using a Synergy H1 Multi-Mode reader (BioTek Instruments, VT, USA). The concentration of drug required to cause 50% reduction in cell growth was calculated as GI50= [(Absp- Abs0)/(AbsDMSO- Abs0)] × 100%, where Abs0is the absorbance value of untreated cells at day 0, and Abspand AbsDMSOare the absorbance values of palbociclib-treated and VC-treated cells after 72 h, respectively.
Click-iT EdU cell proliferation assay
Cell proliferation was determined by the Click-iT EdU assay(Invitrogen, CA, USA) following the manufacturer’s instructions. Briefly, 7 × 104cells per well were grown overnight on glass coverslips and treated with 0.06-0.5 μM palbociclib or 0.5% (v/v) DMSO for 24 h. Cells were incubated with 10 μM 5-ethynyl-2′-deoxyuridine (EdU) for 2-6 h prior to fixation with 3.7% (v/v) formaldehyde. The cells were permeabilized with 0.1% (v/v) Triton X-100 in phosphate buffer, followed by EdU detection via a coppercatalyzed reaction and nuclei staining by Hoechst 33342. The coverslips were then mounted on glass slides using VECTASHIELD® Mounting Medium (Vector Laboratories,Burlingame, CA, USA). Slides were examined on an upright Olympus IX71 microscope (Olympus, Japan) with double bandpass filters to detect fluorescent-stained nuclei (Hoechst 33342-excitation 360-370 nm and emission 420 nm) and Alexa-labeled EdU (Alexa 647: excitation 650 nm and emission 667 nm). Images were captured from 10 randomly chosen fields of each experiment and analyzed with the QuickCount® software. The number of EdU-positive cells and Hoechst 33342-stained cells was counted and the percentage of EdU-positive cells was calculated (from three independent experiments) using the following formula:number of EdU positive cells/number of Hoechst 33342-stained cells × 100. EdU-positive cells broadly represent cells that are undergoing DNA synthesis, whereas Hoechst 33342-stained cells represent all cells in the same field.
Cell cycle assay
Briefly, 7 × 104cells were seeded per well in 12-well plates and treated with 0.06-0.5 μM palbociclib or 0.5% (v/v)DMSO on the following day for 24 h. All floating and attached cells were harvested and fixed in 70% (v/v) ethanol for 16 h at -20°C. Prior to analysis, fixed cells were pelleted and washed in cold phosphate buffered saline, followed by staining with 10 μg/mL propidium iodide solution containing 20 μg/mL RNase for 30 min at 21°C in the dark.Stained cells were analyzed by BD FACSCanto IITMflow cytometer (BD Biosciences, MA, USA) with 10, 000 events collected for each reading. The distribution of DNA in different phases was determined using the ModFit software(Verity Software House, USA). The percentage of cells in each phase was calculated from three independent experiments.
Western blot
Palbociclib (0.06-0.5 μM)-treated and 0.5% (v/v) DMSOtreated cells were lysed on ice in lysis buffer [5 M NaCl, 10%(v/v) NP-40, 1 M Tris pH 8.0, and 0.5 mM DTT]supplemented with HALT protease and phosphatase inhibitor cocktail (Pierce Biotechnology, IL, USA). Cell lysates were then centrifuged at 13,000 × g for 10 min at 4°C prior to estimation of protein content using the BCA method(Thermo Fisher Scientific, MA, USA). For Western blot analysis, 30 μg of total cellular proteins was resolved on a 12% (w/v) sodium dodecyl sulfate-polyacrylamide gel and electrotransferred onto Immobilon-P membrane (PVDF;Millipore, MA, USA) at 100 V for 1.5 h on ice. Membranes were blocked with 5% (v/v) skimmed milk in Tris-buffered saline with 0.1% (v/v) Tween 20 (TBST; Sigma-Aldrich, MO,USA) for 1 h and then probed overnight at 4°C with the indicated primary antibodies at 1:1000 dilution in 1% (v/v)bovine serum albumin in TBST [pRb (S780), pRb (S795),total Rb1, cyclin D1, cyclin E1, CDK2, CDK4, CDK6, pAKT(S473), total AKT1, pS6, total S6 (Cell Signaling Technology,MA, USA), and α-tubulin (Sigma-Aldrich, MO, USA)]. Next,membranes were washed 3 times in TBST for 5 min each.Membranes were then incubated with the corresponding horseradish peroxidase (HRP)-conjugated secondary antibodies (Southern Biotech, AL, USA) at 1:10,000 dilution in 5% (v/v) skimmed milk in TBST for 1 h at room temperature. This was followed by 3 washes in TBST prior to detection by WesternBright Quantum HRP substrate(Advansta Inc., CA, USA) and visualization using the FluorChemTMHD2 imaging system (ProteinSimple, CA,USA).
In vivo mouse experiments
Six-week-old female NOD/SCID mice were subcutaneously implanted with 2 × 106cells in both flanks. When the tumors reached the size of approximately 100-200 mm3, the mice were randomized into experimental groups: vehicle control(VC), palbociclib (150 mg/kg body weight, suspended in DMSO 5% and Cremophor-EL 10%, 500 μl p.o), or PF-04691502 (10 mg/kg body weight, suspended in DMSO 4%and 0.5% methylcellulose, 200 μL p.o). Treatment was administered daily by oral gavage for 21 days and tumor measurements were taken using a digital calliper every 3-4 days throughout the experiment. Tumor volumes (V) were calculated as V = ½ length × (width)2. Growth curves were plotted as an average tumor volume of each experimental group against the set time points. At the end of experiment,mice were sacrificed and tumors were harvested for tissue processing and histopathological analysis. All procedures involved in the animal study were reviewed and approved by the Animal Ethics Committee of Universiti Kebangsaan Malaysia (Approval No. CARIF/2016/CHEONG/18-MAY/762-JUNE-2016-JUNE-2019).
Statistical analysis
All data are expressed as mean ± standard deviation (SD)from three independent experiments unless stated otherwise.Statistical analysis was performed using IBM SPSS Statistics for Windows, version 21.0 (IBM Corp., NY, USA).Significant differences between groups were analyzed using 2-tailed independent t-test or one-way analysis of variance(ANOVA). A P-value less than 0.05 was considered statistically significant.
Results
Palbociclib inhibits the growth of OSCC cells in culture
To examine the effect of palbociclib on OSCC, we treated a panel of OSCC cell lines with 0-2 μM of palbociclib for 72 h.Among the 16 OSCC cell lines tested, 13 were sensitive to palbociclib with a GI50(concentration for 50% of maximal inhibition of cell growth) of less than 1 μM, while three lines with a GI50of more than 1 μM indicated resistance to palbociclib based on the cytostatic assay (Figure 1A). The sensitive and resistant groups were significantly different from each other with ~5-fold differences in average GI50values (P < 0.01). To further validate the inhibitory effect of palbociclib on cell proliferation, we performed Click-iT EdU proliferative assay on representative palbociclib-sensitive cells(CAL27 and ORL-48) and palbociclib-resistant cells (ORL-150 and ORL-115). Relative to the VC group, palbociclib treatment (0.06 to 0.5 μM) for 24 h reduced the number of EdU-positive cells (represented as red stained-cells) in CAL27 and ORL-48 cells (Figure 1B), suggesting a decrease in DNA synthesis activity and blockade of cell cycle progression from G1to S phase. As expected, the reduction in EdU-positive cells was not as pronounced in ORL-150 and ORL-115 cells where the reduction in dividing cells was less than 50% even at the highest drug concentration (Figure 1B and 1C).
Anti-proliferative activity of palbociclib is due to G1 cell cycle arrest
As a selective inhibitor of CDK4/6, palbociclib is known to prevent the initiation of S phase of the cell cycle. To confirm this, we next investigated the anti-proliferative effect of palbociclib through cell cycle analysis following propidium iodide staining in palbociclib-sensitive CAL27 and ORL-48 cells. The cells were treated with palbociclib (0.06 to 0.5 μM)for 24 h. Palbociclib treatment increased the percentage of cells in the G0/G1phase, and concurrently reduced the percentage of S phase cells in a dose-dependent manner for both cell lines (Figure 1D). This indicates the inhibition of S phase initiation by palbociclib, which resulted in the arrest of cell cycle progression at the G0/G1phase in OSCC cells.Palbociclib did not induce death in CAL27 and ORL-48 cells as seen by the constant level of cell population in the sub-G1phase across the different treatment doses (Supplementary Figure S1).
Palbociclib reduces Rb protein levels and phosphorylation
We further investigated the biochemical effect of palbociclib on proteins involved in the Rb pathway, which is critical in regulating the initiation of DNA replication. We first performed a dose-dependent protein expression analysis on palbociclib-sensitive (CAL27 and ORL-48) and palbociclibresistant cells (ORL-150 and ORL-115) for Rb phosphorylation at S780, S795, and S807/811 as these sites are known to be phosphorylated by CDK4/615. We observed that treatment with palbociclib, especially at 0.5 μM, reduced Rb phosphorylation at all three phosphorylation sites in CAL27, ORL-48, and ORL-150 cells (Figure 1E), indicating the blockage of cell cycle progression upon palbociclib treatment. In contrast, ORL-115 cells retained the expression and phosphorylation of Rb at all doses tested (0.06-0.5 μM),validating the resistance of this cell line towards palbociclib,as shown in the proliferative assays described above. As the greatest effect of Rb dephosphorylation was observed at 0.5 μM dose, this concentration was used to conduct a timedependent protein expression analysis of palbociclib-sensitive cells (CAL27 and ORL-48). Complete dephosphorylation of Rb was observed after 24 h of palbociclib treatment,particularly at S780 and S795 (Figure 1F). Additionally,palbociclib steadily downregulated the expression levels of Rb in a time-dependent manner, culminating in low Rb expression at 24 h after treatment in both cell lines. We also noted that the protein levels of cyclin D1, CDK4, and CDK6 were slightly increased with time as a result of palbociclib treatment, consistent with observations in other studies on breast and colorectal cancers16,17.
Palbociclib inhibits tumor growth in subcutaneous xenograft model
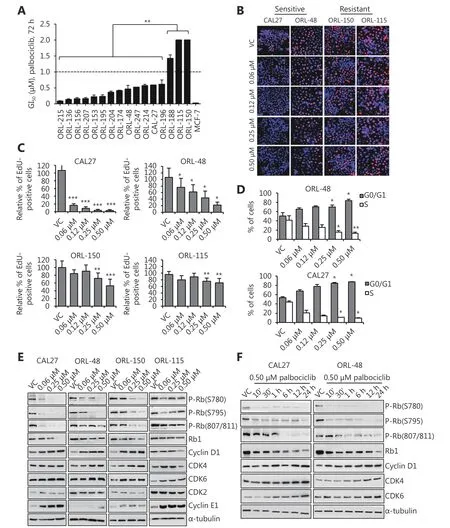
Figure 1 Majority of OSCC cell lines were sensitive to palbociclib. (A) Crystal violet cytostatic assay performed on a panel of OSCC cell lines showed GI50 of less than 1 μM in 80% of the cell lines tested, while three lines (ORL-188, ORL-150 and ORL-115) indicated resistance(GI50>1μM). Cell lines were treated with 72h of 0.008 to 2 μM of palbociclib. (B,C) Cytostatic effect of palbociclib was validated in representative sensitive and resistant cell lines using Click-iT EdU assay. Blue (Hoecsht 33342) represents the total number of cells in any field and red (Alexa 647) represents proliferating cells that have incorporated the EdU label. Palbociclib (24 h) exhibits anti-proliferative effects on CAL27 and ORL-48 as demonstrated by the reduction in the percentage of EdU-positive cells which represents actively dividing cells, but not observed in ORL-115. (D) Escalating dose of palbociclib (24 h) increased G0/G1 cell population in CAL27 and ORL-48 cells by propidium iodide analysis. (E) Dose-dependent Western blot analysis of palbociclib showed reduction of Rb phosphorylation (greatest at 0.5 μM)on CAL27, ORL-48 and ORL-150 but not on ORL-115. Cells were treated with the indicated concentrations for 24 h. (F) Time-dependent Western blot analysis on sensitive cells indicated reduction of Rb phosphorylation within 24 h at 0.5 μM palbociclib treatment. Symbols *, **and *** denote P < 0.05, P < 0.01, P < 0.001, respectively. Western blot results shown are representative of at least two independent experiments.
As demonstrated earlier, palbociclib exerted potent inhibitory effect on OSCC cell growth in vitro. Next, we investigated in vivo efficacy of palbociclib in subcutaneous xenograft models using CAL27 and ORL-48 cells. Mice were randomized to receive either 150 mg/kg of palbociclib or VC when tumors reached an average volume of 100-150 mm3.We observed that palbociclib treatment for 21 days significantly inhibited the growth of CAL27 (VC: 477.5 ±101.2 mm3vs. palbociclib: 159.76 ± 22.6 mm3; P = 0.007)and ORL-48 (VC: 755.06 ± 51.07 mm3vs. palbociclib: 68.47± 15.07 mm3, P = 1 × 10-7) xenograft tumors (Figure 2A and 2B). Next, we investigated the expression and phosphorylation of Rb protein in xenograft tumors that were harvested at the end of the study. As shown in Figure 2C,palbociclib treatment resulted in the reduction in Rb phosphorylation at S795 in CAL27 xenograft tumors. The effect was more apparent in ORL-48 xenograft tumors,wherein reduction in Rb phosphorylation was observed at both S780 and S795 sites, consistent with the observation that tumor volume control was more significant in this cell line compared to that in CAL27 cells (Figure 2A and 2B).Although we observed marked reduction in Rb phosphorylation at S780 and S795, Rb phosphorylation was not affected at S807/811 (Figure 2C), which was consistent with in vitro observations for ORL-48 (Figure 1E). In addition, palbociclib treatment caused reduction in the total Rb levels of both CAL27 and ORL-48 tumors, consistent with that observed in vitro. These observations confirmed that palbociclib inhibited the CDK4/6-Rb pathway, which eventually led to tumor control in mice.
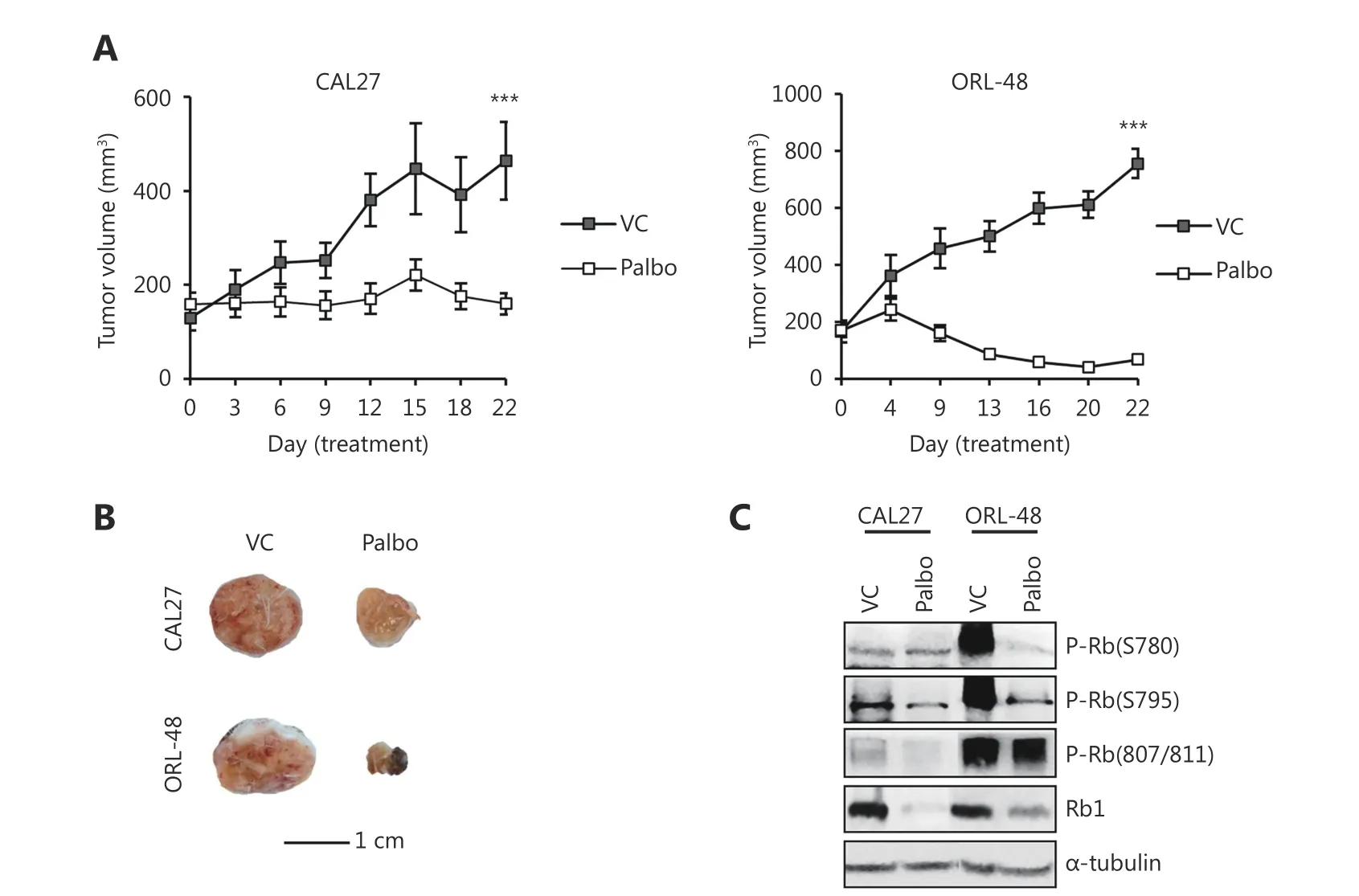
Figure 2 Palbociclib controls OSCC xenograft tumors. (A) Mice bearing CAL27 and ORL-48 xenograft tumors were treated with either vehicle control (VC) or 150 mg/kg palbociclib (Palbo) for 21 d. Palbociclib significantly inhibited growth of OSCC subcutaneous xenograft ORL-48 and CAL27 tumors. Results shown are mean ± SEM (n = 10 tumors per group). Symbol *** denotes P < 0.001. (B) Representative tumors harvested at the end of treatment showed decreased tumor volume in the palbociclib treatment group. (C) Western blot analysis on proteins extracted from tumors at end-point showed palbociclib treatment reduced Rb phosphorylation in mouse tumors particularly at S780 and S795 phosphorylation sites.
Cancer cells harboring PIK3CA mutation are less responsive to palbociclib
From our data, we observed that a subset of OSCC cell lines was resistant to palbociclib. We investigated whether certain genetic features could lead to palbociclib resistance. To achieve this, we compared the mutational profiles of palbociclib-sensitive and -resistant cell lines as determined by cytostatic assay in Figure 1A. We found that two out of three cell line in the resistant group carried mutations in PIK3CA(H1047L and Q546R in ORL-115 and ORL-150,respectively)12,18, while none of the palbociclib-sensitive cell lines had any mutations in this gene (P < 0.001; Figure 3A).This observation was validated with a larger data set from Genomics of Drug Sensitivity in Cancer (GDSC) comprising of 789 palbociclib-treated cancer cell lines of all available cancer types in the database. These cell lines were grouped based on their PIK3CA mutational status [wild-type (WT) or having any mutations in PIK3CA] and their geometric means of IC50were compared. Cancer cells having mutation in PIK3CA have a higher geometric mean IC50, implying that they are more resistant to palbociclib compared to cell lines with WT PIK3CA; however, the difference was not statistically significant (P = 0.101; Figure 3B). This led us to further investigate the role of PIK3CA mutations in conferring palbociclib resistance in OSCC.

Figure 3 Palbociclib-resistant cell lines harbour PIK3CA mutation. (A) Comparison of the mutational status between sensitive and resistant cells obtained from the cytostatic data of Figure 1A revealed significant association of PIK3CA mutation with palbociclib resistance. Symbol*** denotes P < 0.001. (B) According to Genomics of Drug Sensitivity in Cancer (GDSC) database, cancer cells having any mutation in the PIK3CA gene have higher geometric mean IC50, implying them to be more resistant to palbociclib compared to wild-type cancer cells. A total number of 789 palbociclib-treated cancer cell lines of all available cancer types are included in the analysis.
PIK3CA mutation decreases response to palbociclib
To test whether PIK3CA mutations confer resistance to palbociclib in OSCC, we used CAL27 cells that were previously retro-engineered to stably express PIK3CAH1047Ralteration14. The mutational status of PIK3CA in isogenic(mutant and WT) CAL27 cell lines was verified by Sanger sequencing, where the mutant CAL27 cells have base substitution from A (WT) to G, resulting in the replacement of histidine (H) with arginine (R) at position 1047(Supplementary Figure S2A). The presence of the H1047R mutation was also confirmed by western blotting, wherein increased Akt phosphorylation was observed in mutant cells(Supplementary Figure S2B).
The differential sensitivity to palbociclib in these isogenic cells was first evaluated by EdU proliferation assay. The inhibition of cell proliferation was more pronounced in WT cells as the percentage of EdU-positive cells was observed to be lower by ~4-fold compared to that in PIK3CA mutant cells at a dose of 0.5 μM (P < 0.001; Figure 4A and 4B). In addition, we assessed whether the presence of PIK3CA mutation would affect the response to two other clinically approved CDK4/6 inhibitors19,20. We treated isogenic CAL27 cells with ribociclib and abemaciclib (0.25-0.5 μM) and found that the number of EdU-positive cells was also significantly lower in WT cells as compared to that in PIK3CA mutant cells (Figure 4C). These results indicated that the presence of PIK3CA mutation reduced the sensitivity of CAL27 to CDK4/6 inhibitors, and this observation was not restricted to palbociclib.
Next, flow cytometry was performed to evaluate the differences in G1cell cycle arrest in the respective WT and mutant cells. As demonstrated previously, palbociclib treatment arrests cells at the G0/G1phase. As shown in Figure 4D, the percentage of cells in the G0/G1phase for PIK3CA mutant cell line was lower than that for WT cells at all doses tested, with statistical significance at 0.5 μM palbociclib (P =0.02). Concurrently, PIK3CA mutant cell lines showed greater percentage of cells in the S phase than WT cells at 0.5 μM treatment dose (P = 0.03), indicating higher cell cycle activity, consistent with the results of palbociclib resistance.
PIK3CA mutant cells escape CDK4/6 inhibition via the CDK2 pathway
To investigate the biochemical mechanism that caused PIK3CA mutant cells to exhibit higher resistance to palbociclib, proteins were extracted from the isogenic cells after 24 h of treatment with escalating doses of palbociclib(0.06 to 0.5 μM), and proteins within the Akt and Rb pathway were examined by Western blot (Figure 4E). Due to the presence of the PIK3CAH1047Rmutation, mutant cells showed significantly higher phosphorylation of Akt as compared to WT cells across all palbociclib doses tested. The reduction in Rb phosphorylation, particularly at S795 and S807/811, was not as marked in PIK3CA mutant cells treated with palbociclib as compared to that in WT cells. Less reduction in Rb phosphorylation at the S780 site was also detected; however, it was only apparent at the 0.06 μM treatment dose. Lack of reduction in Rb phosphorylation at S807/811 and S795, but clear reduction at S780 in the PIK3CA mutant cells suggested an S phase entry through phosphorylation of Rb by the CDK2/cyclin E1 complex15,21,22.

Figure 4 PIK3CA-mutant cells were less inhibited by palbociclib. (A, B) Less inhibition by palbociclib (24 h) was observed in PIK3CAmutated CAL27 cells as indicated by the increased percentage of dividing cells as compared to WT by Click-iT EdU assay. Blue (Hoechst 33342) represents the total number of cells in any field and red (Alexa 647) represents proliferating cells that have incorporated the EdU label. (C) PIK3CA-mutated CAL27 cells were less sensitive to all available specific CDK4/6 inhibitors (palbociclib, ribociclib and abemaciclib)at 0.25 and 0.5 μM by Click-iT EdU assay. (D) Reduced G0/G1 and increased S phase population were seen in PIK3CA-mutated cells as compared to WT by cell cycle (propidium iodide) analysis, following 24 h palbociclib treatment. (E) Western blot analysis showed less reduction of Rb phosphorylation and expression in PIK3CA-mutated cells as compared to WT upon 24 h palbociclib treatment. Increased expressions of CDK2 and Cyclin E1, but no changes in CDK4, CDK6 and cyclin D1 suggested S phase entry through CDK2/cyclin E complexes in PIK3CA-mutated cells. Symbols *, ** and *** denote P < 0.05, P < 0.01, and P < 0.001 respectively. Western blot results shown are representative of at least two independent experiments.
We noted that palbociclib treatment (0.06 to 0.5 μM) caused reduction in CDK2 expression in WT cells, however CDK2 expression was sustained throughout the range of palbociclib dose tested in the mutant cells. Cyclin E1, which partners with CDK2 to phosphorylate Rb, showed increased levels of expression in the mutant cells as compared to those in WT cells when palbociclib treatment was given. On the contrary,we did not see any changes in CDK4, CDK6, and cyclin D1 protein levels between WT and mutant cells across all treatment doses, suggesting that the CDK2/cyclin E1 pathway is a potentially compensatory pathway that led to Rb phosphorylation in PIK3CA mutant cells, resulting in an escape from palbociclib treatment.

Figure 5 Palbociclib-resistant cell line was re-sensitized when co-inhibited with a PI3K/mTOR inhibitor. (A) From Click-iT EdU assay, ORL-115 (naturally carrying a H1047L PIK3CA mutation) cells showed resistance to 0.5 μM palbociclib (24 h) in relative to CAL27 WT cells.Combining the treatment with 0.25 μM PF-04691502 (a PI3K/mTOR inhibitor) significantly improved growth inhibition in ORL-115 cells.Symbol *** denotes P < 0.001. (B) Western blot analysis of proteins extracted from ORL-115 cells treated with 0.5 μM palbociclib and 0.25 μM PF-04691502 (24 h) showed non-phosphorylated Rb and prevention of cell cycle kinases upregulations. (C) Mice bearing ORL-115 xenograft tumor were treated with either VC, palbociclib (150 mg/kg), PF-04691502 (10 mg/kg), or combination of both for 21 d. Only the combination treatment resulted in the gradual decrease of tumor volumes. Results shown are mean ± SEM (n = 10 tumors per group). (D)Representative ORL-115 xenograft tumor harvested at the end of experiment showed the greatest tumor inhibition from the combination group.
Combination of CDK4/6 and PI3K inhibitors sensitizes the PIK3CA mutant cell line ORL-115
Since PIK3CA mutations confer resistance to palbociclib, we used the inherently resistant cell line ORL-115 to determine whether PIK3CA mutant cells could be sensitized to palbociclib with a combination treatment targeting the PI3K pathway. Using ORL-115, which harbors the PIK3CAH1047Lmutation, we showed that treatment with a combination of palbociclib and a PI3K/mTOR inhibitor (PF-04691502)substantially increased sensitivity to palbociclib, where the lowest percentage of EdU-positive cells was observed in the combination group (Figure 5A). Consistent results were observed for ORL-150 (data not shown). The Western blot results in ORL-115 cells indicated that treatment with palbociclib alone resulted in the upregulation of cyclin E1 and sustained levels of CDK2, possibly leading to Rb phosphorylation especially at S795 and S807/811 sites(Figure 5B). Inhibiting both CDK4/6 and PI3K pathways prevented the upregulation of cyclin E1, reduced CDK2 levels, and resulted in the accumulation of nonphosphorylated Rb at all three sites, suggesting improved growth inhibition in ORL-115 cells. Additionally, cotreatment with PF-04691502 resulted in the reduction in levels of other cell cycle kinases, such as CDK4, CDK6, and cyclin D1 as compared to palbociclib treatment alone.Consistently, in vivo, we demonstrated that ORL-115 cells were less responsive to palbociclib, as sustained tumor growth was still observed in animals treated with palbociclib,with tumor volume steadily increasing from 112 mm3(day 1)to 296 mm3(day 22) throughout the treatment period(Figure 5C). Only when the combined treatment was given, a sustained tumor volume reduction was achieved from 124 mm3(day 1) to 76 mm3(day 22). ORL-115 tumors that were treated with PF-04691502 also demonstrated better tumor growth control compared to those treated with palbociclib; however, this was not as marked as compared to the combination treatment of palbociclib and PF-04691502(Figure 5C and 5D).
Discussion
Alterations within the cell cycle machinery are common molecular alterations found in HNSCC, including mutations in TP53 (74%), loss of p16 (49%), and amplification of CCND1 (28%)8. These alterations implicate the use of cell cycle inhibitors in OSCC. In recent years, cell cycle inhibitors with targeted activity towards specific components of the cell cycle have been developed and approved in a clinical setting for breast cancer19,23. Here, we describe the effects of the CDK4/6 inhibitor palbociclib on OSCC cell lines and tumor models. We found that majority of the OSCC cell lines responded to palbociclib treatment with concomitant reduction in RB phosphorylation, which resulted in inhibition of cell proliferation and G1cell cycle arrest. The in vitro observations were confirmed by significant tumor control in subcutaneous mouse models. A recent Phase I clinical trial on OSCC also reported tumor response in patients with OSCC treated with palbociclib, including those previously resistant to cetuximab or cisplatin11, strongly suggesting that palbociclib could be a promising therapeutic drug for patients with OSCC.
Understanding the molecular changes that could influence therapeutic response is vital for identifying subgroups that are most likely to benefit from palbociclib treatment. We leveraged on genomic information on well-characterized cell lines12,18,24and showed that PIK3CA mutations are associated with the resistance of OSCC cell lines to palbociclib. Using CAL27 isogenic cell lines, we confirmed that mutations in PIK3CA indeed confer resistance to palbociclib and other clinically approved CDK4/6 inhibitors. In PIK3CA mutant cell lines, this resistance is accompanied by an increase in cyclin E and CDK2 expression. PIK3CA mutations have been shown to increase the expression of cyclins and their dependent kinases. For example, in a non-tumorigenic human breast epithelial cell line, PIK3CA mutations resulted in the upregulation of cyclin D1, resulting in an increase in cell proliferation25. Furthermore, upregulation of cyclin D1 and CDK2-dependent Rb phosphorylation by PI3K activation was also reported in MCF7 cells in response to CDK4/6 inhibition16. PIK3CA mutations were also previously reported to increase cyclin E expression, where PI3Kdependent upregulation of cyclin E1 levels was associated with acquired resistance to CDK4/6 inhibition in pancreatic ductal adenocarcinoma26. PI3K activation can drive the cell cycle by regulating the expression of cell cycle inhibitors,including p21 and p27, as reported previously27-29.Furthermore, PIK3CA mutations could directly regulate cyclin D levels by inhibiting glycogen synthase kinase-3β expression, resulting in the stabilization and localization of cyclin D1 to the nucleus25,30. Based on our observation that activation of PI3K confers resistance to palbociclib, we demonstrated that treatment with a combination of palbociclib and a PI3K/mTOR inhibitor resulted in complete tumor growth control in the animal model. While PI3K/mTOR inhibition alone was also able to delay tumor growth, it was obvious that blocking PI3K signaling restored sensitivity to palbociclib in OSCC models. Concurring with our in vitro observations, complete tumor growth control was observed concomitantly with reduction in cyclin E1 and CDK2 expression. This is consistent with data from other studies where PI3K/mTOR inhibitor synergistically cooperated with CDK4/6 inhibitor in suppressing cell proliferation by reducing the levels of critical cell cycle regulatory proteins, such as cyclin E1, cyclin D1, CDK2, and cyclin A26,31,32.
In summary, we demonstrated that palbociclib is effective in controlling tumor growth in most OSCC cell lines. Our data also revealed that PIK3CA mutational status may serve as a biomarker for response to palbociclib and other clinically approved CDK4/6 inhibitors. Finally, we demonstrated that simultaneous inhibition of PI3K/mTOR and CDK4/6 is effective in controlling tumor growth in resistant OSCC cell lines. Given that clinical studies on CDK4/6 inhibitors in OSCC have been initiated, our study provides a rationale for testing biomarkers of response and suggests possible combinatorial approaches for patients who may not respond to CDK4/6 inhibitors as single agents.
Acknowledgements
This study was supported by a grant from High Impact Research, Ministry of Higher Education (HIR-MOHE) from University of Malaya (Grant No. UM.C/625/1/HIR/MOHE/DENT-03) and Cancer Research Malaysia.
Conflict of interest statement
No potential conflicts of interest are disclosed.
 Cancer Biology & Medicine2019年2期
Cancer Biology & Medicine2019年2期
- Cancer Biology & Medicine的其它文章
- Hepatitis B virus X protein enhances hepatocarcinogenesis by depressing the targeting of NUSAP1 mRNA by miR-18b
- LAG-3 expression on tumor-infiltrating T cells in soft tissue sarcoma correlates with poor survival
- Genetic polymorphisms and gastric cancer risk: a comprehensive review synopsis from meta-analysis and genome-wide association studies
- Identification of anticancer drugs to radiosensitise BRAFwild-type and mutant colorectal cancer
- Cancer stem-like cells directly participate in vasculogenic mimicry channels in triple-negative breast cancer
- Factors associated with upstaging in patients preoperatively diagnosed with ductal carcinoma in situ by core needle biopsy