
LAG-3 expression on tumor-infiltrating T cells in soft tissue sarcoma correlates with poor survival
2019-06-18 07:06YiQueZhixinFangYuanxiangGuanWeiXiaoBushuXuJingjingZhaoHuoyingChenXinkeZhangMushengZengYaoLiangXingZhang
Cancer Biology & Medicine 2019年2期
Yi Que, Zhixin Fang, Yuanxiang Guan, Wei Xiao, Bushu Xu, Jingjing Zhao, Huoying Chen, Xinke Zhang,Musheng Zeng, Yao Liang, Xing Zhang
1Department of Medical Melanoma and Sarcoma, State Key Laboratory of Oncology in South China, Collaborative Innovation Center for Cancer Medicine, Sun Yat-sen University Cancer Center, Guangzhou 510060, China;
2Department of Laboratory Medicine and Central Laboratories, Guangdong Second Provincial General Hospital, Guangzhou 510000, China;
3Department of Gastric and Pancreatic Surgery, State Key Laboratory of Oncology in South China, Collaborative Innovation Center for Cancer Medicine, Sun Yat-sen University Cancer Center, Guangzhou 510060, China;
4Department of Pathology,State Key Laboratory of Oncology in South China, Collaborative Innovation Center for Cancer Medicine, Sun Yat-sen University Cancer Center, Guangzhou 510060, China;
5State Key Laboratory of Oncology in South China, Collaborative Innovation Center for Cancer Medicine, Sun Yat-sen University Cancer Center, Guangzhou 510060, China
ABSTRACT Objective:To elucidate the role and prognostic significance of lymphocyte activation-gene-3 (LAG-3) in soft tissue sarcoma(STS).Methods:The expression of LAG-3 in patient and matched normal blood samples was analyzed by flow cytometry. The localization and prognostic values of LAG-3+ cells in 163 STS patients were analyzed by immunohistochemistry. In addition, the expression of tumor-infiltrating CD3+ T, CD4+ T, and CD8+ T cells and their role in the prognosis of STS were evaluated by immunohistochemistry. The effect of LAG-3 blockade was evaluated in an immunocompetent MCA205 fibrosarcoma mouse model.Results:Peripheral CD8+ and CD4+ T cells from STS patients expressed higher levels of LAG-3 than those from healthy donors.LAG-3 expression in STS was significantly associated with a poor clinical outcome (P = 0.038 ) and was correlated with high pathological grade (P < 0.001), advanced tumor stage (P = 0.016). Additionally, LAG-3 expression was highly correlated with CD8+ T-cell infiltration (r = 0.7034, P < 0.001). LAG-3 was expressed in murine tumor-infiltrating lymphocytes, and its blockade decreased tumor growth and enhanced secretion of interferon-gamma by CD8+ and CD4+ T cells.Conclusions:LAG-3 blockade may be a promising strategy to improve the effects of targeted therapy in STS.
KEYWORDS Soft tissue sarcoma; LAG-3; expression; prognosis; tumor-infiltrating lymphocytes
Introduction
Soft tissue sarcomas (STS) are a heterogeneous group of rare mesenchymal tumors, which account for approximately 1%of all adult cancers1. Surgery is the primary treatment for localized STS. Anthracycline, either as a single agent or in combination with ifosfamide, is a conventional treatment for advanced or metastatic STS2. However, the prognosis of metastatic or advanced STS remains poor3,4. In the last decade, the utility and tremendous success of immunotherapy in metastatic melanoma and renal cell cancer has been demonstrated. In recent years, responses in“non-immunogenic” cancers, including bladder and lung cancers, have highlighted the potential for immunotherapy in STS. Additionally, sarcomas are among the first tumors to be considered for immunotherapeutic investigations. However,from the application of Coley’s toxin in the early 1900s5,6to the present promising strategy of using T-cell therapy in synovial sarcoma expressing NY-ESO-1, immunotherapy unquestionably induces certain but limited responses in STS7.
Immune checkpoint inhibitors have enabled new strategies for cancer treatment and opportunities to investigate and explore STS8-10. Targeting immune inhibitory checkpoints,such as programmed cell death protein 1 (PD-1) and cytotoxic T-lymphocyte-associated antigen 4 (CTLA-4),induces durable responses and potential antitumor effects against various malignancies11-14. An open-label, single-arm phase II study (SARC028) using the anti-PD-1 antibody pembrolizumab in patients with advanced soft-tissue and bone sarcomas showed promising results in specific histological subtypes15. However, the primary endpoint of an overall response was not met in this clinical trial. Thus, other novel immune inhibitory molecules must be identified for STS treatment. As one of the next generation of inhibitory checkpoints, lymphocyte activation-gene-3 (LAG-3) is coordinately upregulated in exhausted or dysfunctional T cells in cancer16,17. LAG-3 protein binds to a nonholomorphic region of major histocompatibility complex 2(MHC class II)18and interacts with ligand designated LSECtin to dampen antitumor immunity by inhibiting interferon-gamma (IFNγ) secretion from antigen-specific effector T cells19.
Recent studies have demonstrated LAG-3 expression in various cancer types. In chronic lymphocytic leukemia, high LAG-3 expression indicates poor treatment outcomes20.Increased LAG-3 expression was observed in tumorinfiltrating lymphocytes (TILs) in hepatocellular carcinoma21. In gastric cancer, the expression of LAG-3 on CD4+and CD8+T cells is elevated and might impair cellmediated immunity22. LAG-3 is also expressed on CD8+PD-1+TILs in melanoma, and a blocking antibody was reported to restore the function of tumor-reactive lymphocytes23.Additionally, targeting of LAG-3 and PD-1 could potentially improve the antitumor function of NY-ESO-1-specific CD8+T cells in human ovarian cancer17. Moreover, dual blockade of LAG-3 and PD-1 improved the proliferation and cytokine production of tumor antigen-specific CD8+T cells in preclinical cancer models24. However, the association between LAG-3 and STS outcomes has not been reported. It remains unknown whether LAG-3 plays a role in STS and whether LAG-3 blockade can reactivate the antitumor response of T cells.
Advanced and metastatic STS remains a deadly disease and immunologic treatments demonstrate a promising therapeutic approach. Thus, it is important to define the immune-related biomarkers that may predict a patient’s prognosis and further guide immunotherapy. In the present study, we aimed to investigate the expression and prognostic significance of the novel immune checkpoint receptor, LAG-3, in STS and to preliminarily explore the role of anti-LAG-3 immunotherapy by analyzing tumor growth in the MCA205 sarcoma model.
Materials and methods
Patients
Peripheral blood was isolated from 12 advanced STS patients and 11 matched healthy donors. Peripheral blood mononuclear cells were collected by density centrifugation.Tumor specimens and matched peripheral blood were obtained from two advanced STS patients from Sun Yat-sen University who consented to a protocol approved by the institutional review board (IRB).
A cohort of 163 primary patients with STS who had undergone extensive and radical resection at the Sun Yat-sen University Cancer Center from 2000 to 2010 was enrolled.The patients’ characteristics are listed in Supplementary Table S1. The median patient age at resection was 39 years(range, 5-77 years). The patients were followed-up for 1-176 months, with a median of 75 months. Written informed consent was obtained from each patient, and ethical approval was given by the medical ethics committee of the Sun Yat-sen University Cancer Center IRB (Approval No. YB2017-30).
TCGA sarcoma samples
The gene level 3 mRNA data were downloaded from The Cancer Genome Atlas (TCGA) portal, which is publicly accessible25. The TCGA mRNA expression data were then normalized using the RSEM algorithm.
Flow cytometry and cytokine detection
Single-cell suspensions isolated from tumors and spleen were acquired as previously described26. In brief, tumors were dissociated by sufficient mincing in complete RPMI 1640 medium followed by collagenase Type IV (Sigma-Aldrich)and DNase I (Sigma-Aldrich) digestion for 1 hour at 37°C and filtering through 70-μm nylon cell strainers in phosphate buffer saline. The cells were immediately analyzed by flow cytometry. Human-specific antibodies were purchased from ENZO Life Science (LAG-3, 17B4) and BD Pharmingen(CD3, CD4, CD8). Isotype controls were purchased from BD Pharmingen. Mouse-specific antibodies were purchased from eBioscience (Cd3e, Cd4, and Cd8a). Intracellular staining for IFN-γ was performed using the Fixation and Permeabilization Buffer Kit (BD Bioscience). Cells were sorted using the Beckman flow cytometer, and the data were analyzed using FlowJo software.
Immunohistochemical and immunofluorescence staining
Serial 4-μm formalin-fixed, paraffin-embedded tissue sections from primary STS were stained manually. The antibodies used for immunohistochemical and immunofluorescence staining are listed in Supplementary Table S2.
For immunohistochemical staining, each section was dewaxed and rehydrated using xylene and then washed using a graded alcohol series. Antigen retrieval was performed for 2.5 min at high pressure using sodium citrate. The sections were incubated with primary antibodies against human CD3,CD4, CD8 or LAG-3, and then with goat secondary antibody against rabbit and mouse immunoglobulins (DAKO) for 30 min at 37°C.
For immunofluorescence staining, human tissue samples were fixed with 4% paraformaldehyde and embedded in paraffin. The sections were then dewaxed and rehydrated using xylene and washed using a graded alcohol series.Antigen retrieval was performed for 2.5 min using sodium citrate. Next, the sections were blocked with bovine serum albumin for 30 min at 37°C and incubated with LAG-3 and CD8 antibodies at 4°C overnight at the indicated dilutions.The sections were then incubated with fluorochromeconjugated secondary antibodies (Alexa 594 anti-mouse and Alexa 488 anti-rabbit; Invitrogen).
Immunohistochemical quantification
Slides were imaged at 4 × and 20 × magnification using an Eclipse 80i microscope (Nikon). Infiltrating CD3+, CD4+,CD8+and LAG3+cells were enumerated from 40× fields using inForm analysis software (Perkin-Elmer). The density of staining in infiltrating T cells was calculated by the number of positive cells/mm2as previously described27,28. The positively stained cells were quantified independently by two pathologists.
Mouse model
Six-to-eight-week-old C57/BL6 female mice were purchased from the Animal Experiments Center in Guangdong Province, China. For in vivo studies, mice were subcutaneously inoculated with 4×105MCA205 sarcoma cells. Approximately one week after tumor inoculation, mice were randomized into two treatment groups: isotype control[10 mg/kg, in vivo monoclonal antibody (mAb) mouse IgG1 isotype control, Clone: MOPC-21, Bioxcell] and anti-LAG-3 group (10 mg/kg; in vivo mAb LAG-3 antibody, Clone:C9B7W, Bioxcell). The mice were intraperitoneally injected every other day during the next 10 days as previously described29. Tumor volume was assessed using a micrometer caliper and calculated by the formula: (width2× length)/2.All mice were euthanized at the end of the study. The animal experiments followed protocols approved by the IRB at Sun Yat-sen University Cancer Center (Approval No.L102012017030R).
Statistical analysis
The differences between two groups were analyzed by an unpaired t-test. The correlation of LAG-3 mRNA expression with CD3/CD4/CD8 mRNA expression was evaluated by two-tailed Pearson’s statistics in GraphPad Prism 6.0 software. The data are presented as the mean ± SEM, and the statistical significance was determined as P < 0.05. The Kaplan-Meier method and log-rank test were used for the univariate analysis of overall survival (OS) and disease-free survival (DFS) by SPSS 17.0 software. Associations between clinical and pathological parameters with LAG-3 were evaluated using χ2or Fisher’s exact test.
Results
T cells infiltrating human STS express high levels of LAG-3 compared with those in blood
We performed flow cytometry on peripheral T cells from 12 tumor patients and 11 healthy donors. The example of gating strategy is shown in Figure 1A. Peripheral CD8+and CD4+T cells from STS patients expressed higher levels of LAG-3 than cells from healthy donors. LAG-3 was expressed at the highest levels in both peripheral CD4+and CD8+T cells(50.3% and 40.7%, respectively) from STS patients (Figure 1B and 1C). To confirm the protein expression of LAG-3 by tumor-infiltrating T cells in STS, we assessed surface LAG-3 expression in human tumor tissue of two STS patients.Intratumoral T cells displayed greater cell surface expression of LAG-3 compared with T cells from matched blood (Figure 1D).
LAG-3expression in STS is correlated with high pathological grades, advanced tumor stage, and poor prognosis
To further determine the expression of LAG-3 in STS, we performed immunohistochemical staining in human STS tissue samples from 163 patients who had undergone extensive resectable surgery. Importantly, the high expression of LAG-3 was significantly associated with a high pathological grade (low grade vs. high grade, P < 0.001), a more advanced tumor stage (I+II vs. III+IV, P = 0.016)(Table 1).
Representative images of the IHC staining of highly and poorly LAG-3-infiltrated lesions and isotype control are shown in Figure 2. LAG-3 expression was detected in some common subtypes of STS, including rhabdomyosarcoma,undifferentiated pleomorphic sarcoma, synovial sarcoma,and fibrosarcoma. Among the 163 patients, 22 were considered LAG-3high(subdivided by the optimal P-value cutoff); they displayed a shorter OS (P = 0.038), but did not reach significance for DFS (P = 0.336) (Figure 3A). In the undifferentiated pleomorphic sarcoma subgroup, the LAG-3highgroup was also associated with poor OS (P = 0.048) (data not shown).
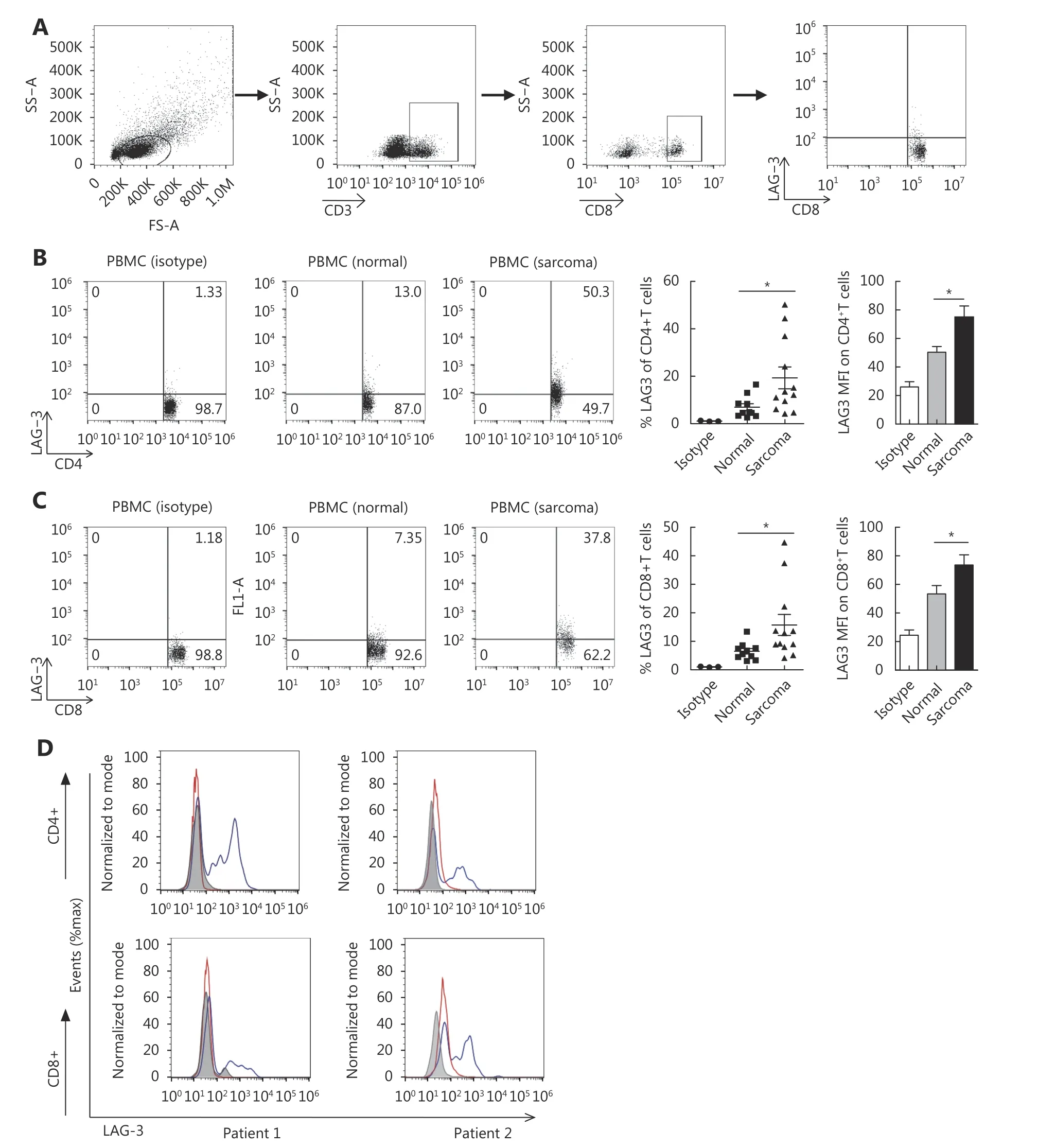
Figure 1 LAG-3 is expressed by human peripheral blood lymphocytes and tumor-infiltrating lymphocytes. (A) The example of gating strategy. (B) Representative FACS plots of LAG-3 expression by peripheral CD4+ T cells and CD8 + T cells from sarcoma patient blood and matched normal donor peripheral blood (left). Shown is the quantitation of LAG-3+ cells as a percentage of all CD4+ T cells (middle).Quantitation of LAG-3 MFI is also shown (right). *P < 0.05. Isotype control staining for CD4+ T cells is shown. (C) Representative FACS plots of LAG-3 expression by peripheral CD8+ T cells from sarcoma patient blood and matched normal donor peripheral blood (left). Shown is the quantitation of LAG-3+ cells as a percentage of all CD8+ T cells (middle). Quantitation of LAG-3 MFI is also shown (right). *P < 0.05. Isotype control staining for CD8+ T cells is shown. (D) Histograms of LAG-3 expression on CD4+ and CD8+ cells in blood (red) and tumor (blue) from a representative sample. Gray histograms represent isotype controls, which stain CD4+ and CD8+ T cells in the tumor.
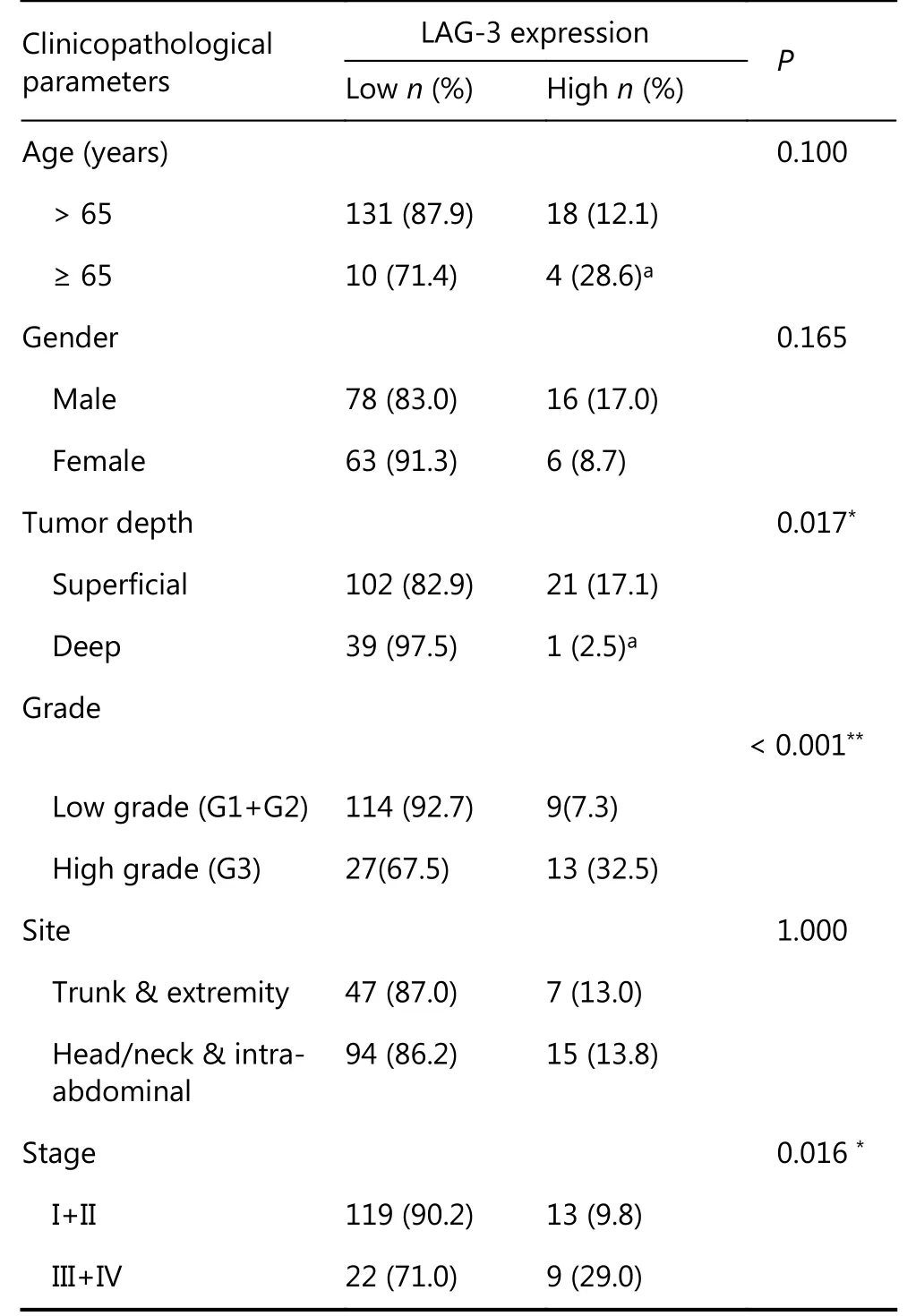
Table 1 Clinicopathologic variables and the expression status of LAG-3 in soft tissue sarcoma
Density of CD8+ T lymphocytes is correlated with poor prognosis in STS
The density of CD3+/CD4+/CD8+T cells in the primary tumors was heterogeneous in the cohort of 163 primary STS patients. Based on the optimal P-value (OPv) cutoff for DFS(OPv, 137 cells/mm2), the CD8high(n = 44, 27.0%) group displayed a significantly shorter DFS (P < 0.031) and OS (P <0.001) (Figure 3B). The OPv for CD3+T cell density was 114 cell/mm2, and the CD3high(n = 43, 26.3%) group showed a significantly shorter DFS (P = 0.006) and OS (P = 0.013)(Figure 3C). Furthermore, the CD4high(OPv, 156 cells/mm2)group had a significantly shorter DFS (P = 0.019) and OS(P = 0.01) (Figure 3D).
LAG-3 is highly correlated with CD8+ T-cell infiltration
We next analyzed the association between the expression of the immune inhibitory receptor LAG-3 and T-cell infiltration available from the TCGA public database of 265 STS cases.LAG-3 expression was highly correlated with CD3ε expression (Spearman’s rank correlation coefficient = 0.6391,P < 0.001; Figure 4A). LAG-3 expression in STS samples was highly correlated with CD8A and, to a lesser extent, with CD4 (r = 0.7128 and r = 0.4175, respectively). Furthermore,we observed intratumoral positive correlation between the cell density values for LAG-3 with CD3 (r = 0.6073, P <0.001), LAG-3 with CD4 (r = 0.4602, P < 0.001), and LAG3 with CD8 (r = 0.7034, P < 0.001) by immunohistochemistry(Figure 4B).
To confirm that LAG-3 expression was associated with CD8+T infiltrating cells in human STS, we performed immunofluorescence in human STS tissue samples and detected the expression and localization of LAG-3 predominantly in the membrane of tumor-infiltrating CD8+T lymphocytes (Figure 4C). These results suggested that tumor-infiltrating T cells, particularly CD8+T cells, may express high levels of LAG-3.
LAG-3 is expressed in murine TILs, and the blockade of LAG-3 decreases tumor growth
To confirm the upregulation of LAG-3 in the MCA205 mouse model, we quantified LAG-3 expression on CD4+and CD8+T cells by flow cytometry. One week after inoculation,when the MCA205 tumor had grown to 150-200 mm3in size,LAG-3 was expressed in approximately 40.5% of CD8+TILs and 43.4% CD4+TILs (Figure 5A and 5B). Given that LAG-3 was highly expressed by TILs, we investigated the efficacy of LAG-3 blockade in vivo using an immunocompetent MCA205 fibrosarcoma mouse model.
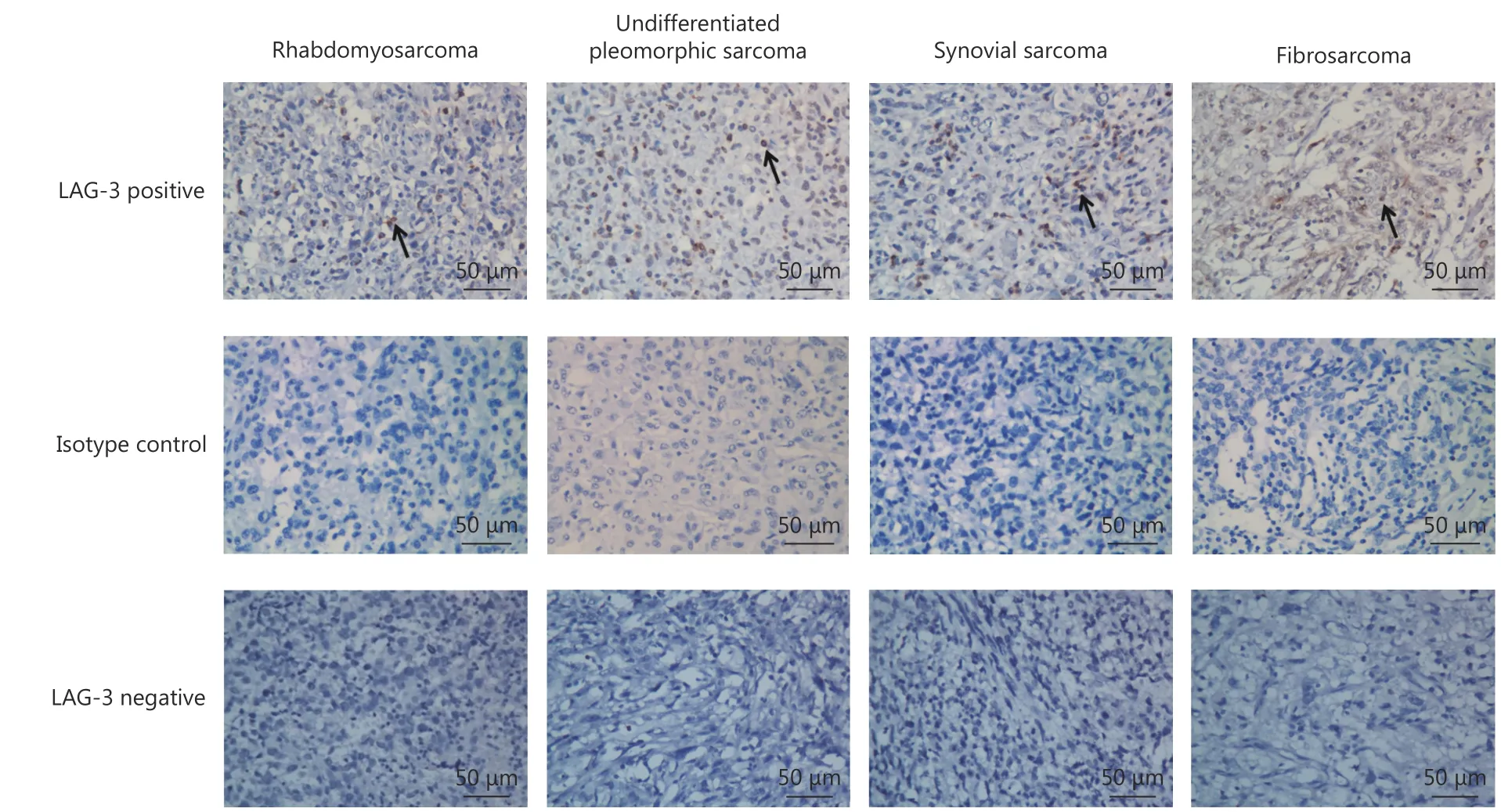
Figure 2 Representative immunohistochemical staining of LAG-3 in subtypes of human STS tissue samples. LAG-3 is expressed in the membrane of tumor infiltration lymphocytes in rhabdomyosarcoma, undifferentiated pleomorphic sarcoma, synovial sarcoma and fibrosarcoma. Arrows indicate LAG-3-positive lymphocytes. (IHC staining, 400 x).
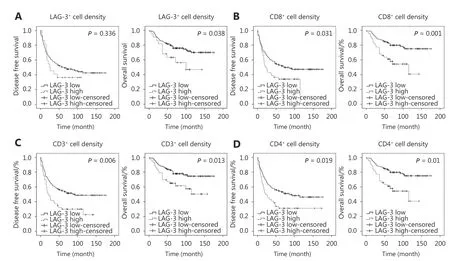
Figure 3 Expression of tumor-infiltrating lymphocytes and immune checkpoint LAG-3 correlates with unfavorable clinical outcome for patients with STS. OS and DFS according to the presence of a high density and low density of (A) LAG-3+ cells, (B) CD8+ cells, (C) CD3+ cells and (D) CD4+ cells. The P-values are also shown. The method to identify the high and low density has been described.
Mice were inoculated with MCA205 tumor cells, and blocking antibodies against LAG-324or control were intraperitoneally injected into mice every other day for the next 10 days (Figure 5C). In this model, treatment with anti-LAG-3 antibody significantly decreased the tumor burden(P < 0.001) (Figure 5D). We then assessed the functional consequences of LAG-3 inhibition on TILs. Tumorinfiltrating CD8+T cells and CD4+T cells from mice treated with anti-LAG-3 antibodies showed a significant increase in IFNγ compared with control mice (Figure 5E). These results were consistent with the idea that LAG-3 blockade can reactivate antitumor response of T cells.
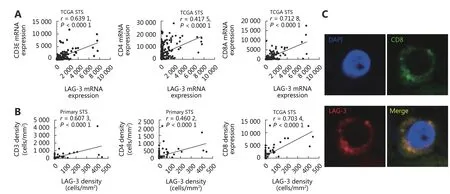
Figure 4 Expression of LAG-3 correlates with CD8+ T cell infiltration in primary STS. (A) Correlation of LAG-3 and CD3ε, CD4, CD8A RNA expression in primary STS from the TCGA database. (B) Dot-plot of CD3, CD4, CD8 (y-axis) cell density against LAG-3 cell density (x-axis).Pearson r value and the number of samples for correlation are displayed. (C) Immunofluorescence staining on paraffin-embedded STS showing colocalization of CD8 (green), LAG-3 (red) proteins in lymphocytes; nuclei are stained with DAPI.
Discussion
Cytotoxic CD8+T cells play a significant role in antitumor response in the tumor microenvironment30. The tumor microenvironment is highly immunosuppressive with a pronounced T-cell dysfunction; the exhaustion and dysfunction of effective T cells can be reversed by blocking the key immune checkpoint receptors31,32. In a majority of cancers, tumor infiltration by CD8+cytotoxic T cells is associated with good prognosis33. However, in diffuse large B-cell lymphoma34, Hodgkin lymphoma35and clear cell renal cell carcinoma36, high densities of CD8+T cells have been associated with poor clinical outcome. In STS, the prognostic role of CD4+T and CD8+T lymphocytes is still controversial.
Here, we performed immunohistochemical staining in human STS tissue samples from 163 patients and demonstrated that high densities of CD8+T cells correlated with a poor clinical outcome. LAG-3 was highly correlated with human tumor-infiltrating CD8+T cells in STS. In addition, LAG-3 expression in STS was also significantly associated with a poor clinical outcome. Meanwhile, higher LAG-3 expression in TILs in human STS tissues indicated higher pathological grades, larger tumor size. These findings suggest that the presence of LAG-3 in TILs facilitates tumor cell proliferation and induces tumor recurrence or metastasis.
Sorbye et al.37reported that CD4+cells represent a positive prognostic factor in STS based on univariate analyses, but they did not find a statistically significant correlation (P =0.15) between CD8+T cells and prognosis, which differs from our results. However, the different CD8+and CD4+cutoff values, different sarcoma subtypes, and different patient cohorts might hinder the comparability of their and our results.
LAG-3 was shown to improve tumor-specific CD8+T-cell activation, which did not require CD4+positive cells38. This supports a direct role for LAG-3 in regulating CD8+T cells.In addition, LAG-3 was reported to localize with CD8 molecule with 96% LAG-3+CD8+co-capped cells present39,which is consistent with our results.
Preclinical studies have suggested a role for immunotherapy in the therapeutic response of sarcomas, but clinical data remain limited. The results of another phase 2 clinical trial (NCT02406781) assessing pembrolizumab with metronomic cyclophosphamide in patients with metastatic STS showed that only 3 of 50 patients achieved progressionfree survival at 6 months40. Furthermore, a phase IB study(NCT01643278) examined the safety and efficacy of ipilimumab combined with dasatinib in eight patients with STS, but no objective responses were observed41. These results underscore the urgent need to investigate and explore novel immune checkpoint inhibitors in STS.
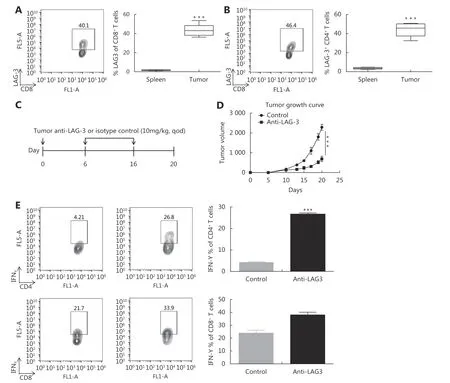
Figure 5 Anti-LAG-3 antibody retards tumor growth in MCA205 mouse model and regulates tumor-infiltrating CD8+ and CD4+T cell effector function (A) and (B) C57BL/6 mice were inoculated with MCA205 fibrosarcoma cells. Splenocytes and TILs were analyzed one week after inoculation when tumors had reached approximately 200 mm3 in size. Data are representative of more than three independent experiments (n=5-6). Shown is a representative FACS plot of LAG-3 expression by tumor-infiltrating CD8+ T cells and CD4+ T cells, with LAG-3+ cells boxed. Also shown is the quantitation of frequency of LAG-3+ T cells as a percentage of all T cells (P < 0.01). (C) Schematic image showing the immune therapeutic strategy for the MCA205 mouse model. C57BL/6 mice were randomly assigned into two groups and treated with isotype control and anti-mouse LAG-3 antibodies. (D) Tumors were measured with the electronic caliper and are presented as tumor volumes. Seven mice per group. ***P < 0.001. (E) Quantitation of IFNγ-producing tumor-infiltrating CD8+ and CD4+ T cells as percentages of total tumor-infiltrating CD8+ and CD4+ T cells respectively. *P < 0.05, **P < 0.01.
The present data show that the anti-LAG-3 antibody inhibited the growth of MCA205 fibrosarcoma in syngeneic C57BL/6 mice, supporting the clinical development of this novel immunotherapy in sarcoma. LAG-3 was highly expressed by both human and murine tumor-infiltrating CD8+T cells. In MCA205 tumor-bearing mice, antibody blockade of LAG-3 efficiently decreased tumor growth and enhanced CD8+T-cell effector function. Thus, blockade of LAG-3 was necessary to restore the potency of CD8+T cells within the immune-suppressive microenvironment in STS.This study provides a rationale for LAG-3 blockade for immune therapeutic approaches in future clinical trials.
There are some limitations in our study. LAG-3 is expressed on T cells, natural killer cells, plasmacytoid dendritic cells, and T regulatory cells42. Consistent with previous studies, we showed that LAG-3 expression identified a population of CD4+T cells and CD8+T cells. Moreover,LAG-3 blockade retarded tumor growth and enhanced antitumor response of CD8+T cells. However, we have not demonstrated LAG-3 expression and function in other lymphocyte subtypes, such as T regulatory cells and dendritic cells, which need to be explored in the future.
Conclusions
In summary, to the best of our knowledge, this is the first report on the poor prognostic impact associated with high density of LAG-3+cells in human STS. In addition,overexpression of LAG-3 is correlated with the clinicopathological parameters of STS. Our results suggest that LAG-3 is overexpressed on TILs, and anti-LAG-3 restrains tumor progression. The results from the evaluation of the expression level of LAG-3 in human STS provide a rationale for targeting LAG-3 to induce a significant antitumor immune response in STS.
Acknowledgements
This work was supported by grants from the National Key R& D Program of China (Grant No. 2017YFA0505600-04),and National Natural Science Foundation of China (Grant No. 81372887, 81572403, and 81772863).
Conflict of interest statement
No potential conflicts of interest are disclosed.
 Cancer Biology & Medicine2019年2期
Cancer Biology & Medicine2019年2期
- Cancer Biology & Medicine的其它文章
- Effects of palbociclib on oral squamous cell carcinoma and the role of PIK3CA in conferring resistance
- Genetic polymorphisms and gastric cancer risk: a comprehensive review synopsis from meta-analysis and genome-wide association studies
- Identification of anticancer drugs to radiosensitise BRAFwild-type and mutant colorectal cancer
- Hepatitis B virus X protein enhances hepatocarcinogenesis by depressing the targeting of NUSAP1 mRNA by miR-18b
- Cancer stem-like cells directly participate in vasculogenic mimicry channels in triple-negative breast cancer
- Factors associated with upstaging in patients preoperatively diagnosed with ductal carcinoma in situ by core needle biopsy