
不同海拔核桃叶中非结构性碳水化合物相关指标的变化及相关分析
2019-06-11 06:03张有福廉小芳陈春艳侯小改
植物资源与环境学报 2019年2期
张有福, 廉小芳, 陈春艳, 侯小改
(河南科技大学农学院, 河南 洛阳 471023)
近年来,碳循环及全球气候变化受到人们的广泛关注。 以木本植物为主的森林生态系统是全球最大的陆地碳库,约占陆地可利用碳量的45%[1]。 植物通常将光合作用固定的大部分碳水化合物用于自身生长,将另一部分碳用于防御或以可移动碳形式储存在体内[2]。 研究结果表明:植物的碳分配因环境改变而发生较大变化,并且,这种变化很大程度上依赖于非结构性碳水化合物(non-structural carbohydrate,NSC)的协调作用[3]。 非结构性碳水化合物是植物体内可移动碳的主要储存形式,其浓度反映了植物整体的碳供应状况[4];非结构性碳水化合物是植物碳供给(光合作用)和碳需求(呼吸与生长)代谢平衡的结果[2],能较好地反映植物生长和代谢对环境因子的响应和适应状况。 非结构性碳水化合物主要包括淀粉和可溶性糖,其中,淀粉只有在水解后才能参与植物的生理代谢和转运过程,主要起平衡和调节代谢的作用,是植物体内碳的长期储存形式;可溶性糖主要为蔗糖和果糖,可直接参与植物的各项生理代谢过程(如逆境渗透调节等)[5-6],其储存量反映了植物对逆境胁迫的适应性。 可见,非结构性碳水化合物组成变化在一定程度上能够反映植物响应环境变化的碳储存特征[7],探究植物非结构性碳水化合物的积累规律及其与环境变化的关系有利于揭示植物生长与碳储存的协调机制。
逆境条件下,植物体内非结构性碳水化合物的储存有利于弥补其光合碳源的降低和防御性代谢碳消耗的增加,从而保证植物正常的生理代谢[8],因此,增加组织中非结构性碳水化合物的储存量及降低生长消耗的非结构性碳水化合物的量是植物对逆境的适应性策略。 目前,研究者们关于海拔对植物生长和碳储存的影响存在较大争议。 Liang 等[9]认为,海拔升高可造成低温和生理性干旱,并缩短植物生长季节,使植物光合碳同化与碳消耗的关系失调,从而导致碳供应不足,最终使植物生长受限或面临生存危机。 然而,“生长抑制假说”却认为海拔升高不会导致碳供应不足,但可直接导致植物生长受限[10-14]。目前,缺乏海拔与落叶阔叶树种关系的系统研究[9],了解植物非结构性碳水化合物含量及其组成随海拔变化的规律有助于解析植物对高山环境的适应机制,为高山环境下植物种质资源的保护和利用提供参考。
核桃(JuglansregiaLinn.)又名胡桃,隶属于胡桃科(Juglandaceae)胡桃属(JuglansLinn.),为世界四大坚果树种之一[15],广泛分布于中国西部,具有显著的经济效益和生态效益。 中国是全世界核桃第一生产大国,西藏为主产区之一,所产核桃果仁含油率高、品质佳;并且,西藏的核桃分布区被认为是国内核桃的四大地理生态型之一[16]。 核桃在西藏地区栽培历史悠久,分布面积广,分布区海拔落差较大,主要分布在海拔2 400~3 500 m 的区域[17]。 这样的核桃海拔分布范围不但不可能出现在北方山地,而且远超同纬度东部地区和相邻川滇地区核桃的海拔分布范围。 另外,西藏核桃分布区的人为干扰较少,是研究植物与海拔和气候因子关系的理想场所。 鉴于此,作者对青藏高原东南部12 个不同海拔(2 500 ~3 868 m)样点的核桃叶中非结构性碳水化合物相关指标(包括非结构性碳水化合物、可溶性总糖、蔗糖、果糖和淀粉的含量以及可溶性总糖含量与淀粉含量的比值)变化进行了分析,并分别对各指标间及其与气候因子的关系进行了Pearson 相关性分析和逐步回归分析,以期探明非结构性碳水化合物积累和组成与海拔和气候因子的关系,为全面了解高海拔地区核桃的生态适应性和地理分布格局提供参考,并为高海拔地区核桃栽培区规划和核桃种质资源筛选提供基础资料。
1 材料和方法
1.1 材料
选取青藏高原东南部核桃分布区内12 个样点的成年核桃树进行采样,各样点的纬度、经度和海拔见表1。 在每个采样点核桃天然混交林边缘人为干扰较少且远离地表径流的区域,随机选择4 ~5 株树龄在50 a 以上、生长正常、无明显缺陷且病虫害较少的核桃植株,采集树冠外围阳面复叶上的第2 对小叶,每株至少采集10 枚小叶。 将单株小叶混匀,置于装有硅胶的自封袋中带回实验室,于60 ℃烘箱中烘干至恒质量,每株随机选取4 枚小叶,将同一样点所有样株的小叶混匀,去除叶脉后研磨成细粉,过100 目筛,保存在塑料自封袋中。
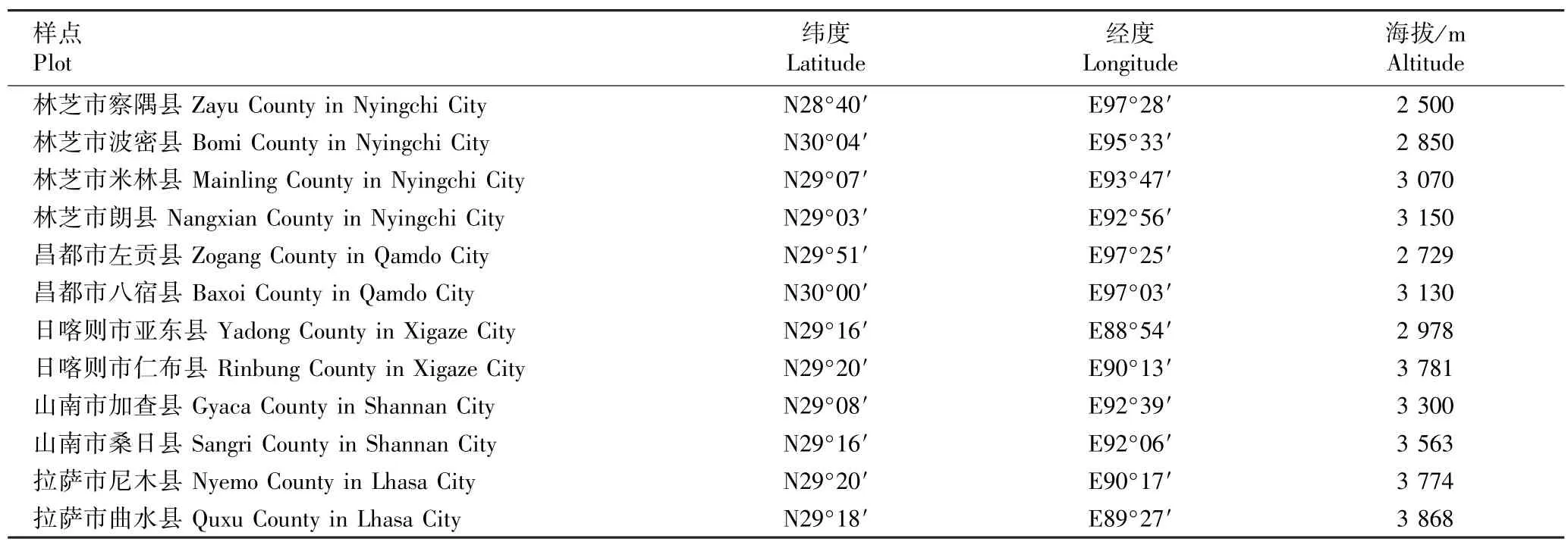
表1 供试核桃不同样点的具体位置Table 1 Specific locations of different plots of Juglans regia Linn. tested
从中国气象数据网(http:∥data. cma. cn/data/)获得各样点的气候因子(包括年平均气温、最热月平均气温、最冷月平均气温、月平均气温范围、年平均降水量、最冷月平均降水量和最热月平均降水量)。
1.2 方法
参照周永斌等[18]的方法分别测定可溶性总糖、蔗糖、果糖和淀粉的含量,并计算非结构性碳水化合物含量(即可溶性总糖含量和淀粉含量的总和)以及可溶性总糖含量与淀粉含量的比值。
1.3 数据分析
采用EXCEL 2007 软件制图,并采用SPSS 17.0统计分析软件进行Pearson 相关性分析和逐步回归分析。
2 结果和分析
2.1 不同海拔核桃叶中非结构性碳水化合物相关指标的变化
不同海拔核桃叶中非结构性碳水化合物相关指标的变化见图1,并以海拔为自变量(x)、非结构性碳水化合物及其主要组分的含量和比值为因变量(y)拟合线性回归方程。
2.1.1 非结构性碳水化合物含量的变化 研究结果表明:随海拔升高,核桃叶中非结构性碳水化合物含量呈上升趋势,获得的线性回归方程为y=0.095 7x-187.020 0(R=0.845 2,P=0.000 5),说明核桃叶中非结构性碳水化合物含量与海拔呈极显著正相关。并且,多个样点核桃叶中非结构性碳水化合物含量偏离回归直线,呈现一定的波动变化。
2.1.2 主要组分含量的变化 研究结果表明:随海拔升高,核桃叶中可溶性总糖、蔗糖和果糖的含量均呈上升趋势,获得的线性回归方程分别为y=0.089 7x-174.405 8(R=0.825 2,P=0.001 0)、y=0.030 2x-61.769 3(R=0.833 2,P=0.000 8)和y=0.021 3x-41.841 5(R=0.791 4,P=0.002 1),说明核桃叶中可溶性总糖、蔗糖和果糖的含量与海拔均呈极显著正相关。 并且,多个样点核桃叶中可溶性总糖、蔗糖和果糖的含量偏离回归直线,呈现一定的波动变化,且可溶性总糖和果糖含量的波动变化较蔗糖含量更明显。 另外,核桃叶中淀粉含量与海拔也呈极显著正相关,获得的线性回归方程为y=0.006 0x-12.618 0(R=0.883 2,P=0.000 1),并且,多个样点核桃叶中淀粉含量偏离回归直线,呈现一定的波动变化。
2.1.3 可溶性总糖含量与淀粉含量比值的变化 研究结果表明:随海拔升高,核桃叶中可溶性总糖含量与淀粉含量的比值呈抛物线型变化趋势,获得的回归方程为y=-2.091 4×10-5x2+0.136 6x-201.244 0(R=0.355 5,P=0.138 6),说明核桃叶中非结构性碳水化合物组成随海拔变化而发生改变,但相关性不显著,生长在海拔3 300 m 的核桃叶中可溶性总糖含量与淀粉含量的比值最大。

图1 不同海拔核桃叶中非结构性碳水化合物相关指标的变化Fig. 1 Variations of related indexes of non-structural carbohydrate in leaf of Juglans regia Linn. at different altitudes
2.2 Pearson 相关性分析
不同海拔核桃叶中非结构性碳水化合物相关指标的Pearson 相关性分析结果见表2。 由表2 可见:除了淀粉含量与非结构性碳水化合物含量和可溶性总糖含量呈显著(P<0.05)正相关外,非结构性碳水化合物含量、可溶性总糖含量、蔗糖含量、果糖含量和淀粉含量间均存在极显著(P<0.01)正相关,说明核桃叶中非结构性碳水化合物含量增加是可溶性总糖和淀粉共同积累的结果,可溶性总糖含量增加也是蔗糖和果糖共同积累的结果。
由表2 还可见:核桃叶中可溶性总糖含量与淀粉含量的比值与非结构性碳水化合物含量、可溶性总糖含量、蔗糖含量和果糖含量均呈正相关,且相关性不显著;但其与淀粉含量呈负相关,且相关性也不显著。值得注意的是,核桃叶中可溶性总糖含量与淀粉含量的比值与可溶性总糖含量相关系数的绝对值明显大于其与淀粉含量相关系数的绝对值,说明核桃叶中非结构性碳水化合物组成受可溶性总糖含量影响较大。
2.3 逐步回归分析
不同海拔核桃叶中非结构性碳水化合物相关指标与气候因子的逐步回归分析结果见表3。 由表3可见:核桃叶中非结构性碳水化合物相关指标与年平均气温、最热月平均气温、月平均气温范围和年平均降水量4 个气候因子有关,其中,非结构性碳水化合物含量、可溶性总糖含量、蔗糖含量、果糖含量和淀粉含量与这4 个气候因子显著(P<0.05)相关,而可溶性总糖含量与淀粉含量的比值与这4 个气候因子不显著相关;并且,各回归方程中年平均降水量的系数的绝对值明显小于年平均气温、最热月平均气温和月平均气温范围。 说明温度和降水因子对核桃叶中非结构性碳水化合物及其主要组分的含量和比值均有一定的影响,且温度因子的影响更大。
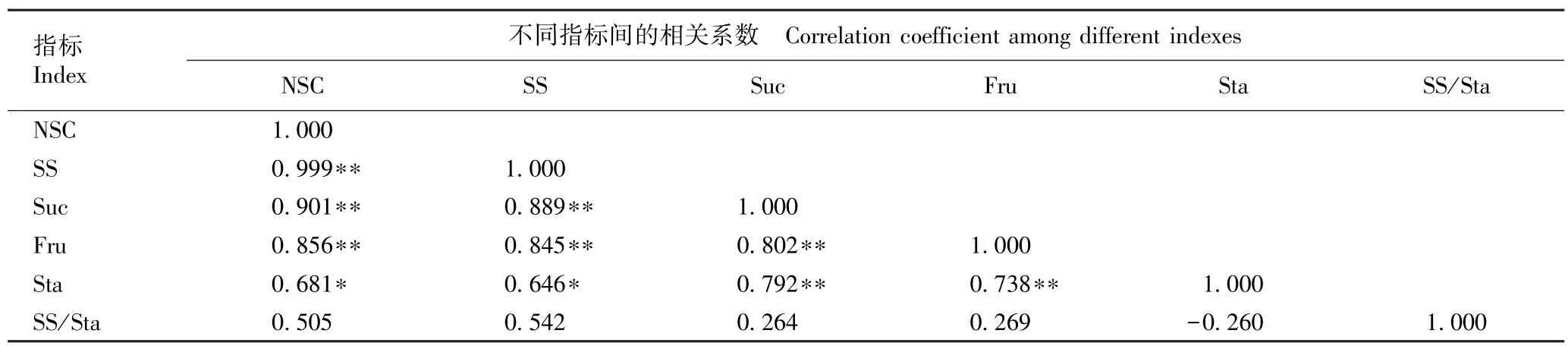
表2 核桃叶中非结构性碳水化合物相关指标的Pearson 相关性分析1)Table 2 Pearson correlation analysis on related indexes of non-structural carbohydrate in leaf of Juglans regia Linn. 1)

表3 核桃叶中非结构性碳水化合物相关指标与气候因子的逐步回归分析Table 3 Stepwise regression analysis on related indexes of non-structural carbohydrate in leaf of Juglans regia Linn. with climatic factors
3 讨论和结论
植物通过光合作用固定的碳是其生存的物质和能量基础,充足的碳供给有利于植物的生长发育及其在逆境中的生存[19]。 许多研究结果表明:高海拔区常绿和落叶树种的非结构性碳水化合物含量较高,并随海拔升高而增大[10-11,13,20],因此,有研究者认为非结构性碳水化合物积累量随海拔升高而增大是植物抵御逆境的进化反映[21]。 本研究中,核桃叶中非结构性碳水化合物、可溶性总糖、蔗糖、果糖和淀粉的含量均随海拔升高而增大,且非结构性碳水化合物含量与可溶性总糖含量、蔗糖含量和果糖含量呈极显著(P<0.01)正相关,与淀粉含量呈显著(P<0.05)正相关,说明随海拔升高核桃体内的可利用碳水化合物储存量增大,据此推测高海拔区域生长的核桃碳供应充足。 然而,本研究结果与美国东北部高山树种和热带高山树种的研究结果[22-23]却存在明显差异,这可能与不同植物种类对环境变化响应的阈值存在差异有关[24]。 值得注意的是,多个样点核桃叶中非结构性碳水化合物、可溶性总糖、蔗糖、果糖和淀粉的含量偏离回归直线,这可能是因为这些样点处于高山地区,由于山体的阻挡作用形成了不同的小气候环境,导致相同水平海拔样点的气候多样化,从而影响了核桃对光合作用固定的碳代谢和贮备。
相关研究结果[25]表明:海拔升高时,一方面,环境温度和CO2浓度降低,紫外线辐射强度增大,植物光合作用周期缩短,导致体内碳水化合物的净产量下降;另一方面,植物形成新细胞和新组织的速度逐渐下降甚至停止,并且,低温造成植物的呼吸作用下降,最终导致体内碳的消耗量下降。 有研究者认为植物生长限制导致其体内非结构性碳水化合物储存量增大[26];还有研究者认为植物体内非结构性碳水化合物储存量增大是植物的进化或可塑性反映,并不是生长限制的结果[27]。 本研究中,虽然核桃叶中可溶性总糖和淀粉的含量均随海拔升高而增大,但其可溶性总糖含量与淀粉含量的比值却随海拔升高呈抛物线型变化,这与核桃的胸径和相关生长指标随海拔升高的变化趋势一致(另文发表)。 在较低海拔区域,非结构性碳水化合物储存量增大可能是植物为了适应海拔变化而主动积累碳的结果[28],并且,非结构性碳水化合物含量和可溶性总糖含量与淀粉含量的比值增大可促进植物生长,保障防御性代谢消耗。 随着海拔进一步升高,高山地区的树木往往不能充分利用光合产物进行细胞生长[29],导致植株出现生长限制,造成体内碳的消耗量下降,从而使非结构性碳水化合物储存量增大变为可能[10]。 当植物出现生长限制时,可移动碳储量的增大更多依赖于淀粉[30],本研究结果也印证了这一结论,即在较高海拔区域,核桃叶中可溶性总糖含量与淀粉含量的比值随海拔升高而下降。 综上所述,核桃叶中非结构性碳水化合物含量在一定海拔范围内随海拔升高而增大的主要原因是植物适应环境的主动调节,另一个重要原因是在接近海拔上限,植株出现生长限制后的被动积累。
植物的地理分布受到环境中水热条件的共同作用[31]。 逐步回归分析结果表明:核桃叶中非结构性碳水化合物含量、可溶性总糖含量、蔗糖含量、果糖含量和淀粉含量与年平均气温、最热月平均气温、月平均气温范围和年平均降水量的相关性均达到显著水平,结合各气候因子的系数,认为温度因子是影响核桃叶中非结构性碳水化合物积累和组成的重要因子,同时,降水因子(年平均降水量)也对其积累和组成有一定影响。
综合上述研究结果,核桃叶中非结构性碳水化合物的积累随海拔升高而增大,且受到温度和降水因子的共同影响,并以温度因子影响为主。
致谢:西藏农牧学院马和平副教授在样品采集过程中提供了帮助,在此表示感谢!
猜你喜欢
文化创新比较研究(2020年7期)2021-01-13
China International Studies(2021年5期)2021-01-03
少儿科技(2021年8期)2021-01-02
矿产勘查(2020年6期)2020-12-25
铁道通信信号(2020年9期)2020-02-06
婚姻与家庭·性情读本(2019年11期)2019-11-18
保健与生活(2019年7期)2019-07-31
科技与创新(2016年22期)2017-03-30
家庭科学·新健康(2016年10期)2016-11-15
科技视界(2016年20期)2016-09-29
