
发酵豆粕替代鱼粉及添加包膜氨基酸、植酸酶对花鲈幼鱼生长性能及养殖水质的影响
2019-06-04 03:34:48杨艳玲曹俊明黄燕华孙育平王国霞莫文艳
饲料工业 2019年10期
■杨艳玲 陈 冰 彭 凯 曹俊明* 黄燕华 孙育平 王国霞 莫文艳
(1.广东省农业科学院动物科学研究所,广东广州510640;2.华南农业大学海洋学院,广东广州510642;3.农业部华南动物营养与饲料重点实验室,广东广州510640;4.广东省畜禽育种与营养研究重点实验室,广东广州510640)
随着集约化水产养殖规模的不断扩大,鱼粉资源日益短缺,继而导致鱼粉价格居高不下[1]。降低鱼粉用量或寻求鱼粉替代品是维持水产养殖可持续发展中亟待解决的问题。现阶段,水产养殖仍大量使用鱼粉,鱼粉中磷的含量较高,但大多数鱼类对鱼粉中磷的利用率低,导致磷不能被有效利用,而随粪便排入养殖水体,污染环境,恶化水质。花鲈(Lateolabrax japonicus),又名七星鲈,属鲈形目鮨科,广泛分布于我国沿海地带,是近年来兴起的名优水产养殖品种,其肉质细嫩、营养丰富,深受消费者喜爱。我国淡水花鲈的主要养殖区域为广东省,年产量达10万吨,在促进广东省渔业增效和水产养殖业发展中起支撑作用。随着集约化水产养殖的不断发展,花鲈的养殖密度也越来越高,造成水体中的氮、磷含量不断累积,水体富营养化,养殖水质恶化日益严峻,与国家倡导的健康生态养殖矛盾日益突出,因此开发低排放、低污染配合饲料是水产养殖包括花鲈健康养殖的关键。
与豆粕相比,发酵豆粕中抗营养因子含量很低[2],饲料风味和利用率较高,可促进动物肠胃对营养物质的吸收[3]。研究表明,发酵豆粕可减少饲料中鱼粉用量,降低水体污染。冯建等[4]研究表明,当发酵豆粕替代15%、30%和45%鱼粉时,不改变大黄鱼幼鱼的存活率、增重率和特定生长率,饲料中发酵豆粕添加量以30%较为适宜。包膜氨基酸[5-8]和植酸酶[9-12]具有促进动物生长,消除植物饲料中抗营养因子,改善生产性能,提高动物免疫力[13]等作用。Alam等[14]研究发现,饲料中添加包膜蛋氨酸和赖氨酸可显著提高日本对虾的生长性能。孟祥科等[15]研究表明,当植酸酶添加量大于1 000 U/kg时,红鳍东方鲀幼鱼的特定生长率和终末体质量显著升高,饵料系数显著降低。目前,关于发酵豆粕替代鱼粉添加包膜氨基酸、植酸酶对花鲈生长和养殖水体水质的影响上尚未见报道。本试验以花鲈幼鱼为试验动物,研究不同比例发酵豆粕替代鱼粉及外源添加氨基酸、植酸酶对花鲈生长性能及养殖水质的影响,以期为发酵豆粕、包膜氨基酸和植酸酶在花鲈饲料中的合理应用提供理论依据。
1 材料与方法
1.1 试验饲料
试验饲料配方如表1所示,其中鱼粉、发酵豆粕和大豆浓缩蛋白作为鱼类营养的主要蛋白来源,高精面粉作为主要糖类来源,鱼油、豆油及磷脂油作为主要脂肪来源。G0为基础饲料,在此基础上降低5%的鱼粉,添加12%发酵豆粕,记为G1。在G1的基础上,分别通过外源添加包膜氨基酸(包膜赖氨酸,其中L-Lys HCl含量为50%,L-Lys含量为40%;包膜蛋氨酸,DL-Met含量为40%)(包膜氨基酸由广州飞禧特水产科技有限公司提供)和植酸酶(2 500 U/g)(植酸酶由广东溢多利生物科技股份有限公司提供),记为G2和G3。在G0的基础上,降低10%的鱼粉,添加24%的发酵豆粕,记为G4。饲料原料经粉碎后过60目筛,维生素和矿物质预混料采取逐级扩大法混匀。用SLX-80型双螺杆挤压机将饲料制成粒径为2.0 mm的颗粒饲料,55 ℃烘干,自然冷却后放入密封袋中,置于-20 ℃冰箱中保存备用。
1.2 饲养管理
试验所用花鲈购自福建诏安县英港育苗场。驯养2周,期间投喂基础饲料。饲养试验在广东省农业科学院动物科学研究所水产研究室的室内循环水养殖系统中进行。每个处理组用一个单组循环系统(包含4个试验缸和1个过滤缸),每个处理组水体独立循环,试验缸容积为200 L(直径80 cm,高70 cm,水体容积150 L),进水速率为1.5 L/min。选取初始体重为(13.00±0.02)g的花鲈400尾,随机分为5组,每组4个重复,每个重复20尾,分别投喂5种试验饲料。每天投喂2次(9:00和18:00),饱食投喂,饲养42 d。试验期间为自然光照,且各组分别单独微量排污、清除粪便。

表1 试验饲料配方及营养水平(干基,%)
1.3 采样与分析
试验结束后,禁食24 h,每缸鱼称重并记录总数。每缸随机挑选8 尾鱼,用于测量体重和体长,计算肥满度。取其中4 尾保存于-20 ℃冰箱,用于全鱼常规营养成分分析。另外4尾解剖分离内脏团、肝脏并称量,用于计算肝体比和脏体比。
试验期间,于每天上午投喂饲料前测量水温和pH值。分别在试验的第1、7、14、21、28、35 d和42 d,测定水体溶氧量并采集每缸中部同一位置的水样并置于4 ℃冰箱保存,于48 h内测定氨氮、总氮、总磷及亚硝酸盐氮含量。
全鱼及饲料原料的水分、粗灰分、粗脂肪和粗蛋白质的测定均按照国标方法进行。水分的测定为105 ℃烘干至恒重(GB/T 6435—2014)。粗灰分的测定为箱式电阻炉550 ℃灼烧法(GB/T 6438—2007)。粗脂肪的测定采用索氏抽提法(GB/T 6433—2006)。粗蛋白的测定采用凯氏定氮法(GB/T 6432—1994)。钙的测定采用乙二胺四乙酸二钠络合滴定法(GB/T 6436—2002)。磷的测定采用分光光度法(GB/T 6437—2002)。
pH值使用笔式pH计(PHB-3,上海三信仪表厂)测定,校正后直接读取pH值。溶氧量(DO)使用FiveGo™便携式溶氧仪(METTLER TOLEDO F4,梅特勒-托利多国际贸易上海有限公司)测定,校准后直接读取溶氧值。总氮利用PLab-8型多功能比色测定仪(广州华宙环境科技发展有限公司)采用酚二磺酸分光光度法测定。氨氮利用PLab-8型多功能比色测定仪采用纳氏试剂比色法测定。总磷利用PLab-8型多功能比色测定仪采用钼酸铵分光光度法测定。亚硝态氮利用PLab-8型多功能比色测定仪采用亚硝酸盐氮分光光度法测定。
1.4 指标测定
成活率(SR,%)=(末尾数-初始尾数)/初始尾数×100
增重率(WGR,%)=(末均重-初均重)/初均重×100
特定生长率(SGR,%/d)=(ln 末均重-ln 初均重)/饲养天数×100
饲料系数(FC)=摄入饲料总量/(终末体重-初始体重)
肝体比(HSI,%)=肝脏重/体重×100
脏体比(VSI,%)=内脏重/体重×100
肥满度(CF,g/cm3)=体重/体长3×100
1.5 数据处理
试验结果采用“平均值±标准误”,采用SPSS 22.0软件对试验结果的差异显著性进行单因素方差分析(oneway ANOVA)和Duncan’s均值多重比较法对试验结果差异性进行分析处理,若不满足方差齐性则采用Dunnett-T3检验法进行多重比较。P<0.05表示差异显著。
2 结果与分析
2.1 生长性能(见表2)
由表2可知,G1、G2组和G3组中花鲈的末均重、增重率和特定生长率均显著高于G0组(P<0.05),G4组与G0组无显著差异(P>0.05)。与G1组相比,G2组中花鲈的末均重、增重率和特定生长率均显著增加(P<0.05)。G3 组中花鲈的末均重、增重率和特定生长率高于G1组,但差异不显著(P>0.05)。与G0组相比,G1、G2组和G3组饲料系数显著下降(P<0.05),但G4组与G0组无显著差异(P>0.05)。与G1组相比,G2组饲料系数显著下降(P<0.05)。G3组中饲料系数低于G1组,但差异不显著(P>0.05)。与G0组相比,G4组中花鲈的肝体比显著下降(P<0.05)。各组间成活率和肥满度无显著差异(P>0.05)。
2.2 全鱼体成分(见表3)

表2 花鲈生长性能

表3 花鲈全鱼体成分
由表3 可知,花鲈全鱼粗蛋白质、粗脂肪、灰分、水分、总磷和钙含量组间无显著差异(P>0.05)。
2.3 水质指标
2.3.1 pH值
由图1 可知,各组水体pH 值随养殖时间的延长呈下降趋势,第14 d 后下降趋势变缓。同一时间,各组间pH值差异不显著(P>0.05)。
2.3.2 溶氧量(DO)
由图2 可知,各组水体DO 随养殖时间的延长呈整体下降趋势,第21 d后DO基本不变。同一时间,各组间DO差异不显著(P>0.05)。
2.3.3 总氮(TN)

图3 水体总氮的变化
由图3 可知,水体TN 含量随养殖时间的延长呈逐渐上升趋势。同一取样时间,各组间TN 差异不显著(P>0.05)。
2.3.4 氨氮(NH4-N)

图4 水体氨氮含量的变化
由图4可见,水体NH4-N含量随养殖时间的延长呈逐渐上升趋势。第28 d之后,G0组NH4-N含量高于其它组(P<0.05),其中G4 组NH4-N 含量比G0 组低14.54%。第42 d,G4组NH4-N含量比G0组低20.60%。
2.3.5 亚硝酸盐氮(NO2-N)

图5 水体亚硝酸盐氮含量变化
由图5可见,水体NO2-N含量随养殖时间的延长出现波动变化。G0组NO2-N含量均高于其它组。第14 d之后,G2组和G3组NO2-N含量显著低于G0组(P<0.05)。
2.3.6 总磷(TP)
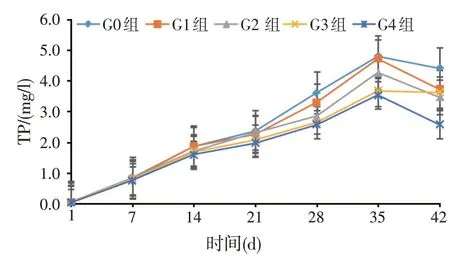
图6 水体总磷含量变化
由图6可知,水体TP含量随养殖时间的延长呈先上升后下降趋势。第21 d之后,G0组水体TP含量均显著高于G2、G3 组和G4 组(P<0.05)。其中第28 d,G3组比G0组降低26.92%。
3 讨论
3.1 发酵豆粕替代鱼粉及添加包膜氨基酸、植酸酶对花鲈生长性能的影响
与G0 组相比,G1 组花鲈的末均重、增重率和特定生长率显著升高,饲料系数显著降低,说明适量发酵豆粕替代鱼粉可提高花鲈幼鱼的生长性能。发酵豆粕是一种优质饲料蛋白源,其抗营养因子含量低,营养价值高[16],且含有丰富的小肽、氨基酸和消化酶等营养因子[17],可提高鱼体对饲料的消化吸收能力,进而提高其生长性能。Ding等[18]研究表明,在日本沼虾(Macrobrachium nipponense)饲料中用发酵豆粕替代25%的鱼粉时,饲料系数和增重率达到最佳并优于对照组。赵丽梅等[19]研究表明,用34%发酵豆粕替代鱼粉可提高卵形鲳鲹(Trachinotus ovatus)的生长性能。本试验结果表明,G4组花鲈的末均重、增重率和特定生长率与G0 组无显著差异,表明适量发酵豆粕在低鱼粉饲料中具有补偿生长作用。这与程成荣等[20]研究结果相似,当发酵豆粕替代40%鱼粉时对杂交罗非鱼(Oreoehromisniloticus ♀× O.aureus ♂)特定生长率、增重率和饲料效率无显著影响,当超过40%替代水平时,罗非鱼生长性能下降。
本试验结果表明,添加包膜氨基酸提高了花鲈的生长性能。冷向军等[21]研究发现,饲料中添加包膜氨基酸可显著提高异育银鲫(Carassius auratus gibelio)的增重率,降低饲料系数。原因可能是氨基酸通过包膜处理后,减少了其在水中的溶失率,从而使氨基酸在消化道中的吸收速度变缓,同时也促进了蛋白态的氨基酸与外源添加的氨基酸的同步吸收,进而促进花鲈的生长。相关研究表明[22-23],饲料中适量添加植酸酶可以提高水产动物的生长性能。植酸酶具有消除抗营养因子植酸的作用,促进植酸水解成肌醇与磷酸,从而提高鱼类对一些营养物质的消化吸收率,促进其生长。本试验结果表明,饲料中添加0.2%植酸酶不影响花鲈末均重、增重率和特定生长率。陈新宇等[24]研究发现,在基础饲料中添加500~2 000 U/kg 植酸酶,对凡纳滨对虾的饲料利用率和增重率均无显著影响。植酸酶对水产动物生长性能的影响可能与动物品种、大小、饲料营养和养殖环境相关。
本试验结果表明,发酵豆粕对鱼粉的替代比例对花鲈的肥满度没有产生显著影响,这与刘兴旺等[25]在大菱鲆(Scophthatmus maximus L.)饲料中利用大豆浓缩蛋白替代鱼粉的研究结果相似。
3.2 发酵豆粕替代鱼粉及添加包膜氨基酸、植酸酶对花鲈体成分的影响
本试验中,各组花鲈全鱼粗蛋白、粗脂肪、水分、灰分、钙和磷均无显著差异,说明发酵豆粕替代鱼粉及添加氨基酸、植酸酶不影响花鲈体成分。Luo等[26]在石斑鱼饲料中,用发酵豆粕替代不同比例鱼粉后,各试验组全鱼蛋白质、脂肪、水分、灰分含量无显著差异。王冠等[27]研究发现,添加包膜氨基酸不影响异育银鲫(Carassius auratus gibelio)肌肉中粗蛋白质、粗脂肪、水分和灰分含量。张璐等[28]研究表明,大黄鱼饲料中添加植酸酶对鱼体粗蛋白质、粗脂肪和水分无显著影响。
3.3 发酵豆粕替代鱼粉及添加包膜氨基酸、植酸酶对花鲈养殖水质的影响
在水产动物养殖过程中,良好的水体环境是保障动物健康生长的首要因素。集约化高密度养殖条件下的水体污染、水质下降等问题亟待解决,其中饲料是主要的影响因素之一。张加润[29]研究表明,鱼虾对鱼粉中氮、磷的利用率很低,大量不能被消化吸收的氮、磷便随之排入水体中,从而造成水体污染。姬红臣[30]研究发现,随着饲料中蛋白含量的增加,养殖水质的污染程度不断加重。因此,饲料营养组成直接影响到养殖水质和鱼类的健康生长。
通常,pH 值不适、溶氧低、氨氮及亚硝酸盐氮含量过高等均是水质污染严重的主要体现。有研究表明,高浓度的氨氮可使虾体致死[31]。本试验结果表明,随着发酵豆粕替代鱼粉比例的升高,水体中氨氮水平下降。原因可能是发酵豆粕替代鱼粉比例升高,减少了鱼粉的使用量,且发酵豆粕富含乳酸和酶类,可促进鱼类的消化吸收,减少水体中的残饵量,从而降低水体中的氨氮含量。同时使用发酵豆粕可提高氮的利用率,进而可减少水质净化剂的使用,降低水体中有害物质的残留,减少对养殖水体的污染。
过高的亚硝酸盐氮可使养殖动物的机体免疫力下降[32]。本试验中,添加包膜氨基酸和植酸酶使得养殖水体亚硝酸盐氮含量下降。原因可能是氨基酸和植酸酶具有增强饲料的适口性、消除植物性蛋白源中抗营养因子的作用,从而提高水产动物的消化能力,使得氮磷的排放量减少,进而降低对水体的污染,改善养殖水质。杨雨虹等[33]通过在斑点叉尾鮰饲料中添加植酸酶,发现随着植酸酶添加量的升高,水体氨氮和总磷不断下降。
本试验研究表明,发酵豆粕替代鱼粉及添加氨基酸和植酸酶可降低水体中总磷含量。原因可能是发酵豆粕经微生物发酵后,可将豆粕中含有的一些矿物质如植酸磷转化为鱼类可利用的矿物质,从而使饲料中总磷含量降低。又包膜氨基酸和植酸酶可提高鱼类对饲料中蛋白质和矿物质的利用率,减少粪便中磷的排泄量,从而降低磷对水体的污染。舒秋艳等[34]研究发现,添加植酸酶于青鱼(Mylopharyngodon piceus)饲料后,水体中氮磷含量上升趋势减缓。
4 结论
发酵豆粕替代鱼粉及添加包膜氨基酸和植酸酶可提高花鲈的生长性能,降低养殖水质中氨氮、亚硝酸盐氮和磷的排放,改善花鲈养殖水质。
猜你喜欢
当代水产(2022年8期)2022-09-20 06:46:06
当代水产(2022年6期)2022-06-29 01:13:12
国外畜牧学(猪与禽)(2021年6期)2022-01-11 05:59:40
国外畜牧学(猪与禽)(2021年4期)2021-08-31 01:53:40
当代水产(2019年6期)2019-07-25 07:52:08
当代水产(2019年1期)2019-05-16 02:41:50
中国畜禽种业(2018年1期)2018-01-18 23:05:48
渔业信息与战略(2016年2期)2016-12-02 09:54:55
河北渔业(2016年4期)2016-07-30 10:31:22
动物营养学报(2015年9期)2016-01-07 11:29:34
