
不同黄瓜砧木对低温弱光胁迫的响应及与ABA含量的关系
2019-06-04 10:58李福德毕焕改艾希珍
中国蔬菜 2019年5期
李福德 付 鑫 毕焕改艾希珍
(山东农业大学园艺科学与工程学院,作物生物学国家重点实验室,农业部黄淮地区园艺作物生物学与种质创制重点开放实验室,山东泰安 271018)
选用抗性砧木进行嫁接栽培可提高瓜类、茄果类等蔬菜作物对低温、高温、干旱和盐渍等非生物胁迫的抗性(Zhou et al.,2007;Han et al.,2013;Li et al.,2014;Xing et al.,2015),但不 同砧木对接穗的影响存在显著差异。于贤昌等(1998,1999)研究了不同耐冷性的南瓜砧木与黄瓜接穗对嫁接苗的影响,发现耐冷性均较强的接穗与砧木嫁接时才可获得耐冷性最强的嫁接组合,且嫁接苗耐冷性的增强与ABA、GA等激素的含量变化有关。李思思等(2018)研究指出,选用五叶香丝瓜作为砧木嫁接黄瓜较黑籽南瓜、日本南瓜等砧木能更好地提高黄瓜嫁接苗对根际高温的耐受性。田雪梅等(2012)选用不同耐盐性的南瓜砧木分别与新泰密刺黄瓜嫁接,嫁接苗对NaCl胁迫的响应不同,且嫁接苗耐盐性的高低与叶片Na+/K+有关。因此,探明不同砧木对非生物胁迫的响应机制有利于生产上不同抗性砧木的筛选。
黄瓜(Cucumis sativusL.)是北方日光温室的主栽蔬菜,在栽培过程中经常遭遇非生物胁迫逆境,尤其是秋冬茬日光温室黄瓜栽培中低温弱光环境最为常见,造成黄瓜产量和品质大幅下降(艾希珍 等,2004)。嫁接因其经济、简单、有效成为增强设施黄瓜耐冷性的主要手段,黑籽南瓜耐冷性较强,被广泛用于黄瓜嫁接生产,但因其不具备脱蜡粉能力,常导致嫁接苗果实外观和品质下降(Lee et al.,1999;李红丽 等,2006;刘青 等,2012)。因此,筛选耐冷性强,且对接穗品质影响较小的白(黄)籽南瓜砧木对日光温室黄瓜嫁接生产具有重要的意义。本试验以不同白籽南瓜为试材,以常规砧木黑籽南瓜为对照,通过分析低温胁迫下不同砧木耐冷指标和ABA含量的变化,筛选出耐冷性强的白籽南瓜砧木,并探明其高耐冷性与ABA的关系,以期为优化黄瓜嫁接栽培提供优良砧木。
1 材料与方法
1.1 试验设计
试验于2017年在山东农业大学南校区试验站进行。以津优35号黄瓜和常规砧木黑籽南瓜为对照,以金妈妈519、金妈妈619、格顿联丰、格顿西光、格顿金凯、博强102、博强103、博强105、博强4号等白籽南瓜砧木为试材(表1),4月5日播种于日光温室8cm×8cm营养钵中。每品种播种100株,选取80株正常生长幼苗进行试验。温室环境为:光量子通量密度(PFD)日均值为600μmol·m-2·s-1,昼 /夜温度均值为25℃/18℃。待幼苗长至一叶一心时移入日本山崎黄瓜专用营养液(含0.5mmol·L-1NH4H2PO4,2.0mmol·L-1Ca(NO3)2·4H2O,3.2mmol·L-1KNO3和1.0mmol·L-1MgSO4·7H2O,微量元素为全浓度,电导率2.2~2.5mS·cm-1,pH值6.8~7.0)进行水培,至三叶一心时(5月3日)转移至宁波产光照培养箱中进行低温弱光处理,条件如下:昼/夜温度8℃/5℃,光照100μmol·m-2·s-1,光周期11h/13h(昼/夜)。

表1 试验材料及来源
1.2 测定项目
1.2.1 冷害指数 冷害指数的调查参照于贤昌等(1998)的方法。幼苗冷害症状分级标准:0级,叶片正常,无受害症状;1级,叶片稍皱缩,第1叶或第2叶叶缘发黄或略失水,第3叶和心叶无受害症状;2级,叶片皱缩,第1叶和第2叶叶缘严重失水,第3叶叶缘发黄或略失水,心叶无明显受害症状;3 级,第1叶和第2叶中部出现脱水斑,第3叶叶缘严重失水,心叶轻微失水;4 级,第1叶和第2叶中部脱水斑连接成片,叶片萎蔫,第3叶中部始现脱水斑,心叶失水较明显,但常温下心叶尚能恢复;5 级,所有叶片严重失水萎蔫,幼苗在常温下不能恢复。
于低温弱光胁迫3、5、7 d后不同品种幼苗分别选取10株调查冷害症状,3次重复,按上述分级方法判定冷害级数,计算冷害指数。
冷害指数 =(1×S1+2×S2+3×S3+4×S4+5×S5+0×S0)/(低温胁迫总株数×5)
式中S为每一冷害级的株数。
1.2.2 电解质渗漏率与丙二醛含量 分别于低温胁迫0、7 d时,取不同品种幼苗3株,3次重复,选择由上向下数第2片叶,打取直径0.8 cm的叶圆片,放入具塞试管中,用美国产 ORION TDS 型电导率仪测定叶片的电导率,计算电解质渗漏率(EL)。
EL=(E1-E0)/(E2-E0)×100%
式中,E1:初电导率,E2:沸水30 min后电导率,E0:去离子水电导率。
低温胁迫7 d后取不同品种幼苗3株,3次重复,将由上向下数第2片叶剪碎称取0.5 g,采用硫代巴比妥酸(TBA)显色法测定丙二醛(MDA)含量(李合生,2000)。
1.2.3 根系活力 低温胁迫7 d后,取不同品种幼苗3株,3次重复,将根系均匀剪成1 cm的小段并称取0.2 g至试管中,采用TTC法测定根系活力(李合生,2000)。
1.2.4 气体交换参数 采用Ciras-3型光合仪(PPSystems公司,美国)测定胁迫0 d和3 d时不同砧木由上向下数第2片叶的净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr),测定时PFD、CO2浓度和叶温分别由仪器的可调光源、内置式 CO2供气系统和温度监控装置调控。每个品种测定幼苗6株,3次重复。
1.2.5 幼苗生长量 在低温弱光胁迫0 d和7 d时,分别选取不同品种幼苗10株,3次重复。用直尺测量株高及第1、2、3片叶叶柄到叶尖的长度,用游标卡尺测量茎粗,并按照龚建华和向军(2001)的方法计算叶面积,将植株烘干至恒重后用称重法称量干物质量。
日增量=(低温胁迫7 d的生长量指标-未经低温处理的生长量指标)/7
1.2.6 隶属函数值 参照宋洪元等(1998)模糊数学中隶属函数的方法,计算参试材料耐冷性指标的隶属函数值,并累加比较不同品种的耐冷性。
1.2.7 ABA含量 低温胁迫0 d和1 d分别剪取不同品种幼苗由上向下数第2片叶和根系,用锡箔纸包好置于液氮中速冻,参照Andygene公司生产的酶联免疫试剂盒(货号:AD0021Pl)说明书测定叶片和根系中的ABA含量,每品种3次重复。
1.3 数据处理
试验数据采用Microsoft Excel软件进行处理,运用DPS软件进行单因素方差分析,并运用Duncan检验法对显著性差异(P<0.05)进行多重比较,利用SigmaPlot软件作图。
2 结果与分析
2.1 低温弱光胁迫下不同砧木幼苗的冷害指数
从表2可以看出,随着低温弱光胁迫时间的延长,不同品种幼苗的冷害指数均逐渐增加,冷害加剧。津优35号的冷害指数除在胁迫3 d时显著低于格顿金凯,其余时间均为最高,受冷害影响最大。常规砧木黑籽南瓜的冷害指数一直较低,低温耐受性表现较好。
从表2还可以看出,整个胁迫过程中,博强105和金妈妈519白籽南瓜的低温耐受性表现亦较好,二者的冷害指数与黑籽南瓜无显著差异;其余砧木幼苗的低温耐受性在胁迫5~7 d时均强于津优35号,但低于黑籽南瓜。结合图1不同砧木幼苗的表现亦可知,低温弱光胁迫7 d后,津优35号幼苗的受害程度最大,叶片严重发黄萎蔫,而其余砧木表现则均优于津优35号,受害程度较轻,其中冷害指数较低的黑籽南瓜、博强105和金妈妈519表现较好。

表2 低温弱光胁迫下不同砧木幼苗的冷害指数

图1 低温弱光胁迫7 d后不同砧木幼苗的冷害症状

图2 低温弱光对不同砧木幼苗电解质渗漏率(A)和丙二醛含量(B)的影响
2.2 低温弱光对不同砧木幼苗电解质渗漏率(EL)和丙二醛(MDA)含量的影响
图2-A表明,低温弱光胁迫7 d后,不同品种幼苗的电解质渗漏率均出现不同程度的上升,其中格顿金凯的电解质渗漏率与0 d相比增加了53.1个百分点,增加幅度最大;津优35号增加了33.1个百分点,而黑籽南瓜仅增加了11.4个百分点,博强105、金妈妈519分别增加了12.9和12.1个百分点,增幅较小。低温弱光胁迫7 d后不同幼苗的电解质渗漏率从小到大依次为:黑籽南瓜<金妈妈519<博强105<博强103<博强4号<金妈妈619<格顿联丰<格顿西光<津优35号<博强102<格顿金凯,说明砧木黑籽南瓜、博强105和金妈妈519幼苗受到的低温伤害较轻。
低温弱光胁迫后不同品种幼苗叶片中MDA含量的变化与电解质渗漏率的变化趋势相似,即胁迫7 d后MDA含量均增加,津优35号的上升幅度较大,黑籽南瓜、博强105和金妈妈519的增加幅度较小(图2-B)。
2.3 低温弱光对不同砧木幼苗根系活力的影响
由图3可以看出,不同品种在常温条件下的根系活力差异较大。低温胁迫前黑籽南瓜、博强105、金妈妈519根系活力较高,低温胁迫7 d后除格顿联丰外其余品种根系活力均有不同程度的升高,其中金妈妈519根系活力最高,黑籽南瓜、博强4号次之。

图3 低温弱光对不同砧木幼苗根系活力的影响

图4 低温弱光对不同砧木幼苗气体交换参数的影响
2.4 低温弱光对不同砧木幼苗气体交换参数的影响
因监测的Pn在胁迫3 d时开始出现负值,因此图4只显示不同品种幼苗低温弱光处理0 d和3 d的光合气体交换参数。从图4-A可以看出,胁迫前所有砧木幼苗叶片的Pn均明显高于津优35号,以博强105和金妈妈519最高,甚至显著高于黑籽南瓜;胁迫处理3 d后,砧木Pn明显降低,但仍表现为所有南瓜砧木幼苗的Pn值均高于津优35号,其中金妈妈519和博强105的表现最好,亦显著高于黑籽南瓜。
低温胁迫前,各品种的Tr(图4-B)、Ci(图4-C)和Gs(图4-D)多差异不显著,而胁迫3 d后,不同品种幼苗的Tr和Gs均明显降低而Ci除黑籽南瓜、博强4号、金妈妈519出现降低,其他品种均出现不同程度上升。与津优35号相比,南瓜砧木幼苗的Gs与Tr均较高或与津优35号差异不显著,而Ci值均低于津优35号。
2.5 低温弱光对不同砧木幼苗生长量的影响
从表3可以看出,黑籽南瓜、金妈妈519及博强105的株高、茎粗、叶面积和干物质量的日增量均表现较好,其中黑籽南瓜表现最好,株高、干物质量的日增量均显著高于其他品种,茎粗、叶面积的日增量与博强105、金妈妈519差异不显著。津优35号和博强102、格顿金凯表现较差,因受冷害较重,叶缘严重失水卷曲,叶面积的日增量甚至出现负值。
2.6 低温弱光胁迫下不同砧木幼苗耐冷性综合评价
如表4所示,各砧木幼苗的隶属函数值均高于津优35号,其中金妈妈519最高,其次为博强105,且二者均高于黑籽南瓜;博强102和格顿金凯的隶属函数值较低。说明白籽南瓜金妈妈519和博强105具有高于常规耐冷砧木黑籽南瓜的低温耐受性,博强102和格顿金凯的低温耐受性较弱。
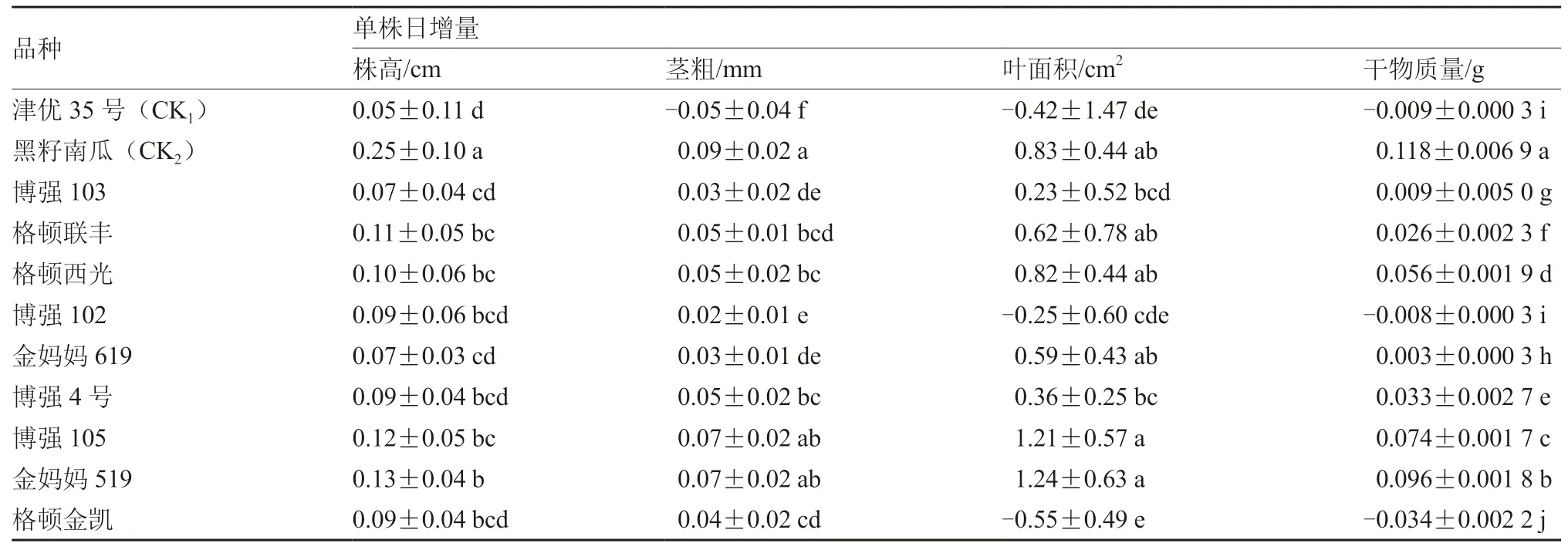
表3 低温弱光对不同砧木幼苗生长量的影响

表4 低温弱光胁迫下不同砧木幼苗耐冷性指标综合评价
2.7 低温弱光对不同砧木幼苗根系和叶片中ABA含量的影响
从图5可以看出,幼苗经过1 d的低温胁迫后根系和叶片ABA含量均有所增加,其中金妈妈519幼苗根系ABA含量最高,黑籽南瓜、博强105、博强4号幼苗根系的ABA含量也显著高于其他品种;金妈妈519幼苗叶片中的ABA含量在常温下虽然最低,但胁迫1 d后却是最高的,其ABA增加量明显高于其他砧木幼苗。对不同砧木根系和叶片中ABA增加量与砧木幼苗隶属函数值进行相关性分析,得出相关系数r=0.765 8,说明不同砧木耐冷性的强弱与根系中及根系+叶片中ABA的增加量呈显著正相关,但与叶片中ABA含量的增加量无显著正相关性(图6)。

图5 低温弱光对不同砧木幼苗ABA含量的影响
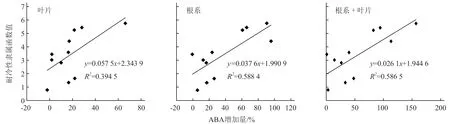
图6 ABA增加量与耐冷性隶属函数的相关性
3 结论与讨论
本试验中,随着低温弱光胁迫时间的延长,幼苗的冷害指数逐渐增加,受害程度加剧。电解质渗漏率和MDA 含量均可反映细胞膜脂过氧化程度及对非生物胁迫耐性的强弱(Chaoui et al.,1997;李天来和李益清,2008;马乐元 等,2017)。本试验中,低温弱光胁迫7 d 后,所有幼苗的电解质渗漏率和MDA 含量均出现不同程度的上升,常规砧木黑籽南瓜的冷害指数、电解质渗漏率和MDA含量最小,且与金妈妈519 和博强105 无显著差异;说明低温弱光胁迫下黑籽南瓜、博强105、金妈妈519 的受伤害程度较低,对低温弱光的耐受性强于其他砧木材料。
嫁接增强接穗抗性的主要机理是砧木具有发达的根系。韩敏等(2018)的研究也证实了砧木根系对嫁接苗耐冷性高低的贡献远大于接穗。本试验中低温胁迫0 d 时除博强102 砧木的根系活力与津优35号的根系活力无显著差异外,其余材料的根系活力均显著高于津优35号。前人研究指出,低温胁迫会导致根系活力下降(周艳虹 等,2003;刘玉凤 等,2017),但在本试验中,低温弱光胁迫7 d后根系活力多出现不同程度的增加,这可能是因为参试砧木幼苗是在水培条件下进行低温处理的,这与王佩(2014)的研究结果一致,即水培条件会增强植物的根系活力,其中以金妈妈519的根系活力最高。
Schupp和Ferree(1990)研究指出,光合速率的变化与根系存在一定的联系。本试验中,低温弱光胁迫3 d后不同品种幼苗的Pn、Tr和Gs明显下降,津优35号的Pn甚至出现负值,但根系活力最高的金妈妈519的Pn亦是最高。这可能是因为根系活力较高有助于吸收更多的矿质元素,促进叶片中色素的合成(王元征 等,2011),有利于Pn增加,积累更多的光合产物,从而使得低温弱光下金妈妈519的株高、茎粗、叶面积和干物质量的日增量均表现较好。
同时,从表4亦可以看出所有砧木的隶属函数值均高于津优35号,其中金妈妈519最高,其次为博强105,且二者均高于常规耐冷砧木黑籽南瓜。于贤昌等(1999)研究指出,嫁接苗耐冷性的增加与ABA、IAA、GA等激素的变化有关;同样,Li 等(2014)发现,以丝瓜为砧木的嫁接黄瓜高温抗性的增加主要是因为砧木根系中ABA含量的积累。本试验结果显示,低温弱光胁迫后砧木材料叶片和根系中ABA 含量均有所增加,且不同砧木耐冷性的高低与根系中及根系+叶片中ABA的增加量呈显著正相关,说明砧木材料耐冷性的高低确与ABA含量的变化有关,可将其作为日后筛选耐冷砧木的一个重要参考指标,但其机理有待于进一步研究。
综上分析可知,金妈妈519和博强105可选作新的黄瓜耐冷砧木,尤其是低温弱光胁迫下金妈妈519仍具有很高的根系活力,更适合在黄瓜嫁接生产上推广应用。
猜你喜欢
江西通信科技(2022年2期)2022-08-08
福建农林大学学报(自然科学版)(2021年5期)2021-10-08
落叶果树(2021年6期)2021-02-12
河北果树(2020年2期)2020-05-25
阅读与作文(小学高年级版)(2019年5期)2019-05-27
新农业(2017年3期)2017-05-17
中国果业信息(2017年8期)2017-01-13
福建农业科技(2016年10期)2016-03-07
摄影世界(2014年7期)2014-09-10
