
菜豆A18-160Co-γ射线诱变和突变体库的构建
2019-06-04 10:58陈其福斯琴图雅赵弘韬刘大军冯国军
中国蔬菜 2019年5期
陈其福 刘 畅 斯琴图雅 赵弘韬 刘大军 冯国军*
(1黑龙江大学农作物研究院,黑龙江哈尔滨 150080;2黑龙江省科学院技术物理研究所,黑龙江哈尔滨 150010)
诱变对基因组成的损伤和改变,涉及大量潜在靶向性状基因的突变,是生物学中鉴定基因功能的基本方法。通过物理、化学或生物手段诱导作物发生突变,增加可利用的基因组变异,是获得所需农艺性状的有效和低成本的手段(Talukdar,2014)。近年来,已经在菜豆(Porch et al.,2009;郭宁 等,2018)、大豆(Hwang et al.,2015)、绿豆(Singh et al.,2000;Wani et al.,2017)、 豌 豆(Sharma et al.,2015)、扁豆(Ali et al.,2010)、鹰嘴豆(Barshile et al.,2009)等主要豆科作物中开发稳定的突变种群和构建饱和连锁图谱方面取得了很大进展,极大地促进了豆科作物的改良和功能基因组研究。突变体是研究植物遗传、育种、基因定位和基因鉴定的理想材料,植物的光合作用(Chua & Bennoun,1975;Mullineaux,1994)及一些代谢途径(Zhang et al.,2010;Beckers et al.,2016)的基础理论研究大都是利用突变材料作为试材而展开的。
菜豆(Phaseolus vulgarisL.)是一种重要的蔬菜作物,因其味道鲜美、营养丰富而深受广大人民的喜爱。菜豆存在丰富的植物学和农业生物学性状的变异。菜豆是自花授粉作物,花为蝶形两性完全花,具有闭花自花授粉的习性。由于花器官的结构特殊,菜豆杂交授粉难度大,因此利用诱变获得生物遗传变异对于菜豆新品种选育尤为重要。在IAEA突变品种数据库(https://mvd.iaea.org/)记录的突变品种中,有多个利用人工诱变获得的突变材料选育的抗叶斑病和金色花叶病毒病的菜豆品种。本试验通过不同剂量的60Co-γ射线辐照菜豆干种子,筛选半致死剂量,为辐射育种技术研究提供参考。构建的突变体库包含多种突变类型,可以为菜豆品种改良和基础研究提供新的材料。
1 材料与方法
1.1 供试材料
供试材料为黑龙江大学农作物研究院提供的矮生菜豆品种A18-1的干种子。该品种的生长习性为有限生长类型,早熟,植株茎表皮和叶脉为浅绿色,花为粉白色,荚为圆棍形、略弯曲,平均荚长15 cm,绿色的幼荚表皮在成熟过程中逐渐转变为黄色,熟期为50~55 d。
1.2 半致死剂量筛选
选择均匀饱满的菜豆品种A18-1的干种子6份,每份100粒,用辐射强度为0.1 Gy·min-1的60Co-γ射线处理,处理时间分别为0、500、1 000、1 500、2 000、2 500 min,辐射剂量分别为 0、50、100、150、200、250 Gy。将辐射处理后的种子用穴盘单粒播种,发芽后逐日记录种子发芽数、出苗数(以初生真叶展开为标准)、苗成活数,直至不再有新的种子发芽,并记录最终成苗数。
发芽率=最终发芽数/播种总粒数
相对发芽率=(处理后种子的实际发芽率/对照种子发芽率)×100%
同理计算不同处理的相对出苗率。用Microsoft Excel软件分析辐射剂量与相对出苗率的线性关系,以辐射剂量为横坐标,相对出苗率为纵坐标建立辐射剂量与相对出苗率的线性回归方程。根据回归方程,计算相对出苗率为50%时的辐射剂量为半致死剂量。
1.3 诱变处理
选择矮生菜豆品种A18-1的种子2 500粒,在黑龙江省科学院技术物理研究所进行60Co-γ辐射处理,辐射强度为0.1 Gy·min-1,辐射剂量为126 Gy。辐射处理后的M1种子于2017年11月在海南播种,最终收获种子的单株共826株,占处理种子总数的33%。
1.4 M2突变体库表型筛选
M2于2018年5月21日在黑龙江大学呼兰实验基地播种。共收集786份M2种子,其中20粒种子以上的材料单行单粒播种20株,其余全部单粒播种,总共播种15 600株M2单株。从出苗后开始观察植株表型的变化,包括叶、茎、熟期、育性、荚和株型等性状,并对每一株突变单株进行编号和表型描述,便于后期采收整理和记录。荚完全成熟后,单独采收每一株突变株的种子;对于无法获得种子的突变株,采收所在株系的其余单株的种子。
2 结果与分析
2.1 半致死剂量筛选
对发芽率的统计结果表明,不同辐射剂量处理的发芽率虽然都低于对照,但不同处理间的相对发芽率变化不大(表1)。辐射处理对M1种子的致死效应在种子发育过程中逐渐显现。低剂量处理的发芽率与出苗率相当,随着辐射剂量的增加,出苗率明显低于发芽率,因此计算半致死剂量时以出苗率为计量标准。对出苗率的统计结果显示,相对出苗率随着辐射剂量增加而明显降低(表1)。根据辐射剂量与相对出苗率的线性回归方程y=-0.003x+0.916(图1),相对出苗率为50%时的辐射剂量为126 Gy,即为半致死剂量。
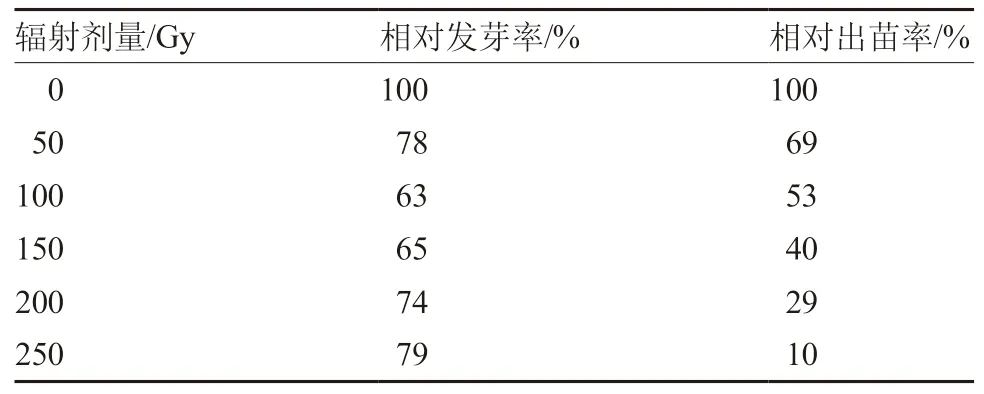
表1 60Co-γ射线处理干种子的相对发芽率和相对出苗率
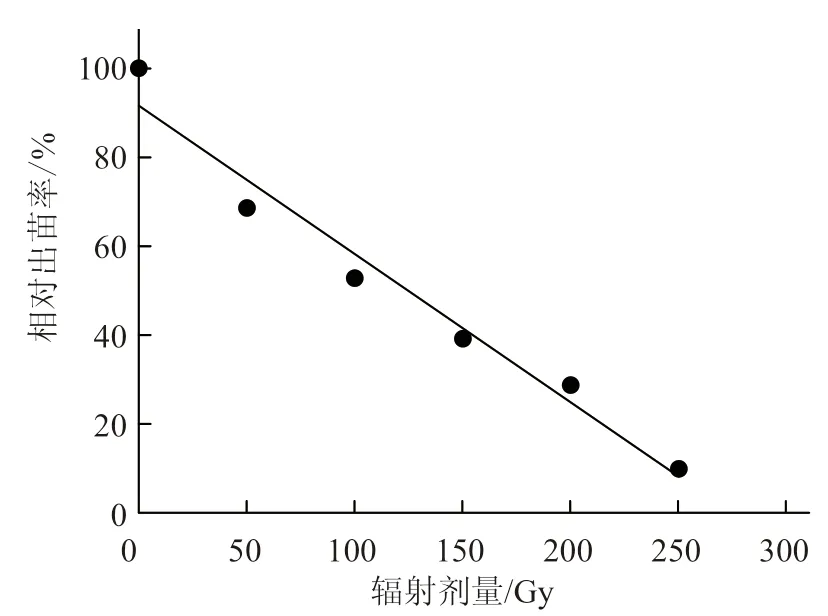
图1 辐射剂量与相对出苗率的线性关系
2.2 M2表型观察
对M2的田间表型进行鉴定,共发现279株突变株(表2),将突变体按突变性状分为生长习性和株型突变、熟期突变、黄化突变、茎突变、叶突变、荚突变、花和育性突变7种类型,总突变率为1.83%。因为花和育性突变直接相关,故将两类突变一起统计。株型性状较为复杂,由多个相关性状组成,在统计的突变性状中突变率最高。
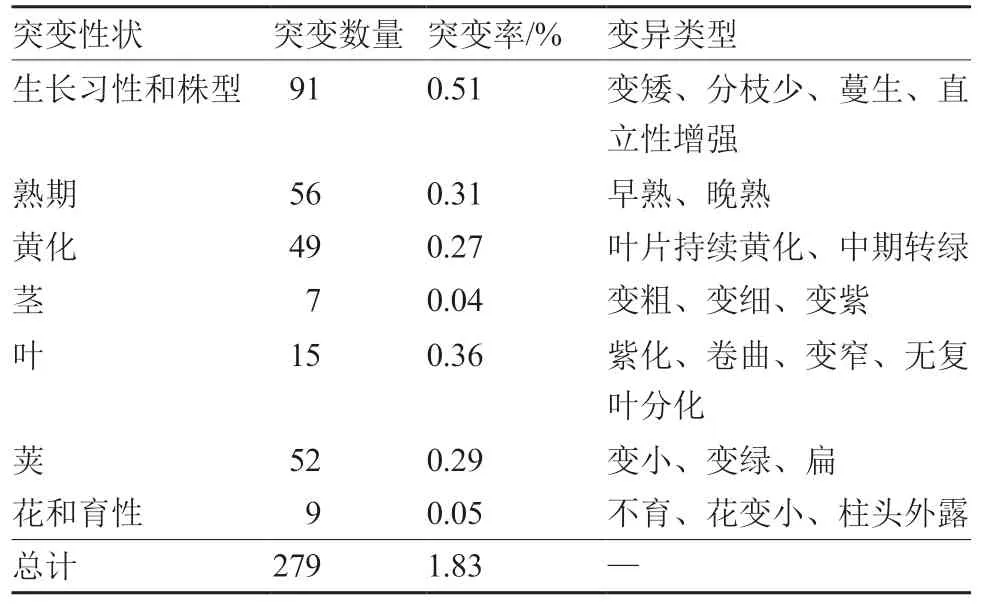
表2 M2表型突变统计及描述
2.2.1 生长习性和株型突变 诱变材料A18-1为有限生长习性的菜豆品种。在A18-1突变体库中观察到3株无限生长习性的突变体1-13-1、23-14-1和20-25-1,其中1-13-1和23-14-1表型相同,表现为开花时间延迟,花器官结构畸形,总状花序表现为无限型,花序轴顶端小花先开放,侧生小花开花延迟或停止发育(图2-a、2-b)。因其无法完成正常的授粉过程,而没有收获种子。20-25-1表现为叶脉紫红色(A18-1为浅绿色),长势弱,荚变宽、表皮出现红晕(图2-c、2-d)。
株型是由多基因控制的质量性状,与菜豆的产量性状直接相关。突变体库中株型突变最为丰富,包括分枝数减少、直立性增强、株高变矮等类型。突变体8-28-1、8-28-2和8-28-3在同一株系,表现出株高变矮、叶片卷曲的变异(图3-a)。而另一株系的5-5-1和5-5-2突变株则表现为分枝数减少(图 3-b)。

图2 生长习性突变

图3 株型突变
2.2.2 熟期突变 在突变体鉴定过程中,发现单行4株表现早熟的突变体(编号5-13-1、5-13-2、5-13-3和5-13-4),约占该株系的1/4,推测该突变为单基因突变。荚色持绿突变株10-7-1和紫花突变株5-22-1也表现为熟期略微提前。因为菜豆品种A18-1为早熟品种,所以熟期突变多表现为熟期延迟,在突变体库中共发现51株熟期明显延迟的突变株,部分单株的熟期甚至比对照平均熟期延迟30 d以上。
2.2.3 花和育性突变 菜豆A18-1的花为粉白色(图4-c),突变体中出现颜色加深的突变株,其中突变株10-6-1的花瓣出现深色的斑块(图4-a),突变株20-25-1的花瓣则变为紫红色(图4-b)。突变体库中还出现了花器官结构畸形(图4-d、4-e、4-f),如柱头外露影响胚珠受精而导致的育性降低。如16-2-1突变株的花序轴缩短,花瓣细长导致雌蕊和雄蕊暴露(图4-f)。在观察过程中还发现一类出现频率很高的独特败育表型,表现为开花时间延迟、花期长、花蕾后期未正常开放,叶色深绿、略微卷曲,荚变小、胚珠不膨大,无法形成正常的种子(图4-g、4-h)。有个别植株形成少量成熟的种子,推测该突变为雄性不育突变,少量正常发育的种子则可能由环境中昆虫传播花粉完成授粉(图4-i)。
2.2.4 茎突变 在突变体库中发现的茎突变较少,其中12-12-1和2-11-1突变株表现为茎和叶柄变细(图5-a),分别在不同的单行发现。此外,发现3株茎变粗的突变株26-9-1、26-9-2和15-21-1,其中26-9-1、26-9-2还表现出叶形的变异(图5-b、5-c)。2株茎颜色突变株5-22-1和20-25-1均表现为茎由绿色突变为紫色,与其他表型的变异同时出现。

图4 花和育性突变

图5 茎突变
2.2.5 叶突变 在突变体库中鉴定出多株叶形和叶色突变,其中5-14-1突变株表现为叶片变厚,叶片靠近叶柄基部的部分变窄(图6-a),且全株长势弱,开花晚、花瓣细长,花开放数天后凋谢,未能结荚。突变株26-9-1和26-9-2在同一单行发现,都表现为叶面积变大,叶由卵圆形变为近圆形(图6-b),该突变体除叶形突变外还表现出茎变粗,开花晚等其他性状的变异。突变株16-2-1叶片变小,三出复叶的小叶有2条主叶脉,形成2个细长的叶尖(图6-c),这一突变株还表现出花结构异常(图4-f)。

图6 叶突变
叶色变异表现为叶片黄化、叶脉突变为紫红色、叶片灰绿色。频率最高的为叶片黄化(图6-d),占突变总数的15%。将黄化突变体按照黄化持续时间分为持续黄化和后期转绿两类,持续黄化的突变株整体表现出植株矮小、长势弱、开花结荚少;而后期转绿的突变株在转绿后与对照植株相比无明显变异。叶色突变体中12-3-1、12-3-2和26-9-3为叶片灰绿色突变体(图6-e),其中12-3-1和12-3-2在同一单行。
此外,本试验还在同一单行发现4株独特的无叶片分化的突变株(21-20-1/-2/-3/-4),在营养生长阶段,植株正常长出2片初生真叶,出现多个分枝,但没有正常的叶片分化(图7-a)。在生殖生长阶段,分化出许多结构异常的小花,表现为花瓣变小、柱头外露(图7-b)。
2.2.6 荚突变 荚突变包括荚色和荚形突变。紫化突变株5-22-1的荚表面产生紫红色的条纹(图8-a),而突变株10-7-1的成熟荚由黄色突变为绿色,并且荚形也由圆棍形突变为宽扁形(图8-b)。突变株11-6-1的荚变小,胚珠不膨大(图8-c);突变株20-6-1则表现为荚变小、弯曲,结少量种子(图8-d)。

图7 无三出复叶分化突变体21-20-1

图8 荚突变
2.2.7 多种表型同时突变 植物的生长发育和器官的形态建成是一个复杂的网络调控过程,一种结构或生理的变化,也会对其他结构器官和生理性状产生影响,一些发育调控基因的突变往往引起多种表型的变化(Mizukami & Fischer,2000;王利,2016)。在突变体观察过程中,发现多种性状往往同时发生改变。例如5-14-1和16-2-1叶形变化的突变体,其花的结构也发生明显的变异。黄化突变体2-2-1,植株矮小,花的柱头明显外露,无法正常完成授粉受精,导致其败育。5-22-1紫化突变体除了表现出茎、叶、花、荚的紫化变异,还表现出荚熟期提前的变异。10-7-1荚色持绿突变株则同时表现出荚形变宽和熟期提前的变异。
3 结论与讨论
研究表明,植物在面临干旱、低温和高光胁迫时,花青素含量显著增加(宋明 等,2012;Ilk et al.,2015),表明花青素在响应逆境胁迫中发挥重要的作用。本试验发现的5-22-1紫化突变株的下胚轴、茎、叶、花和荚表皮均表现出紫色色素积累变异,将有望在干旱和低温抗性品种的选育中发挥作用。
菜豆叶形和叶色突变对于研究叶形态发育和色素积累具有重要的促进作用。在模式植物拟南芥的快中子诱变和EMS诱变体筛选中也发现了叶片灰绿突变体(朱勇,2005;曹雅君,2006)。在其他植物叶形突变体的研究中,已经鉴定了多个与叶形相关的基因,如拟南芥中菜素内酯信号途径相关基因BRH1调节叶片的生长发育并影响叶片的形态(Wang et al.,2018),番茄中的生长素转录因子SlIAA9/ENTIRE/E调控叶片的形态和果实发育(Wu et al.,2018),豌豆中AF和UNI基因影响叶片中生长素含量和分布,进而影响豌豆的叶片形态(Darleen et al.,2013)。在突变体库中,叶形的突变往往与花的结构异常同时出现。在观察到的多个叶形与叶色突变体中,花的结构都发生了一定程度上的变异,甚至直接导致花败育。这可能与叶片和花在形态发生上的同源性有关,虽然花在形态和功能上与叶差异很大,但在形态发生、生长方式和维管系统等方面与叶相似(张爱芹 等,2006)。已有研究证明菜豆生长习性是由PvTFL1y基因控制的质量性状(Repinski et al.,2012),无限生长习性相对于有限生长习性为显性,在拟南芥(Shannon & Meeks-Wagner,1991)、芝麻(Zhang et al.,2018)、木豆(Mir et al.,2014)和大豆(Liu et al.,2010)等多种植物中也发现相同功能的同源基因。本试验在有限生长习性的A18-1菜豆突变体库中观察到无限生长型的突变株,这一突变类型的出现,与Repinski等(2012)描述的菜豆生长习性的遗传模式不符,因此推测菜豆中应该还存在除PvTFL1y之外的控制生长习性的非同源基因。特定生理性状的变异,对于改良作物的品质性状和植株抗性方面具有重要的应用价值。本试验建立的突变体库是基于田间表型观察,而生理性状的突变则无法通过表型观察进行鉴别。为了进一步表征突变体的变异特征,下一步将对突变体的生理生化指标进行测定,明确不同表型变异的生理机制。
本试验利用126 Gy60Co-γ射线辐射处理菜豆的干种子,在M2鉴定了包括叶、花、茎、熟期、株型和育性在内的多种突变类型,为菜豆种质资源创新提供新材料。同时,重要性状的突变,如紫化、荚色和叶形的突变,为研究对应性状的遗传和分子机理提供了试验材料。构建菜豆突变体库,可以从突变体库中选择重要农艺性状突变株系用于杂交育种。
猜你喜欢
南方农业学报(2019年2期)2019-09-10
农业与技术(2017年22期)2017-12-29
现代园艺(2017年2期)2017-03-16
绿色科技(2017年3期)2017-03-14
农民致富之友(2016年22期)2017-02-07
江苏农业科学(2015年2期)2015-03-12
故事大王(2014年3期)2014-05-21
探索财富(2013年10期)2013-11-18
作文大王·中高年级(2008年5期)2008-07-31
