
鸡毒支原体GroEL蛋白通过NF-κB信号通路诱导DF-1细胞释放IL-1β的研究
2019-05-21 07:07朱可蒙赵雅芝郝文君辛九庆
中国预防兽医学报 2019年3期
张 琳,陈 莹,朱可蒙,赵雅芝,潘 巧,郝文君,于 颖*,辛九庆*
(1.中国农业科学院哈尔滨兽医研究所 兽医生物技术国家重点实验室,黑龙江 哈尔滨 150069;2.东北农业大学 动物医学学院,黑龙江 哈尔滨 150030)
鸡毒支原体(Mycoplasma gallisepticum,MG)是引起鸡慢性呼吸道疾病的病原体,在国内外养禽场中广泛流行。MG 感染机体后,可入侵禽类脑组织造成运动障碍,使受感染机体生产性能下降,体重减轻[1],同时造成T 淋巴细胞和B 淋巴细胞增殖异常,导致免疫抑制,从而使机体对其它病原,如:大肠杆菌、禽流感病毒、新城疫病毒和传染性支气管炎病毒等病原的易感性增加,引起继发感染,导致更为严重的呼吸道综合征,增加养殖业损失[2]。
MG 主要粘附和定植在呼吸道上皮细胞和淋巴组织等部位,诱发严重的组织损伤和炎性反应。支原体的膜表面存在大量的脂质相关膜蛋白(Lipid-associated membrane proteins,LAMPs),LAMPs 刺激宿主细胞激活NF-κB 信号通路,促进细胞因子的上调表达[3],从而介导炎症反应。本实验室前期研究显示 GroEL 蛋白是 LAMPs组分之一,在真核生物中被称为热休克蛋白60(Heat shock protein 60,HSP60)[4]。HSP 又称应激蛋白,是机体在高温、缺氧等外界条件刺激下高效表达的蛋白质。在细菌中GroEL 蛋白作为分子伴侣与其辅助蛋白GroES 共同帮助细菌内多种蛋白完成装配,使其具有完整的功能。在应急状态下,GroEL 过表达或异位表达,均能够诱发机体的保护性免疫应答,在信号转导中发挥作用[5-7]。
鉴于MG GroEL 是 LAMPs组分之一,GroEL 蛋白能否通过NF-κB 信号通路促进细胞因子的上调表达? 为此,本研究采用原核表达系统表达了MG 的GroEL 蛋白,通过激光共聚焦试验、SYBR Green I荧光定量 PCR方法、western blot 等检测方法对GroEL 蛋白展开研究,为揭示GroEL 蛋白的生物学功能及阐明MG 的致病机制提供数据支持。
1 材料与方法
1.1 菌株、细胞及载体 MG R 株、pGEX-6p-1 载体、pEGFP-p65 质粒、鸡胚胎成纤维细胞(DF-1)均为本实验室保存;DH5α 感受态细胞和BL21(DE3)感受态细胞购自天根生化科技(北京)有限公司。
1.2 主要试剂 Q5 DNA 聚合酶和T4 连接酶购自NEB 公司;DNA 提取试剂盒、胶回收试剂盒、质粒提取试剂盒均购自天根生化科技(北京)有限公司;限制性内切酶购自Thermo 公司;Glutathione SepharoseTM4B 购自 GE 公司;膜蛋白提取试剂盒购自Novagen 公司;BAY 11-7082(NF-κB 抑制剂)、BCA定量试剂盒、细胞膜红色荧光探针DiI 均购自上海碧云天生物技术有限公司;山羊抗兔FITC-IgG、细胞核荧光染料 DAPI 购自Sigma 公司;IRDye 680RD山羊抗兔IgG 和山羊抗鼠IgG 抗体购自LI-COR Bioscience 公司;SYBR Green Master、反转录试剂盒购自 Roche 公司;GST 抗体和 p65 磷酸化抗体购自CST 公司;GroEL 蛋白的单克隆抗体(MAb)(3G9)由本实验室制备。
1.3 目的基因扩增及重组质粒构建 按照DNA 提取试剂盒的说明书提取MG R 株基因组,并以其为模板,根据 GroEL 基因序列(NM_001012916)分别设计引物(5'-CGCGGATCCATGGCAAAAGAATTAACA TTTGAACA-3'/5'-CGCGTCGACTTATAGGTGATTTA AGCTTGGTTTTTC-3')进行PCR扩增。PCR产物经BamHⅠ/SalⅠ双酶切后克隆至pGEX-6p-1 载体中构建原核重组表达质粒pGEX-GroEL 并测序鉴定。
1.4 重组蛋白的表达与纯化 将重组质粒pGEXGroEL转化至E.coliBL21(DE3)中,重组菌37 ℃180r/min 摇菌至OD600nm0.6 时,加入终浓度为0.1mmol/L IPTG,22 ℃ 180 r/min 诱导 6 h 后离心收集菌体,超声破碎离心后,将上清液利用GST-resin 纯化,纯化的重组GST-GroEL 蛋白经超滤去除咪唑后,应用BCA 定量试剂盒测定纯化蛋白的含量。
1.5 重组蛋白的鉴定 将表达的GST-GroEL 蛋白经 SDS-PAGE后,转移至 NC 膜,经 5 %的脱脂乳封闭后,以GroEL MAb 3G9 (1∶1000)为一抗,IRDye 680RD 山羊抗鼠 IgG (1∶10 000)为二抗,进行western blot 鉴定。
1.6 GST-GroEL 蛋白粘附DF-1 细胞的检测 将1 μg GST-GroEL 蛋白与 DF-1 孵育细胞过夜后,利用4%多聚甲醛室温固定30 min,5%的脱脂乳37 ℃封闭 2 h,以抗 GST 抗体(1∶100)作为一抗,山羊抗兔 FITC-IgG (1∶500)为二抗,并用 DiI 和 DAPI 分别对细胞膜和细胞核染色,利用激光共聚焦显微镜观察重组蛋白粘附情况。同时设立GST 蛋白作为对照组。
1.7 GST-GroEL 蛋白刺激 DF-1 细胞后 IL-1β 转录水平的检测 分别用 0、1 μg/mL、2 μg/mL、4 μg/mL、6 μg/mL GST-GroEL 蛋白刺激 6 孔板中的 DF-1 单层细胞,12 h 后,收集细胞,采用 SYBR Green I 荧光定量 PCR 检测 IL-1β mRNA 转录水平[3]。IL-1β 上游引物:5'-AACATCGCCACCTACAAG-3';下游引物:5'-GACGGTAATGAAACATAAACG-3'。GADPH 上游引 物 :5'-ATTCTACACACGGACACTTCA-3'下游引物:5'-CACCAGTGGACTCCACAACATA-3'。
1.8 激光共聚焦检测 p65 转核 将 2 μg pEGFP-p65 质粒转染 DF-1 细胞,转染 24 h 后,采用 1.7筛选出的最佳剂量的 GST-GroEL 刺激 DF-1 细胞6 h,4%多聚甲醛室温固定30 min,0.1% TritonX-100透膜 30 min,5 %的脱脂乳 37 ℃封闭 2 h,DAPI 染核。利用激光共聚焦显微镜观察p65 入核。
1.9 GST-GroEL 蛋白诱导DF-1 细胞p65 磷酸化的检测 将DF-1 细胞铺于六孔板中培养至密度为80%左右,利用1μg/mL、2 μg/mL、4 μg/mL GST-GroEL 蛋白刺激DF-1 细胞。12 h 后裂解细胞、获取总蛋白,SDS-PAGE 电泳后将蛋白转印于NC膜上,经 5 %的脱脂乳 37 ℃封闭2 h 后,以抗 p65(1∶1 000)抗体作为一抗,IRDye 680RD 山羊抗兔 IgG(1∶10 000)为二抗,同时以 GST 刺激的 DF-I 细胞作为对照,利用western blot 检测p65 磷酸化水平。
1.10 抑制NF-κB 信号通路后检测IL-1β 释放 将DF-1 细胞铺于12 孔板中培养至密度为80 %左右,分别设对照组,GST-GroEL 刺激组(GST-GroEL),抑制剂组(BAY 11-7082),抑制后刺激组(BAY 11-7082+GST-GroEL),每组设 3 孔。对照组不做任何处理;GST-GroEL 刺激组为每孔加入2 μg GST-GroEL 蛋白;抑制剂组为每孔加入终浓度为 5 μmol/L 的 BAY 11-7082;抑制后刺激组为加入终浓度5 μmol/ LBAY 11-7082 培养 2 h 后,加入 2 μg 的 GST-GroEL 蛋白。各组继续培养12 h 后收集细胞,采用SYBR Green I荧光定量PCR检测IL-1β mRNA 转录水平。
2 结 果
2.1 目的基因扩增及重组质粒构建 以MG 基因组为模板,PCR 扩增获得全长约为 1600 bp 的目的基因,其大小与预期相符(图1)。将该片段利用BamHⅠ/SalⅠ双酶切后克隆于pGEX-6p-1 载体中构建重组表达质粒pGEX-GroEL 并测序鉴定。测序结果显示,插入的GroEL 基因序列与MG R 株GroEL 碱基序列一致性为100 %,表明重组质粒 pGEX-GroEL 正确构建。
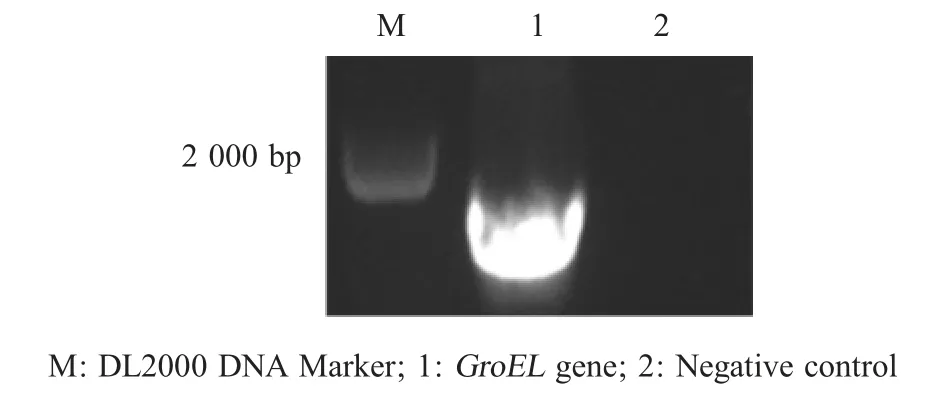
图1 PCR扩增GroEL基因Fig.1 Amplication of the GroEL gene by PCR
2.2 重组蛋白的表达、纯化及鉴定 将重组质粒pGEX-GroEL 转化至 BL21(DE3)感受态细胞,选取单菌落经PCR 鉴定阳性后诱导表达并纯化,SDSPAGE 结果显示,重组蛋白GST-GroEL 在大肠杆菌中得到了表达,大小为86 ku,与预期相符(图2A)。Western blot 鉴定结果显示,在86 ku 处可见特异性反应条带(图2B),表明 GST-GroEL 获得了正确表达,且其有较好的反应原性。

图2 重组蛋白的表达纯化及鉴定Fig.2 Anaylsis of purified GST-GroEL protein by SDS-PAGE (A)and western blot (B)
2.3 GST-GroEL 蛋白对DF-1 细胞粘附特性的检测结果 将 GST-GroEL 蛋白与 DF-1 细胞孵育后,利用激光共聚焦试验检测,结果显示,实验组细胞膜表面出现明显的绿色荧光,对照组中未见绿色荧光(图3)。表明 GST-GroEL 蛋白能够粘附至 DF-1 细胞表面。

图3 GroEL蛋白对DF-1细胞粘附的检测结果Fig.3 The adhesion of GroEL protein to DF-1 cells
2.4 GST-GroEL 蛋白刺激 DF-1 细胞 IL-1β 转录水平检测结果 分别以不同浓度GST-GroEL 蛋白刺激DF-1 细胞,利用荧光定量PCR 检测DF-1 细胞中IL-1β mRNA 转录水平。结果显示,2 μg/mL GSTGroEL 蛋白刺激 DF-1 细胞时,IL-1β mRNA 转录水平最高(p<0.01)(图4)。表明GST-GroEL可以诱导DF-1细胞 IL-1β 的释放,且 2 μg/mL 为 GST-GroEL蛋白最佳剂量。

图4 GST-GroEL蛋白刺激DF-1细胞对其IL-1β转录水平影响的检测Fig.4 Transcription of IL-1β induced with different concentrations of GST-GroEL detected by real-time PCR
2.5 激光共聚焦检测p65 转核的结果将2μg pEGFP-p65质粒转染DF-1细胞后,以 2 μg/mL GST-GroEL 刺激DF-1 细胞6 h 后利用激光共聚焦显微镜观察。结果显示,实验组细胞核内可见绿色荧光 (图5)。表明GST-GroEL能够促进DF-1细胞中pEGFP-p65 质粒转入细胞核。
2.6 GST-GroEL 蛋白诱导DF-1 细胞 p65 磷酸化的检测结果 分别以不同浓度的GST 和GST-GroEL蛋白刺激DF-1 细胞,western blot 检测 p65 磷酸化水平,结果显示,在 1 μg/mL、2 μg/mL、4 μg/mL 的GST-GroEL 蛋白刺激下,实验组p65 磷酸化水平均明显高于对照组(图6)。表明 GST-GroEL 蛋白能够促进DF-1 细胞中p65 磷酸化水平升高。
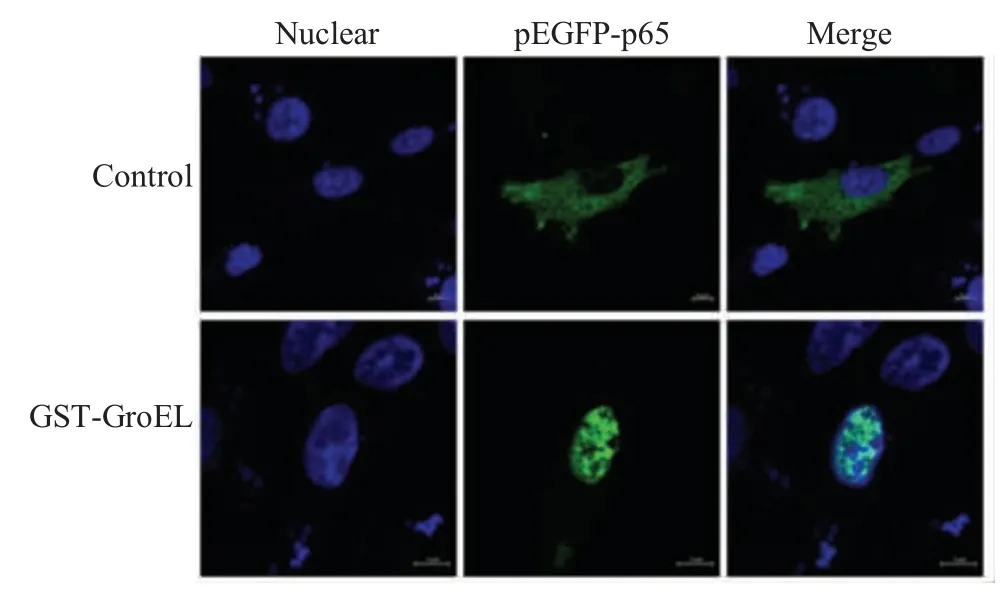
图5 激光共聚焦试验观察GroEL蛋白刺激DF-1后p65的入核Fig.5 The observation of p65 nuclear translocation under the laser-scanning confocal microscope
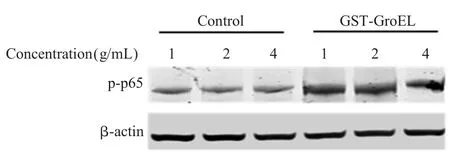
图6 Western blot验证GST-GroEL蛋白对DF-1细胞p65磷酸化的促进作用Fig.6 The level of p-p65 induced with GST-GroEL detected by western blot
2.7 抑制NF-κB 信号通路后IL-1β 转录水平的检测结果 收集各组DF-1 细胞,利用荧光定量PCR检测 DF-1 细胞中 IL-1β mRNA 转录水平。结果显示,对照组和抑制剂组IL-1β mRNA 转录水平基本未变化,而抑制后刺激组(BAY 11-7082+GSTGroEL) IL-1β mRNA 转录水平显著低于 GST-GroEL刺激组(图7)。表明 GST-GroEL 蛋白能够通过NF-κB 信号通路诱导 DF-1 细胞释放 IL-1β。
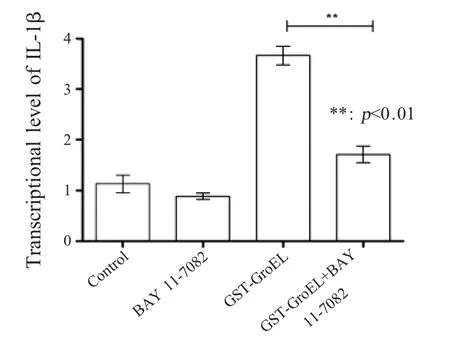
图7 NF-κB信号通路对DF-1细胞分泌IL-1β影响的检测Fig.7 NF-κB signaling pathway impact on the transcription of IL-1β in DF-1 cell detected by real-time PCR
3 讨 论
支原体粘附于宿主细胞表面是引起其致病的前提条件[8]。MG 主要粘附和定植在呼吸道上皮细胞、淋巴组织等部位,并诱发严重的组织损伤和炎性反应[9]。MG 通过 LAMPs 来粘附和感染宿主细胞,诱导宿主细胞产生强烈的免疫应答并释放相应细胞因子[10]。在 MG 的研究中,气管上皮细胞和 DF-1 细胞常被作为细胞模型。由于制备的气管上皮细胞在体外不能连续传代培养,不能满足实验需要,因此本研究没有选择气管上皮细胞;而DF-1 为鸡成纤维细胞系,是一种可稳定传代、无肿瘤基因、能够无限增殖的细胞系,已被广泛应用于鸡相关疾病的研究[11],因此本实验选用DF-1 细胞作为细胞模型。目前已发现多个 MG 粘附因子[12-13]。本研究通过激光共聚焦试验显示GST-GroEL 能够粘附于DF-1 细胞表面,结果表明GroEL 是MG 中一个新发现的粘附因子。
本研究显示GroEL 蛋白不仅可以作为粘附分子,还具有上调 IL-1β表达的能力。IL-1β 是重要的炎性因子,参与机体免疫应答,诱导细胞凋亡及直接杀伤靶细胞等[14]。研究表明 IL-1β 主要由 NF-κB信号通路调控产生[15]。NF-κB家族由p50、p52、p65、c-Rel 和 RelB组成,这 5 个家族成员均具有 N端 Rel 同源结构域(Rel homology domain, RHD)[16]。NF-κB 常以二聚体的形式存在,最常见的二聚体结构为p65/p50。p65 含有转录激活区域,参与转录的起始调节,具有转录激活功能,也是研究最多的经典信号通路中的细胞因子;而 p52、c-Rel 和 RelB则是非经典NF-κB 信号通路中的细胞因子。因此本研究选择p65 分子进行研究[15]。当细胞在静息状态时,NF-κB 与 IκB(Inhibition of κB)结合组成一个三聚体 p50/p65/IkB,由于 IkB 的存在阻碍了 p50/p65复合物形成,核定位信号被掩盖使NF-κB 的活性丧失。当细胞受到病原微生物刺激时,IκB 从三聚体中解离出来,使二聚体表现出NF-kB 活性,发生磷酸化并进入细胞核内,启动下游炎症相关的细胞因子和炎症介质的转录[17]。因此 p65 磷酸化或 p65 从胞浆进入细胞核可以作为NF-κB 信号通路被激活的标志。本研究结果显示GroEL 蛋白促进了p65 入核并提高了 p65 磷酸化水平,从而激活了 NF-κB 信号通路;当 NF-κB 信号通路被抑制后,GroEL 蛋白刺激DF-1 细胞产生IL-1β mRNA 转录水平显著下降。以上结果表明GroEL 蛋白具有粘附DF-1 的能力并能够通过激活 NF-κB 信号通路促进 IL-1β 释放,从而激活宿主的天然免疫应答。
目前,研究许多支原体的 LAMPs 能够激活NF-κB 信号通路诱导 IL-1β 的释放[18-19],从而介导炎症反应和调节宿主免疫应答。但是关于LAMPs 中的哪一种成分能够激活NF-κB 信号通路研究较少。本研究显示GroEL 蛋白作为MG LAMPs 中的关键膜蛋白可以粘附到DF-1 细胞表面,直接刺激宿主细胞并促进宿主细胞的p65 磷酸化和入核,启动了NF-κB 信号通路。本研究从单一组分的角度分析MG 刺激天然免疫的通路,为揭示MG 致病机制的研究提供了参考,对研制基因工程疫苗提供靶标等具有重要价值。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
波谱学杂志(2022年1期)2022-03-15
昆明医科大学学报(2022年1期)2022-02-28
湖南畜牧兽医(2021年6期)2022-01-24
食品安全导刊(2021年21期)2021-08-30
猪业科学(2021年5期)2021-06-02
江西农业学报(2021年4期)2021-04-20
三农资讯半月报(2020年11期)2020-06-21
质量安全与检验检测(2020年6期)2020-02-01
