
蓝舌病病毒10质粒反向遗传操作系统的建立
2019-05-21 07:07郭运泽孙恩成徐青元步志高吴东来王凤龙1
中国预防兽医学报 2019年3期
郭运泽,孙恩成,徐青元,步志高,吴东来*,王凤龙1,*
(1.内蒙古农业大学 兽医学院,内蒙古 呼和浩特 010018;2.中国农业科学院哈尔滨兽医研究所 兽医生物技术国家重点实验室/农业部兽医公共卫生重点开放实验室,黑龙江 哈尔滨 150069)
蓝舌病是由蓝舌病病毒(Bluetongue virus,BTV)引起的,导致多种反刍动物及野生动物发病的急性热性疾病。由于该病传播迅速以及其在全球范围内造成的严重经济影响,被世界动物卫生组织(OIE)列为法定通报疾病,在我国也被列为一类动物疫病。由于近年来蓝舌病在欧洲的暴发,且传播范围的不断扩大,蓝舌病的防控已成为我国乃至国际社会关注的重要动物疫病。BTV 是一种隶属于呼肠孤病毒科环状病毒属的虫媒病毒。其基因组由10 条线性化的双链RNA组成。BTV 的病毒粒子呈二十面体对称,无囊膜,直径约为 70 nm~80 nm。BTV 的核酸由10 个基因节段的双股RNA (double-stranded RNA,dsRNA)组成,按长度大小依次命名为 Seg-1~Seg-10。BTV 的 10 个双股 RNA 节段总长度约 19.2 kb,分别编码7 种结构蛋白(VP1~VP7)和4 种非结构蛋白(NS1、NS2、NS3/NS3A、NS4)[1]。
本研究所采用的10 质粒拯救系统操作过程比现有的体外转录系统简便,可丰富dsRNA 病毒的反向遗传操作系统。同时,该系统为BTV 发病机理等方面的基础研究以及新型疫苗研发方面的应用性研究奠定了一定的基础。
1 材料与方法
1.1 主要实验材料 亲本病毒BTV-16 BN96/16 株(wtBTV-16)和BHK-21、BSR 细胞均由本实验室保存;携带 BTV-16 BN96/16 株的 Seg-1~Seg-10 的重组质粒pB16-s1~pB16-s10 均由本实验室构建保存;反向遗传操作系统通用载体pS111(图1)由本实验室构建保存[2];抗 BTV-16 NS2 蛋白的单克隆抗体(MAb)腹水BTV-NS2-1B2 由本实验室制备保存[3]。
RESRAD-BIOTA广泛应用于美国能源部的部门和项目,以及美国多个洲的环保部门。同时,该软件还用于IAEA的EMRAS项目中各种软件在不同场景中的对比研究中。
1.2 主要试剂 DNA 聚合酶KOD FX 购自TOYOBO公司;限制性内切酶AarI、BbsI、BsaI和BsmB I均购自Thermo Fisher公司;Quick Ligation酶购自NEB公司;TransI 感受态细胞购自北京全式金生物技术有限公司;Plasmids midi kit 购自QIAGEN 公司;Opti-MEM 培养基购自Gibco 公司;转染试剂 LipofectamineLTX & PLUSTMReagent购自Invitrogen 公司;FITC 标记的山羊抗小鼠IgG(IgG-FITC)购自Sigma 公司。引物由安徽通用生物技术有限公司合成。
2.1 病毒10 个基因节段的扩增与反向遗传质粒的构建 采用10 对引物以BTV-16 病毒基因组为模板PCR 扩增各基因节段,分别得到S1~S10 10 个基因节段的 PCR 扩增产物,其长度依次约为4 000 bp、3 000 bp、2 800 bp、2 000 bp、1 800 bp、1 600 bp、1 200 bp、1 100 bp、1 000 bp 和 800 bp,经测序显示扩增产物均与预期一致(图2)。将各个基因采用相应的酶切后克隆至pS111 载体中,将阳性克隆的测序结果登录GenBank(分别为JN671906~JN671915),比对结果显示,10 个阳性克隆基因节段的序列与相应基因标准序列的相似性均为100 %,表明本实验正确构建了重组质粒pS161~pS1610。
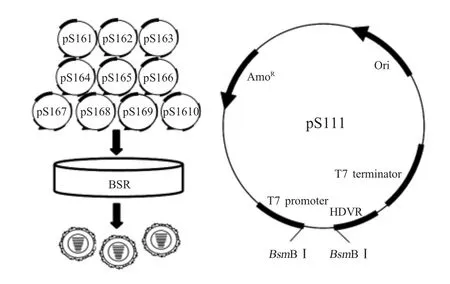
图1 10质粒系统拯救BTV的实验策略以及pS111的质粒图谱Fig.1 Experimental strategy to rescue BTV from 10 plasmids and the diagram of pS111
1.3.1 BTV-16 基因组引物的设计和片段的扩增根据 BTV-16 的基因序列,利用 Primer Premier 5 软件分别针对BTV-16 的10 个基因节段 Seg-1~Seg-10设计含不同酶切位点的上下游引物(表1)。以BTV-16 重组质粒为模板,利用 KOD FX 通过 PCR方法扩增病毒的10 个基因节段的全长序列。

表1 扩增BTV-16 基因组所用引物Table 1 Primers for the BTV-16 genome cDNA amplification
1.3.2 10 个重组质粒的构建 将上述获得的10 个PCR 产物回收,根据各对引物设计的酶切位点分别酶切纯化处理后,使用Quick Ligation 分别将目的片段克隆至同样酶切纯化的pS111 载体,转化后挑取单克隆测序鉴定。将鉴定正确的重组质粒依次命名为 pS161~pS1610。
1.4 质粒的转染和重组病毒的拯救 对构建的10个重组质粒,按照Plasmids midi kit 说明书进行质粒抽提。经超微量紫外分光光度计测定浓度后,根据各基因节段的大小转染质粒的质量pS161~pS1610依次为 1 600 ng、1 350 ng、1 300 ng、1 100 ng、1 050 ng、1 050 ng、900 ng、900 ng、900 ng 和 800 ng。将中提后的10 个重组质粒按照LipofectamineLTX & PLUSTMReagent 说明书进行转染。在转染单层 BSR 细胞 6 h 后补加含 5 %FBS 的 DMEM 培养液,于 37 ℃ 5 % CO2条件下持续培养,并观察细胞病变(CPE)。同时,以仅加入不含转染试剂的单层BSR 细胞作为空白对照。当观察到细胞约有80%出现 CPE 时,收集细胞及上清,冻融 3 次后,收集病毒液,经 BHK-21 细胞连续传代。经鉴定后,将拯救病毒命名为rBTV-16,-80 ℃保存备用。
1.5.4 拯救病毒基因组的测序鉴定 参照Maan 等[5]建立的用于dsRNA 病毒全长(包括两端非编码区序列)基因测序的cDNA 扩增法(Full-length amplification of cDNAs,FLAC),进行 rBTV-16 的 S1~S10 基因节段的PCR 扩增并测序,比对分析拯救病毒与亲本病毒的基因组序列。
1.5 拯救病毒的鉴定
1.5.1 间接免疫荧光(IFA)鉴定检测 将拯救的病毒在 BHK-21 细胞传至第 5 代后,以 BTV-NS2-1B2 MAb (1∶100)为一抗,山羊抗小鼠 IgG-FITC (1∶200)为二抗,按照常规方法进行IFA 的检测。设未感染拯救病毒的BHK-21 细胞为空白对照。
2.5 拯救病毒dsRNA 基因组鉴定 利用TRIzol LS试剂提取病毒液中病毒总RNA,经过纯化后可获得除去ssRNA 后的病毒基因组 dsRNA。经 10 %的非变性聚丙烯酰胺凝胶电泳分析,结果显示rBTV-16的10 个基因节段的dsRNA 条带与带wtBTV-16 一一对应(图5),大小也完全一致。表明正确拯救重组病毒rBTV-16。
1.5.3 病毒dsRNA 基因组的电泳检测 利用TRIzol鉴定法提取rBTV-16 及其亲本病毒wtBTV-16 的总RNA,参照 Attoui[4]和 Maan[5]等的方法获得去除 ss-RNA 后的病毒基因组dsRNA。使用非变性聚丙烯酰胺凝胶电泳检测病毒的dsRNA 基因组。
景观中常用的装饰混凝土有多种类型,其中比较常见的有清水混凝土、彩色混凝土、异型混凝土(GRC、泰科石等),露骨料装饰混凝土等。
2.6 拯救病毒的基因组测序 将提取的病毒RNA反转录后利用PCR 扩增,扩增产物测序结果显示拯救病毒与亲本病毒序列完全一致。进一步表明拯救出的病毒确为目的病毒。
我不知道到哪里去找寻白丽筠,打她的手机停机,上网QQ呼她,头像是灰的,我留心各种媒体报道的车祸事故,甚至去我们这座小城的所有水域巡查,没有任何疑似消息,一概没有。我痛感到我与白丽筠的联系其实多么脆弱,就像孩子手里的一只风筝,只要一阵风就把我们彻底拆散了,再也找不到。最后,我去了白丽筠工作的售楼部,明知道没有用,还是去了。售楼部经理说,白丽筠已经辞职了。“辞职”两个字让我冰凉的心里升起一丝暖意,既然白丽筠想到要辞职,就不像是一个要去自杀的人。
2 结 果
1.3 病毒基因组的扩增和反向遗传质粒的构建10 质粒系统拯救BTV 的原理如图1所示[2]。
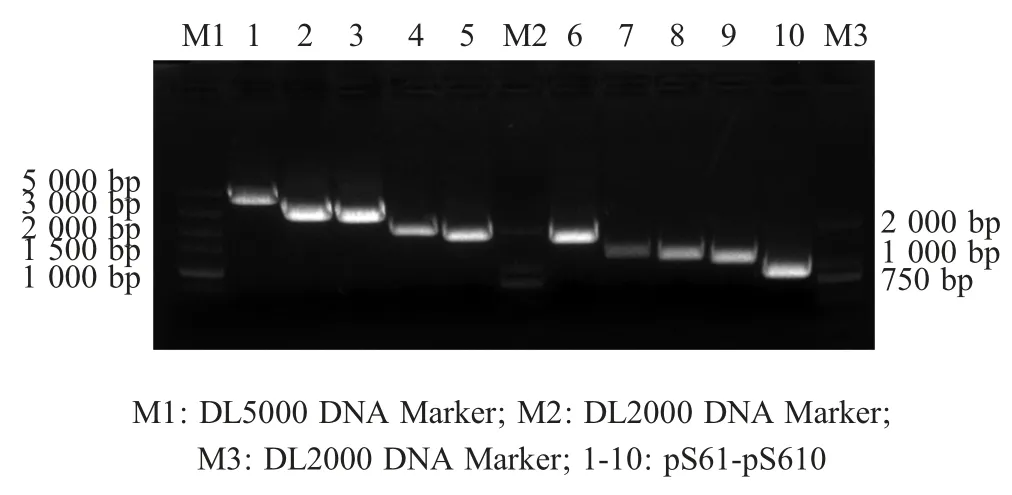
图2 BTV16 10个基因节段的PCR扩增结果Fig.2 Amplification of 10 gene segments of BTV-16 by PCR
2.2 反向遗传重组病毒的拯救 待BSR 细胞长成单层时转染10 个质粒,培养后观察可见实验组细胞出现 CPE,而对照组无 CPE。表明拯救得到重组病毒。
2.3 拯救病毒的 IFA鉴定将rBTV-16 感染BHK-21 细胞 24 h 后经 IFA 鉴定,结果显示感染rBTV-16 的BHK-21 细胞中可见明显的绿色荧光(图3) 。表明拯救的重组病毒可以感染BHK-21 细胞。
路灯心跳的主要目的是保活应用层连接,属于非必须功能,尤其考虑通信按连接收取资费,客户可能会减少甚至取消心跳;因此路灯心跳时间Tm参数设计为可配变量,当前IoT平台设定的应用层老化时间一般为30min,建议设置为25min。如果心跳过于频繁,会对网络资源、用户资费造成浪费。

图3 IFA鉴定拯救的重组病毒感染BHK-21细胞的结果Fig.3 Identification of the rBTV-16 in infected BHK-21 cells by IFA
2.4 拯救病毒的电镜观察 收集病毒液,经处理后在透射电子显微镜下观察,可见直径约为70 nm 的病毒粒子,无囊膜(图4),拯救的重组病毒的形态为典型的环状病毒属病毒粒子形态。表明拯救的重组病毒形态标准。

图4 拯救病毒的电镜照片Fig.4 Morphological examination of rBTV1 by electron microscope(Bar=200 nm)
1.5.2 电镜观察 将拯救的rBTV-16 在BHK-21 细胞中增殖,观察到细胞出现80 % CPE 时反复冻融,按常规方法经负染后电镜观察病毒形态。
目前,国内主要采用体外转录法进行病毒的拯救。根据文献报道,该方法主要是通过先转染真核表达质粒表达 VP1、VP3、VP4、VP6、VP7、NS1、NS2 几个辅助蛋白,再转染带 5' 帽结构的 T7 体外转录物进行病毒的拯救[6-7]。该反向遗传操作系统也被用来拯救呼肠孤病毒科环状病毒属的其它成员,如非洲马瘟病毒(Efrican horse sickness virus,AHSV)、鹿流行性出血热病毒(epizootic hemorrhagic disease virus,EHDV)等[8-9]。而国外已经建立两种反向遗传操作系统:一种为先转染7 个辅助质粒,后转染体外获得的10 个帽化或非帽化的RNA,最后收获拯救病毒[8];另一种为10 质粒拯救系统。本实验室在2014年根据国外发表文献已经构建7 个辅助质粒加10 个体外转录RNA 共转染拯救系统[8],但是该系统效率较低,费时费力。为了优化BTV 的拯救,本实验构建了10 质粒拯救系统,这在国内目前尚属首次报道。

图5 BTV-16基因组的非变性聚丙烯酰胺凝胶电泳Fig.5 Detection of the genomic dsRNA of rBTV-16 in an 10%non-denaturing polyacrylamide gel
1.6 拯救病毒与亲本病毒的生长曲线测定 待BHK-21 细胞汇聚成单层时将rBTV-16 与wtBTV-16传至第 5 代,分别以 MOI 0.1感染细胞,接毒 1 h 后弃去病毒液,PBS 洗两遍后更换 5 %FBS 的DMEM。每间隔 12 h 收取细胞上清液,测定病毒TCID50并绘制生长曲线。
2.7 拯救病毒与亲本病毒生长曲线的测定 将rBTV-16与wtBTV-16分别感染BHK-21细胞,经TCID50测定并绘制二者的生长曲线。结果显示,rBTV-16 与wtBTV-16 具有相似的生长曲线,且最高病毒滴度可达 1×107TCID50/mL 以上(图6)。表明拯救的重组病毒的复制能力与其亲本病毒的复制能力相当。
图6 拯救病毒与亲本病毒的生长曲线Fig.6 The growth curves of rBTV-16 and its parental virus of wtBTV-16
3 讨 论
3.5.2饲料配方小鸡阶段:玉米58%、豆粕30%、鱼粉5%、酵母3%、油脂1%、骨粉1.5%、食盐0.5%、预混合饲料1%。中鸡阶段:玉米62%、豆粕27%、鱼粉3%、酵母3%、油脂2%、骨粉1.5%、食盐0.5%、预混合饲料1%。大鸡阶段:玉米65%、豆粕23%、鱼粉2%、酵母4%、油脂3%、骨粉1.5%、食盐0.5%、预混合饲料1%。
本研究前期采用重叠延伸PCR 方法构建了反向遗传质粒pS111。以pUC57 载体为骨架分别加入T7 RNA 聚合酶启动子序列、限制性酶切位点序列、丁型肝炎核酶序列和T7 RNA 聚合酶终止子序列构建的重组质粒作为载体,并命名为pS111[2]。本研究采用的反向遗传构建的载体pS111,在BTV 基因节段合成的cDNA 5' 端含有T7 RNA 聚合酶启动子序列,3' 端含有 HDVR 和 T7 RNA 聚合酶终止子序列;利用HDVR 的自剪切活性,保证了目的基因在3' 端的真实性[1]。在体外转录系统中,由于在5' 端需要添加帽结构,在使用帽化酶情况下可能仅一部分T7转录物形成了该结构,除了影响翻译过程外,也可能影响RNA 的包装、负链的合成以及下一轮感染的转录过程;同时在体外转录过程中,可能产生比目的基因或长或短的转录物,这些有缺陷的转录物很可能会影响病毒的拯救效率[2,5]。
因为所有BTV 基因节段的5' 端和3' 端各具有4个保守的核苷酸序列,所以本研究设计选用AarI、BbsI、BsaI、BsmB I 这几种限制性内切酶位点序列,使得PCR 产物酶切后得到相应的保守序列粘性末端,并应用于所有BTV 基因节段的克隆[6]。
反向遗传系统不仅能够推进BTV 的基础研究,包括病毒的复制、入侵、包装,还可以应用于基因工程疫苗的研究。尤其是拯救不同基因节段相互替换的基因重组病毒,可以作为筛选针对不同血清型BTV 的疫苗株。国外已经有报道通过基因节段替换拯救出可应用于后续疫苗研发的病毒株[9-10]。
选择我院2016年1月-2017年12月急诊室收治的130例下肢骨折患者,将130例患者入院时护理模式不同进行分组,常规急诊护理患者设定为对照组,循证护理治疗患者为观察组,每组有患者65例,对照组:男性35例,女性30例,年龄18-78岁,平均(42.5±6.3)岁;观察组:男性36例,女性29例,年龄18-79岁,平均(41.5±6.4)岁;两组一般资料无统计学差异,P>0.05。
其实,在上世纪70年代时,运用在钟表机心上的防震器种类繁多,而KIF防震器与Incabloc防震器相较之功能优异、产量大而品质稳定,许多同时期的防震器,因制造昂贵与维修不易,逐渐被大部分的表厂所舍弃。
本研究建立了一个全新的系统扩展了现有的BTV 反向遗传操作平台,不仅在很大程度上减少了试剂的花费、实验的周期,而且提高了病毒拯救的效率。考虑到环状病毒具有相似的结构和复制机制,该系统也可以应用于其它环状病毒属成员的反向遗传操作。
猜你喜欢
铁道建筑技术(2022年10期)2022-10-28
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
西南农业学报(2022年5期)2022-06-06
中国典型病例大全(2022年12期)2022-05-13
中国典型病例大全(2022年9期)2022-04-19
中国糖料(2021年3期)2021-07-13
江西农业学报(2021年4期)2021-04-20
三农资讯半月报(2020年11期)2020-06-21
热带农业科技(2019年1期)2019-01-14
